
Cytotoxic Activity of LLO Y406A Is Targeted to the Plasma Membrane of Cancer Urothelial Cells

 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cytotoxicity of LLO Y406 and wt LLO on NPU and RT4 Cells
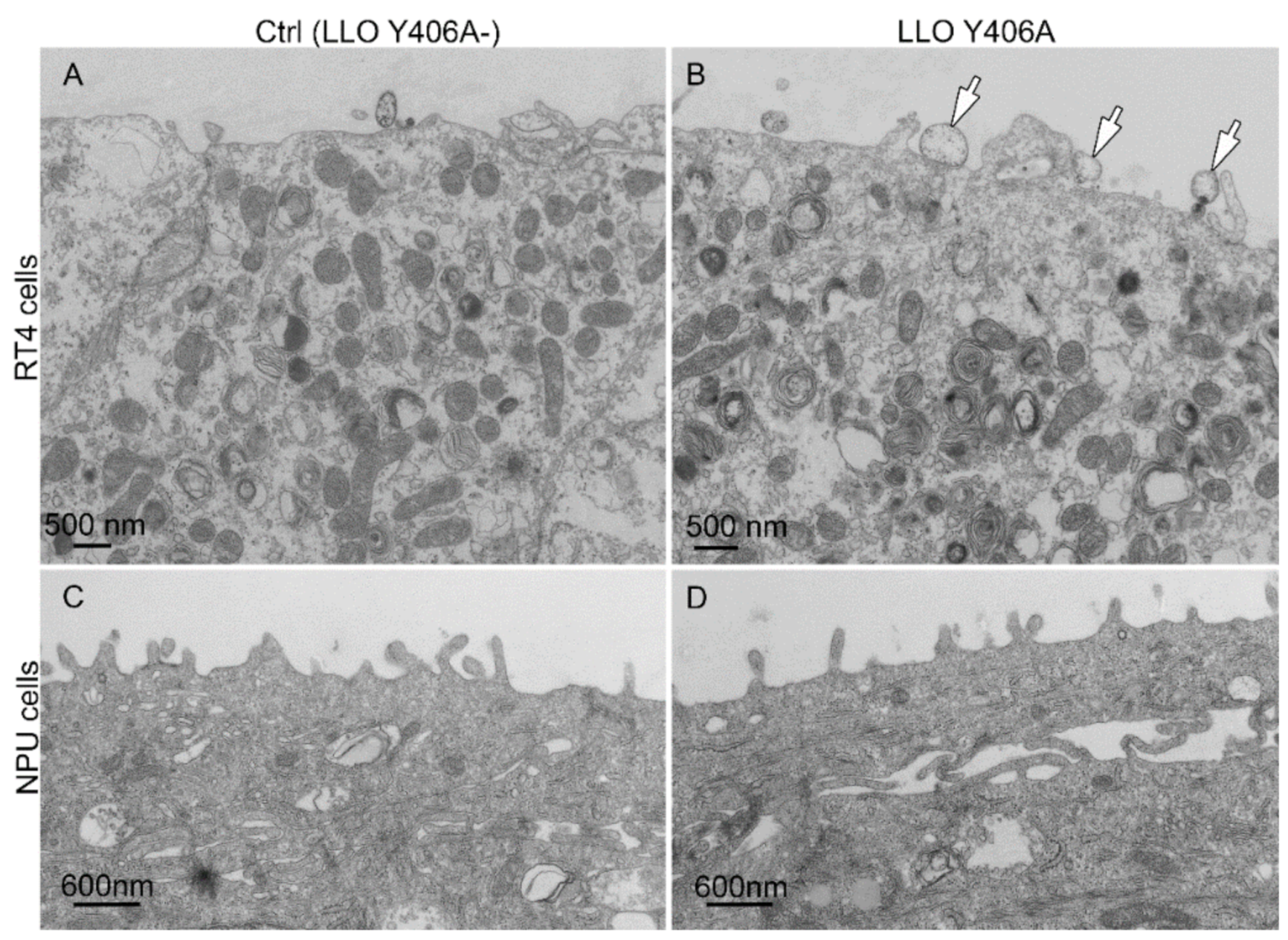
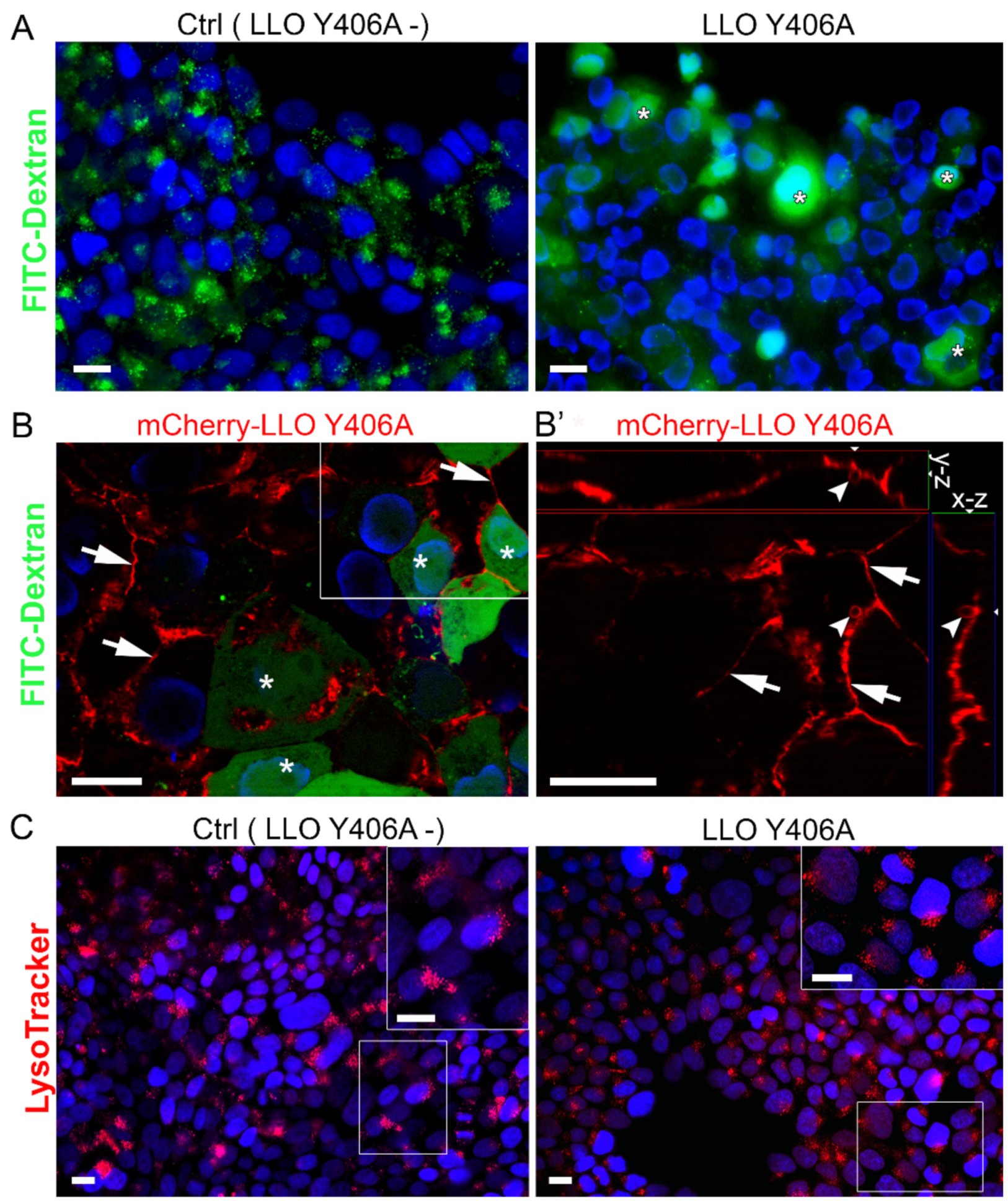
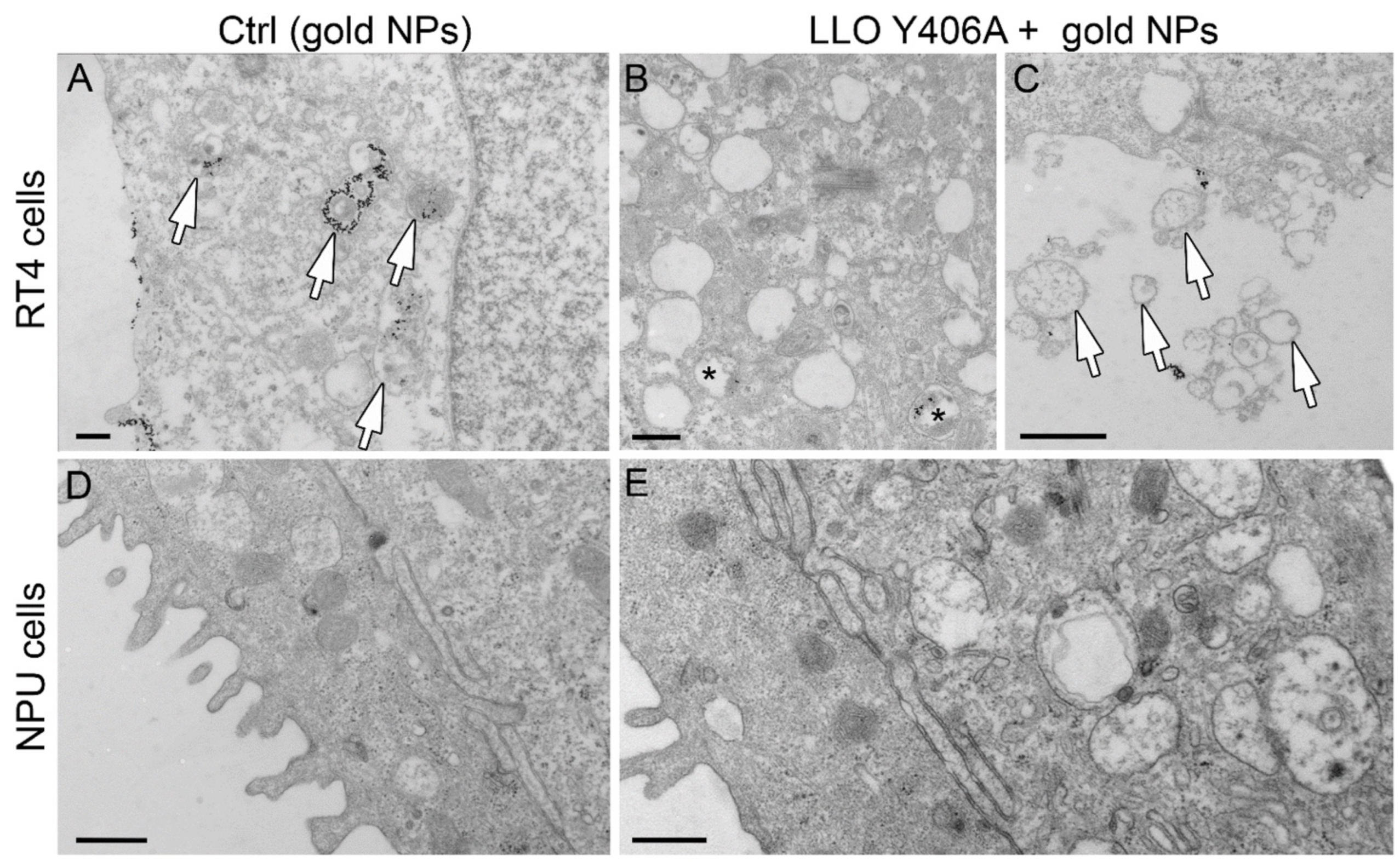
2.2. LLO Y406A Does Not Cause Endosomal Escape and Is Mainly Located at He Plasma Membrane
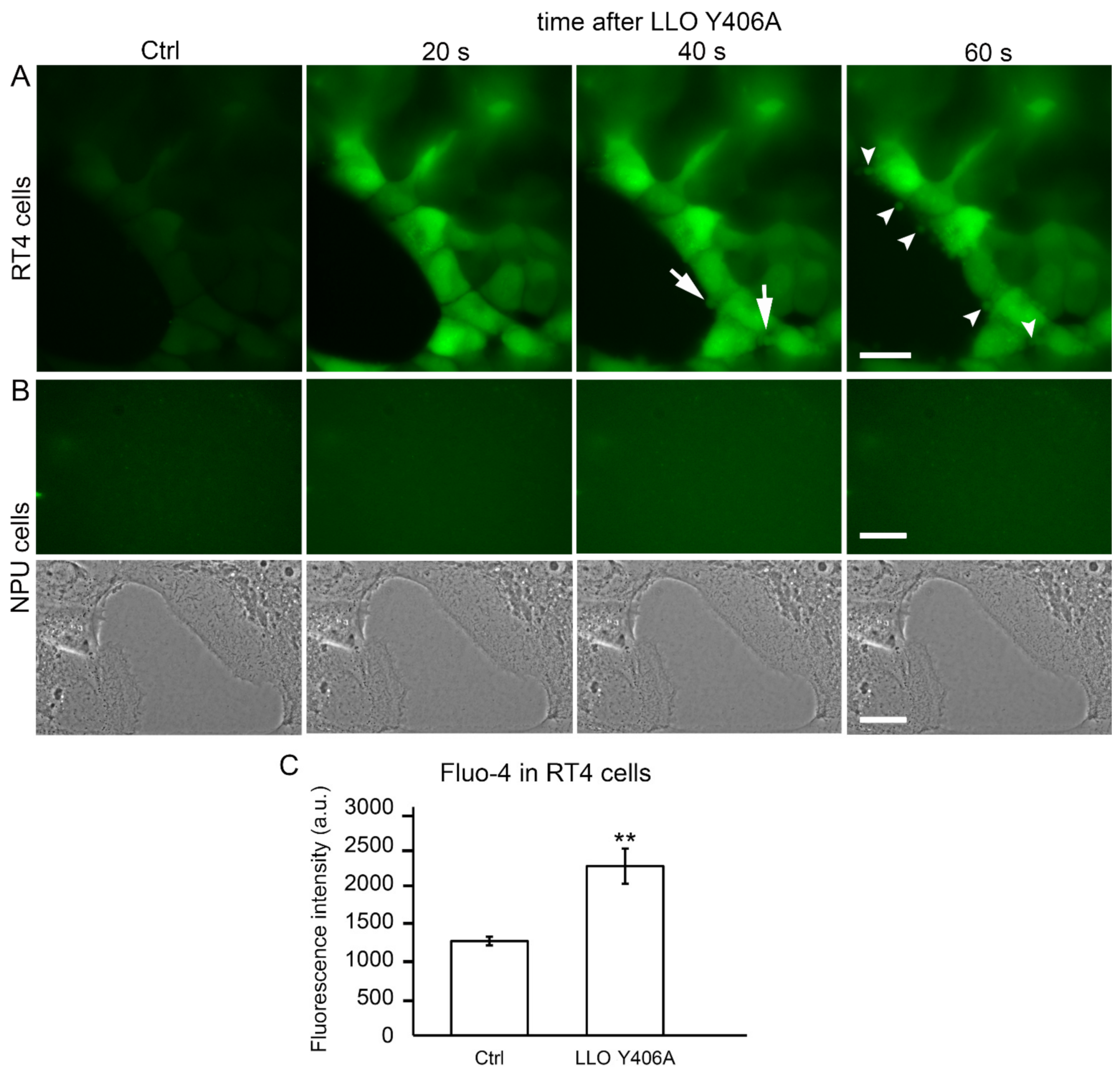
2.3. LLO Y406A Induces Calcium Influx and Cell Blebbing in RT4 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Cell Viability Measurement
4.3. Preparation, Expression and Purification of Recombinant Proteins
4.4. Endosomal Escape
4.5. Immunolabeling
4.6. Fluo-4 AM Calcium Staining
4.7. Generation of Gold Nanoparticles
4.8. Transmission Electron Microscopy
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Richters, A.; Aben, K.K.H.; Kiemeney, L. The global burden of urinary bladder cancer: An update. World J. Urol. 2020, 38, 1895–1904. [Google Scholar] [CrossRef]
- Kirkali, Z. The future of oncological urology in Europe: Are we prepared? Eur. Urol. 2006, 49, 11–12. [Google Scholar] [CrossRef] [PubMed]
- Vadia, S.; Seveau, S. Fluxes of Ca2+ and K+ are required for the listeriolysin O-dependent internalization pathway of Listeria monocytogenes. Infect. Immun. 2014, 82, 1084–1091. [Google Scholar] [CrossRef]
- Mostowy, S.; Cossart, P. Virulence factors that modulate the cell biology of Listeria infection and the host response. Adv. Immunol. 2012, 113, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Alberti-Segui, C.; Goeden, K.R.; Higgins, D.E. Differential function of Listeria monocytogenes listeriolysin O and phospholipases C in vacuolar dissolution following cell-to-cell spread. Cell. Microbiol. 2007, 9, 179–195. [Google Scholar] [CrossRef]
