
Effects of Environmental Conditions on Nephron Number: Modeling Maternal Disease and Epigenetic Regulation in Renal Development

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
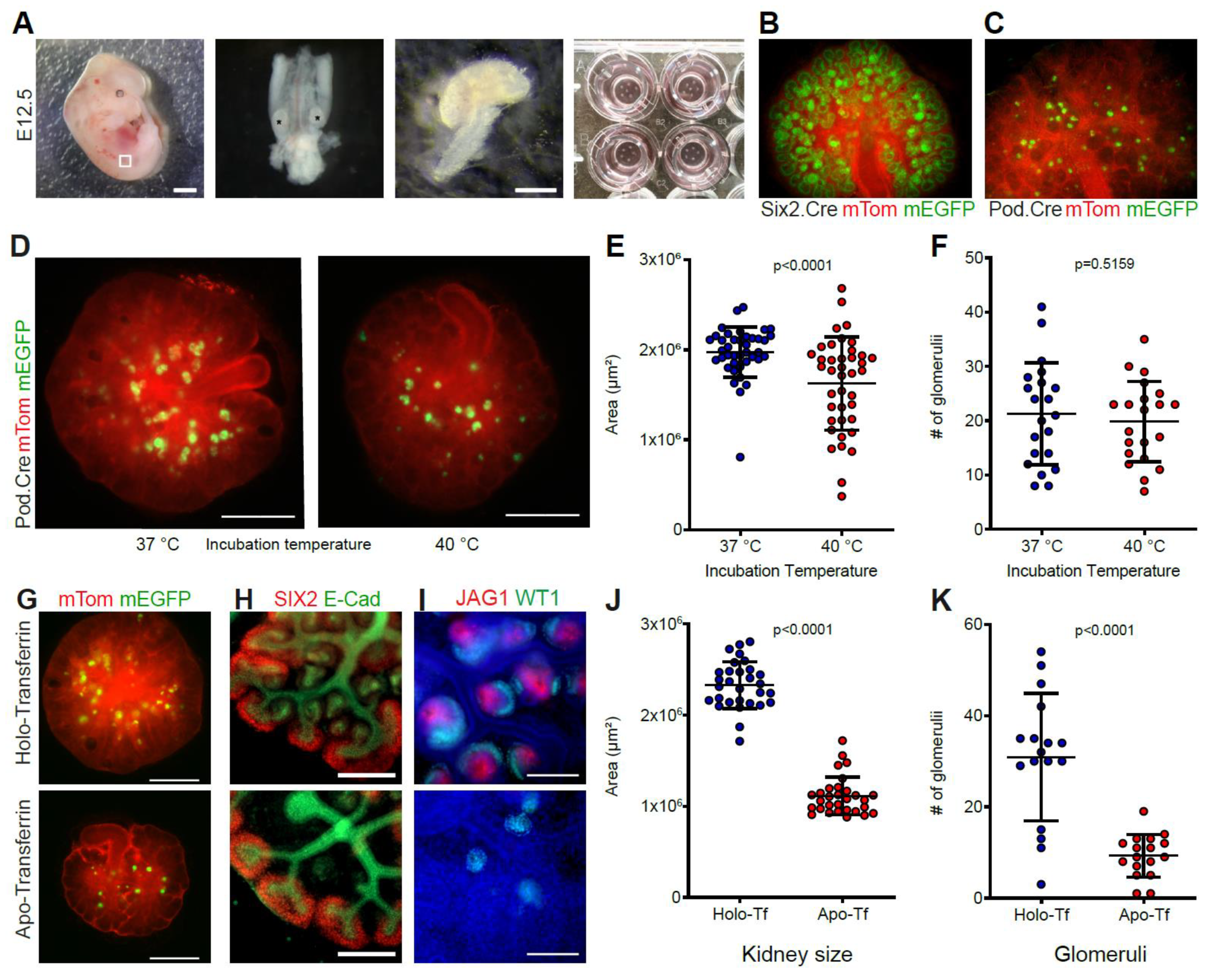
2.1. Use of Metanephric Organ Cultures to Study the Effect of Environmental Conditions on Renal Development
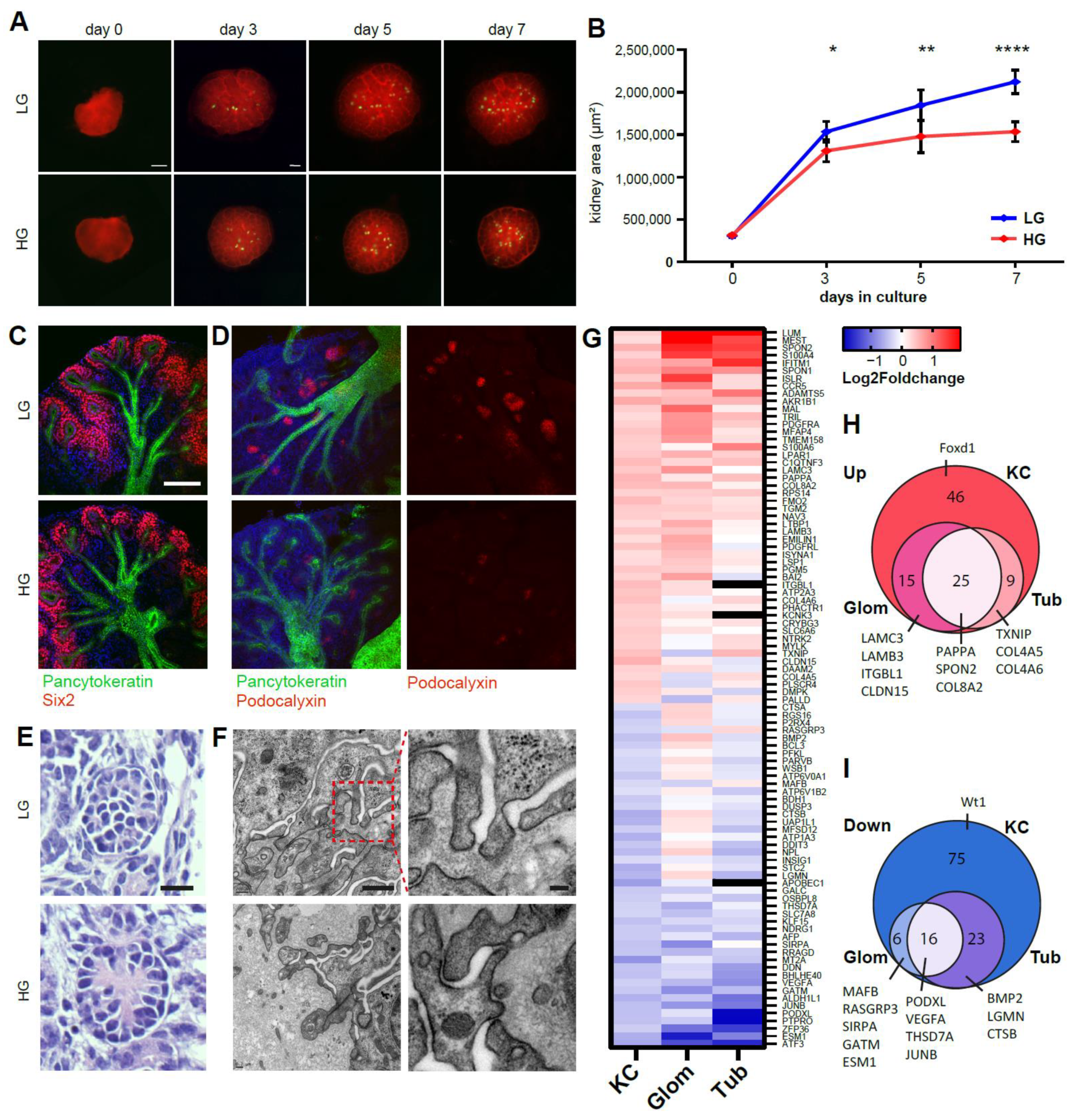
2.2. Ex Vivo High Glucose Exposure Leads to Diabetic Nephropathy-Associated Changes in the Developing Kidney
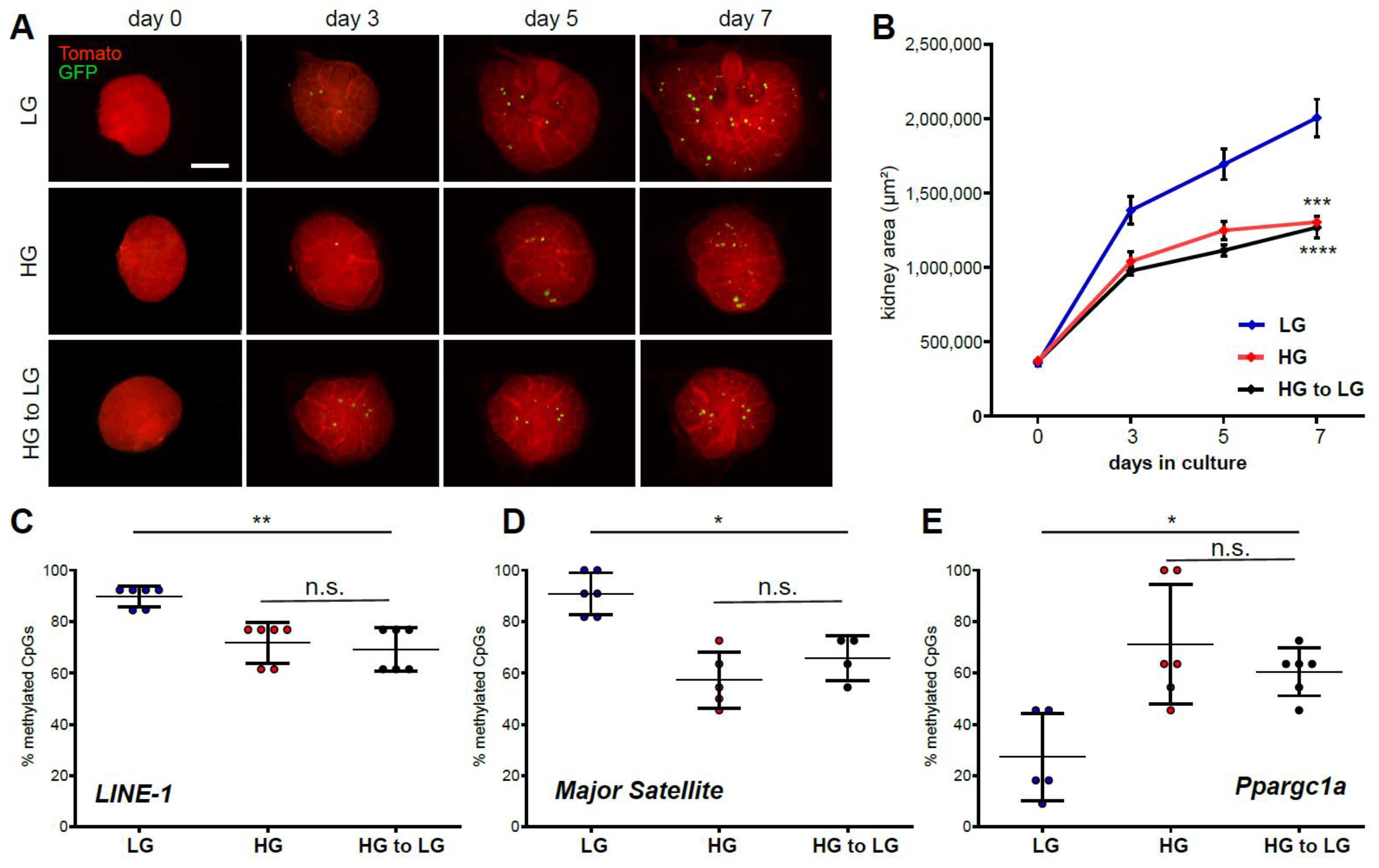
2.3. Ex Vivo High Glucose Exposure Influences to Long-Term Memory Formation via DNA Methylation
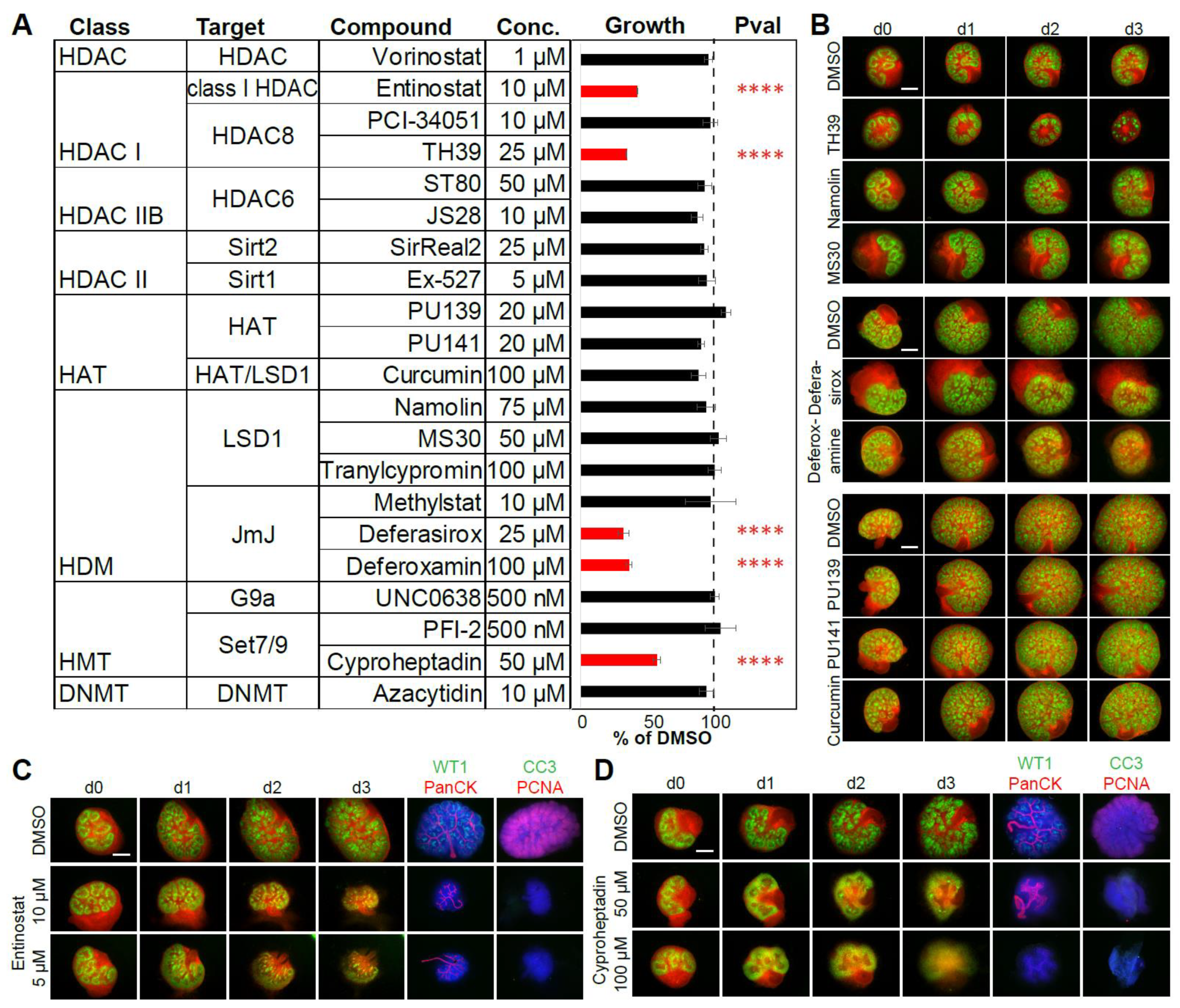
2.4. Ex Vivo Small Compound Screen Identifies Epigenetic Regulators of Renal Development
3. Discussion
4. Materials and Methods
4.1. Animal Handling
4.2. Timed Harvest of Embryos/Microdissection/Culture of Metanephroi
4.3. Genotyping
4.4. Hyperglycemic Conditions
4.5. Hyperthermic Conditions
4.6. Iron-Restriction
4.7. Whole Mount Immunofluorescence Staining of Explants
4.8. Imaging
4.9. Histology
4.10. EM
4.11. Bisulfite-PCR
4.12. RNA Sequencing
4.13. Microarray Analysis of Human Kidney Biopsies
4.14. qPCR
4.15. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J. The fetal and infant origins of adult disease. BMJ 1990, 301, 1111. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Garrett, M.R. Nephron number, hypertension, and CKD: Physiological and genetic insight from humans and animal models. Physiol. Genom. 