- Coconnier, M.H.; Lorrot, M.; Barbat, A.; Laboisse, C.; Servin, A.L. Listeriolysin O-induced stimulation of mucin exocytosis in polarized intestinal mucin-secreting cells: Evidence for toxin recognition of membrane-associated lipids and subsequent toxin internalization through caveolae. Cell. Microbiol. 2000, 2, 487–504. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.A.; Ribet, D.; Stavru, F.; Cossart, P. Listeriolysin O: The Swiss army knife of Listeria. Trends Microbiol. 2012, 20, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Seveau, S. Multifaceted activity of listeriolysin O, the cholesterol-dependent cytolysin of Listeria monocytogenes. Subcell. Biochem. 2014, 80, 161–195. [Google Scholar] [CrossRef]
- Bavdek, A.; Gekara, N.O.; Priselac, D.; Gutierrez Aguirre, I.; Darji, A.; Chakraborty, T.; Macek, P.; Lakey, J.H.; Weiss, S.; Anderluh, G. Sterol and pH interdependence in the binding, oligomerization, and pore formation of Listeriolysin O. Biochemistry 2007, 46, 4425–4437. [Google Scholar] [CrossRef]
- Kisovec, M.; Rezelj, S.; Knap, P.; Cajnko, M.M.; Caserman, S.; Flašker, A.; Žnidaršič, N.; Repič, M.; Mavri, J.; Ruan, Y.; et al. Engineering a pH responsive pore forming protein. Sci. Rep. 2017, 7, 42231. [Google Scholar] [CrossRef] [PubMed]
- Schuerch, D.W.; Wilson-Kubalek, E.M.; Tweten, R.K. Molecular basis of listeriolysin O pH dependence. Proc. Natl. Acad. Sci. USA 2005, 102, 12537–12542. [Google Scholar] [CrossRef]
- Bavdek, A.; Kostanjsek, R.; Antonini, V.; Lakey, J.H.; Dalla Serra, M.; Gilbert, R.J.; Anderluh, G. pH dependence of listeriolysin O aggregation and pore-forming ability. FEBS J. 2012, 279, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Schnupf, P.; Portnoy, D.A. Listeriolysin O: A phagosome-specific lysin. Microbes Infect. 2007, 9, 1176–1187. [Google Scholar] [CrossRef]
- Vadia, S.; Arnett, E.; Haghighat, A.C.; Wilson-Kubalek, E.M.; Tweten, R.K.; Seveau, S. The pore-forming toxin listeriolysin O mediates a novel entry pathway of L. monocytogenes into human hepatocytes. PLoS Pathog. 2011, 7, e1002356. [Google Scholar] [CrossRef]
- Podobnik, M.; Marchioretto, M.; Zanetti, M.; Bavdek, A.; Kisovec, M.; Cajnko, M.M.; Lunelli, L.; Dalla Serra, M.; Anderluh, G. Plasticity of listeriolysin O pores and its regulation by pH and unique histidine [corrected]. Sci. Rep. 2015, 5, 9623. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.; Cabanes, D.; Sarmento Mesquita, F.; Sousa, S. Mechanisms protecting host cells against bacterial pore-forming toxins. Cell. Mol. Life Sci. 2019, 76, 1319–1339. [Google Scholar] [CrossRef] [PubMed]
- Etxaniz, A.; Gonzalez-Bullon, D.; Martin, C.; Ostolaza, H. Membrane Repair Mechanisms against Permeabilization by Pore-Forming Toxins. Toxins 2018, 10, 234. [Google Scholar] [CrossRef]
- Nomura, T.; Kawamura, I.; Kohda, C.; Baba, H.; Ito, Y.; Kimoto, T.; Watanabe, I.; Mitsuyama, M. Irreversible loss of membrane-binding activity of Listeria-derived cytolysins in non-acidic conditions: A distinct difference from allied cytolysins produced by other Gram-positive bacteria. Microbiology 2007, 153, 2250–2258. [Google Scholar] [CrossRef]
- Plaza-Ga, I.; Manzaneda-González, V.; Kisovec, M.; Almendro-Vedia, V.; Muñoz-Úbeda, M.; Anderluh, G.; Guerrero-Martínez, A.; Natale, P.; López Montero, I. pH-triggered endosomal escape of pore-forming Listeriolysin O toxin-coated gold nanoparticles. J. Nanobiotechnol. 2019, 17, 108. [Google Scholar] [CrossRef]
- Watanabe, I.; Nomura, T.; Tominaga, T.; Yamamoto, K.; Kohda, C.; Kawamura, I.; Mitsuyama, M. Dependence of the lethal effect of pore-forming haemolysins of Gram-positive bacteria on cytolytic activity. J. Med. Microbiol. 2006, 55, 505–510. [Google Scholar] [CrossRef]