2017, 49, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Hughson, M.D.; Douglas-Denton, R.; Bertram, J.F.; Hoy, W.E. Hypertension, glomerular number, and birth weight in African Americans and white subjects in the southeastern United States. Kidney Int. 2006, 69, 671–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, G.; Zimmer, G.; Mall, G.; Ritz, E.; Amann, K. Nephron number in patients with primary hypertension. N. Engl. J. Med. 2003, 348, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lelievre-Pegorier, M.; Vilar, J.; Ferrier, M.L.; Moreau, E.; Freund, N.; Gilbert, T.; Merlet-Benichou, C. Mild vitamin A deficiency leads to inborn nephron deficit in the rat. Kidney Int. 1998, 54, 1455–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley-Evans, S.C.; Welham, S.J.; Jackson, A.A. Fetal exposure to a maternal low protein diet impairs nephrogenesis and promotes hypertension in the rat. Life Sci. 1999, 64, 965–974. [Google Scholar] [CrossRef]
- Singh, R.R.; Moritz, K.M.; Bertram, J.F.; Cullen-McEwen, L.A. Effects of dexamethasone exposure on rat metanephric development: In vitro and in vivo studies. Am. J. Physiol. Renal. Physiol. 2007, 293, F548–F554. [Google Scholar] [CrossRef] [Green Version]