- Malet, J.K.; Cossart, P.; Ribet, D. Alteration of epithelial cell lysosomal integrity induced by bacterial cholesterol-dependent cytolysins. Cell. Microbiol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Carbayo, M.; Socci, N.D.; Charytonowicz, E.; Lu, M.; Prystowsky, M.; Childs, G.; Cordon-Cardo, C. Molecular profiling of bladder cancer using cDNA microarrays: Defining histogenesis and biological phenotypes. Cancer Res. 2002, 62, 6973–6980. [Google Scholar]
- Kreft, M.E.; Di Giandomenico, D.; Beznoussenko, G.V.; Resnik, N.; Mironov, A.A.; Jezernik, K. Golgi apparatus fragmentation as a mechanism responsible for uniform delivery of uroplakins to the apical plasma membrane of uroepithelial cells. Biol. Cell 2010, 102, 593–607. [Google Scholar] [CrossRef]
- Resnik, N.; Prezelj, T.; De Luca, G.M.R.; Manders, E.; Polishchuk, R.; Veranič, P.; Kreft, M.E. Helical organization of microtubules occurs in a minority of tunneling membrane nanotubes in normal and cancer urothelial cells. Sci. Rep. 2018, 8, 17133. [Google Scholar] [CrossRef]
- Resnik, N.; Repnik, U.; Kreft, M.E.; Sepčić, K.; Maček, P.; Turk, B.; Veranič, P. Highly Selective Anti-Cancer Activity of Cholesterol-Interacting Agents Methyl-β-Cyclodextrin and Ostreolysin A/Pleurotolysin B Protein Complex on Urothelial Cancer Cells. PLoS ONE 2015, 10, e0137878. [Google Scholar] [CrossRef]
- Višnjar, T.; Kreft, M.E. The complete functional recovery of chitosan-treated biomimetic hyperplastic and normoplastic urothelial models. Histochem. Cell Biol. 2015, 143, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Lojk, J.; Bregar, V.B.; Strojan, K.; Hudoklin, S.; Veranič, P.; Pavlin, M.; Kreft, M.E. Increased endocytosis of magnetic nanoparticles into cancerous urothelial cells versus normal urothelial cells. Histochem. Cell Biol. 2018, 149, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, S.R.; Yoon, H.Y.; Chang, I.H.; Whang, Y.M.; Cho, M.J.; Kim, M.J.; Kim, S.Y.; Lee, S.J.; Choi, Y.W. Poloxamer 407 Hydrogels for Intravesical Instillation to Mouse Bladder: Gel-Forming Capacity and Retention Performance. Korean J. Urol. Oncol. 2017, 15, 178–186. [Google Scholar] [CrossRef]