- Cain, J.E.; Di Giovanni, V.; Smeeton, J.; Rosenblum, N.D. Genetics of renal hypoplasia: Insights into the mechanisms controlling nephron endowment. Pediatr. Res. 2010, 68, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.M.; Forhead, A.J.; Petry, C.J.; Ozanne, S.E.; Hales, C.N. Long-term programming of blood pressure by maternal dietary iron restriction in the rat. Br. J. Nutr. 2002, 88, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Hokke, S.N.; Armitage, J.A.; Puelles, V.G.; Short, K.M.; Jones, L.; Smyth, I.M.; Bertram, J.F.; Cullen-McEwen, L.A. Altered ureteric branching morphogenesis and nephron endowment in offspring of diabetic and insulin-treated pregnancy. PLoS ONE 2013, 8, e58243. [Google Scholar]
- Hughson, M.; Farris, A.B., 3rd; Douglas-Denton, R.; Hoy, W.E.; Bertram, J.F. Glomerular number and size in autopsy kidneys: The relationship to birth weight. Kidney Int. 2003, 63, 2113–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, G.; Fox, U.; Di Francesco, G.F.; Giovanetti, A.M.; Pagetti, D. Blood pressure changes produced by kidney cross-transplantation between spontaneously hypertensive rats and normotensive rats. Clin. Sci. Mol. Med. 1974, 47, 435–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trowell, O.A. A modified technique for organ culture in vitro. Exp. Cell Res. 1954, 6, 246–248. [Google Scholar] [CrossRef]
- Saxen, L.; Lehtonen, E. Embryonic kidney in organ culture. Differentiation 1987, 36, 2–11. [Google Scholar] [CrossRef]