- Hayat, M.A. Colloidal Gold: Principles, Methods, and Applications; Academic Press: San Diego, CA, USA, 1989; Volume 2–3. [Google Scholar]
- Smith, G.A.; Marquis, H.; Jones, S.; Johnston, N.C.; Portnoy, D.A.; Goldfine, H. The two distinct phospholipases C of Listeria monocytogenes have overlapping roles in escape from a vacuole and cell-to-cell spread. Infect. Immun. 1995, 63, 4231–4237. [Google Scholar] [CrossRef] [PubMed]
- Dramsi, S.; Cossart, P. Listeriolysin O-mediated calcium influx potentiates entry of Listeria monocytogenes into the human Hep-2 epithelial cell line. Infect. Immun. 2003, 71, 3614–3618. [Google Scholar] [CrossRef]
- Nygard Skalman, L.; Holst, M.R.; Larsson, E.; Lundmark, R. Plasma membrane damage caused by listeriolysin O is not repaired through endocytosis of the membrane pore. Biol. Open 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Cajnko, M.M.; Marušić, M.; Kisovec, M.; Rojko, N.; Benčina, M.; Caserman, S.; Anderluh, G. Listeriolysin O Affects the Permeability of Caco-2 Monolayer in a Pore-Dependent and Ca2+-Independent Manner. PLoS ONE 2015, 10, e0130471. [Google Scholar] [CrossRef] [PubMed]
- Repp, H.; Pamukci, Z.; Koschinski, A.; Domann, E.; Darji, A.; Birringer, J.; Brockmeier, D.; Chakraborty, T.; Dreyer, F. Listeriolysin of Listeria monocytogenes forms Ca2+-permeable pores leading to intracellular Ca2+ oscillations. Cell. Microbiol. 2002, 4, 483–491. [Google Scholar] [CrossRef]
- Stachowiak, R.; Łyżniak, M.; Grabowska, M.; Roeske, K.; Jagielski, T.; Bielecki, J.; Budziszewska, B.K.; Hoser, G.; Kawiak, J. Cytotoxicity of purified listeriolysin O on mouse and human leukocytes and leukaemia cells. BMC Biotechnol. 2014, 14, 77. [Google Scholar] [CrossRef]