- Andersen, A.M.; Vastrup, P.; Wohlfahrt, J.; Andersen, P.K.; Olsen, J.; Melbye, M. Fever in pregnancy and risk of fetal death: A cohort study. Lancet 2002, 360, 1552–1556. [Google Scholar] [CrossRef]
- Edwards, M.J. Hyperthermia as a teratogen: A review of experimental studies and their clinical significance. Teratog. Carcinog. Mutagen. 1986, 6, 563–582. [Google Scholar] [CrossRef] [PubMed]
- Dreier, J.W.; Andersen, A.M.; Berg-Beckhoff, G. Systematic review and meta-analyses: Fever in pregnancy and health impacts in the offspring. Pediatrics 2014, 133, e674–e688. [Google Scholar] [CrossRef] [Green Version]
- Waller, D.K.; Hashmi, S.S.; Hoyt, A.T.; Duong, H.T.; Tinker, S.C.; Gallaway, M.S.; Olney, R.S.; Finnell, R.H.; Hecht, J.T.; Canfield, M.A.; et al. Maternal report of fever from cold or flu during early pregnancy and the risk for noncardiac birth defects, National Birth Defects Prevention Study, 1997-2011. Birth Defects Res. 2018, 110, 342–351. [Google Scholar] [CrossRef]
- Abe, K.; Honein, M.A.; Moore, C.A. Maternal febrile illnesses, medication use, and the risk of congenital renal anomalies. Birth Defects Res. A Clin. Mol. Teratol. 2003, 67, 911–918. [Google Scholar] [CrossRef]
- Edwards, M.J.; Saunders, R.D.; Shiota, K. Effects of heat on embryos and foetuses. Int. J. Hyperthermia 2003, 19, 295–324. [Google Scholar] [CrossRef]