- Sun, R.; Liu, Y. Listeriolysin O as a strong immunogenic molecule for the development of new anti-tumor vaccines. Hum. Vaccines Immunother. 2013, 9, 1058–1068. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Bae, D.; Wang, C. Listeriolysin O mediates cytotoxicity against human brain microvascular endothelial cells. FEMS Microbiol. Lett. 2015, 362, fnv084. [Google Scholar] [CrossRef]
- Gatenby, R.A.; Gillies, R.J. Why do cancers have high aerobic glycolysis? Nat. Rev. Cancer 2004, 4, 891–899. [Google Scholar] [CrossRef]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Massari, F.; Ciccarese, C.; Santoni, M.; Iacovelli, R.; Mazzucchelli, R.; Piva, F.; Scarpelli, M.; Berardi, R.; Tortora, G.; Lopez-Beltran, A.; et al. Metabolic phenotype of bladder cancer. Cancer Treat. Rev. 2016, 45, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Swietach, P.; Vaughan-Jones, R.D.; Harris, A.L. Regulation of tumor pH and the role of carbonic anhydrase 9. Cancer Metastasis Rev. 2007, 26, 299–310. [Google Scholar] [CrossRef]
- Fischer, K.; Hoffmann, P.; Voelkl, S.; Meidenbauer, N.; Ammer, J.; Edinger, M.; Gottfried, E.; Schwarz, S.; Rothe, G.; Hoves, S.; et al. Inhibitory effect of tumor cell-derived lactic acid on human T cells. Blood 2007, 109, 3812–3819. [Google Scholar] [CrossRef] [PubMed]
- Visnjar, T.; Romih, R.; Zupancic, D. Lectins as possible tools for improved urinary bladder cancer management. Glycobiology 2019, 29, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Zupancic, D.; Kreft, M.E.; Romih, R. Selective binding of lectins to normal and neoplastic urothelium in rat and mouse bladder carcinogenesis models. Protoplasma 2014, 251, 49–59. [Google Scholar] [CrossRef]
- Shewell, L.K.; Day, C.J.; Jen, F.E.; Haselhorst, T.; Atack, J.M.; Reijneveld, J.F.; Everest-Dass, A.; James, D.B.A.; Boguslawski, K.M.; Brouwer, S.; et al. All major cholesterol-dependent cytolysins use glycans as cellular receptors. Sci. Adv. 2020, 6, eaaz4926. [Google Scholar] [CrossRef]
- Chen, H.; Konofagou, E.E. The size of blood-brain barrier opening induced by focused ultrasound is dictated by the acoustic pressure. J. Cereb. Blood Flow Metab. 2014, 34, 1197–1204. [Google Scholar] [CrossRef]
- Atanassoff, A.P.; Wolfmeier, H.; Schoenauer, R.; Hostettler, A.; Ring, A.; Draeger, A.; Babiychuk, E.B. Microvesicle shedding and lysosomal repair fulfill divergent cellular needs during the repair of streptolysin O-induced plasmalemmal damage. PLoS ONE 2014, 9, e89743. [Google Scholar] [CrossRef] [PubMed]
- Wolfmeier, H.; Radecke, J.; Schoenauer, R.; Koeffel, R.; Babiychuk, V.S.; Drucker, P.; Hathaway, L.J.; Mitchell, T.J.; Zuber, B.; Draeger, A.; et al. Active release of pneumolysin prepores and pores by mammalian cells undergoing a Streptococcus pneumoniae attack. Biochim. Biophys. Acta 2016, 1860, 2498–2509. [Google Scholar] [CrossRef]
- Keyel, P.A.; Loultcheva, L.; Roth, R.; Salter, R.D.; Watkins, S.C.; Yokoyama, W.M.; Heuser, J.E. Streptolysin O clearance through sequestration into blebs that bud passively from the plasma membrane. J. Cell Sci. 2011, 124, 2414–2423. [Google Scholar] [CrossRef]
- Monastyrskaya, K.; Babiychuk, E.B.; Draeger, A.; Burkhard, F.C. Down-regulation of annexin A1 in the urothelium decreases cell survival after bacterial toxin exposure. J. Urol. 2013, 190, 325–333. [Google Scholar] [CrossRef]
- Coelho, C.; Brown, L.; Maryam, M.; Vij, R.; Smith, D.F.Q.; Burnet, M.C.; Kyle, J.E.; Heyman, H.M.; Ramirez, J.; Prados-Rosales, R.; et al. virulence factors, including listeriolysin O, are secreted in biologically active extracellular vesicles. J. Biol. Chem. 2019, 294, 1202–1217. [Google Scholar] [CrossRef] [PubMed]