- Edwards, M.J. Congenital defects in guinea pigs: Fetal resorptions, abortions, and malformations following induced hyperthermia during early gestation. Teratology 1969, 2, 313–328. [Google Scholar] [CrossRef]
- Black, R.E.; Victora, C.G.; Walker, S.P.; Bhutta, Z.A.; Christian, P.; de Onis, M.; Ezzati, M.; Grantham-McGregor, S.; Katz, J.; Martorell, R.; et al. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 427–451. [Google Scholar] [CrossRef]
- Lisle, S.J.; Lewis, R.M.; Petry, C.J.; Ozanne, S.E.; Hales, C.N.; Forhead, A.J. Effect of maternal iron restriction during pregnancy on renal morphology in the adult rat offspring. Br. J. Nutr. 2003, 90, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.Y.; Woolley, J.C.; Blohowiak, S.E.; Smith, Z.R.; Siddappa, A.M.; Magness, R.R.; Kling, P.J. Dietary-induced gestational iron deficiency inhibits postnatal tissue iron delivery and postpones the cessation of active nephrogenesis in rats. Reprod. Fertil. Dev. 2016, 29, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Guariguata, L.; Linnenkamp, U.; Beagley, J.; Whiting, D.R.; Cho, N.H. Global estimates of the prevalence of hyperglycaemia in pregnancy. Diabetes Res. Clin. Pract. 2014, 103, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.; Chen, Y.W.; Chenier, I.; Chan, J.S.; Quaggin, S.; Hebert, M.J.; Ingelfinger, J.R.; Zhang, S.L. Maternal diabetes modulates renal morphogenesis in offspring. J. Am. Soc. Nephrol. 