- Višnjar, T.; Chesi, G.; Iacobacci, S.; Polishchuk, E.; Resnik, N.; Robenek, H.; Kreft, M.; Romih, R.; Polishchuk, R.; Kreft, M.E. Uroplakin traffic through the Golgi apparatus induces its fragmentation: New insights from novel in vitro models. Sci. Rep. 2017, 7, 12842. [Google Scholar] [CrossRef]
- Cho, I.C.; Kim, E.K.; Joung, J.Y.; Seo, H.K.; Chung, J.; Park, W.S.; Lee, K.H. Adjuvant intravesical instillation for primary T1G3 bladder cancer: BCG versus MMC in Korea. Anticancer Res. 2012, 32, 1493–1498. [Google Scholar] [PubMed]
- Smith, J.A.; Labasky, R.F.; Cockett, A.T.K.; Fracchia, J.A.; Montie, J.E.; Rowland, R.G. Bladder cancer clinical guidelines panel summary report on the management of nonmuscle invasive bladder cancer (stages Ta, T1 and TIS). J. Urol. 1999, 162, 1697–1701. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Resnik, N.; Tratnjek, L.; Kreft, M.E.; Kisovec, M.; Aden, S.; Bedina Zavec, A.; Anderluh, G.; Podobnik, M.; Veranič, P. Cytotoxic Activity of LLO Y406A Is Targeted to the Plasma Membrane of Cancer Urothelial Cells. Int. J. Mol. Sci. 2021, 22, 3305. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073305
Resnik N, Tratnjek L, Kreft ME, Kisovec M, Aden S, Bedina Zavec A, Anderluh G, Podobnik M, Veranič P. Cytotoxic Activity of LLO Y406A Is Targeted to the Plasma Membrane of Cancer Urothelial Cells. International Journal of Molecular Sciences. 2021; 22(7):3305. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073305
Chicago/Turabian StyleResnik, Nataša, Larisa Tratnjek, Mateja Erdani Kreft, Matic Kisovec, Saša Aden, Apolonija Bedina Zavec, Gregor Anderluh, Marjetka Podobnik, and Peter Veranič. 2021. "Cytotoxic Activity of LLO Y406A Is Targeted to the Plasma Membrane of Cancer Urothelial Cells" International Journal of Molecular Sciences 22, no. 7: 3305. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073305