2008, 19, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Amri, K.; Freund, N.; Vilar, J.; Merlet-Benichou, C.; Lelievre-Pegorier, M. Adverse effects of hyperglycemia on kidney development in rats: In vivo and in vitro studies. Diabetes 1999, 48, 2240–2245. [Google Scholar] [CrossRef]
- Kanwar, Y.S.; Liu, Z.Z.; Kumar, A.; Usman, M.I.; Wada, J.; Wallner, E.I. D-glucose-induced dysmorphogenesis of embryonic kidney. J. Clin. Investig. 1996, 98, 2478–2488. [Google Scholar] [CrossRef] [Green Version]
- Kanwar, Y.S.; Akagi, S.; Nayak, B.; Sun, L.; Wada, J.; Xie, P.; Thakur, A.; Chugh, S.S.; Danesh, F.R. Renal-specific oxidoreductase biphasic expression under high glucose ambience during fetal versus neonatal development. Kidney Int. 2005, 68, 1670–1683. [Google Scholar] [CrossRef] [Green Version]
- Wanner, N.; Vornweg, J.; Combes, A.; Wilson, S.; Plappert, J.; Rafflenbeul, G.; Puelles, V.G.; Rahman, R.U.; Liwinski, T.; Lindner, S.; et al. DNA Methyltransferase 1 Controls Nephron Progenitor Cell Renewal and Differentiation. J. Am. Soc. Nephrol. 2019, 30, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Osterby, R.; Gundersen, H.J. Glomerular size and structure in diabetes mellitus. I. Early abnormalities. Diabetologia 1975, 11, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Mac-Moune Lai, F.; Szeto, C.C.; Choi, P.C.; Ho, K.K.; Tang, N.L.; Chow, K.M.; Li, P.K.; To, K.F. Isolate diffuse thickening of glomerular capillary basement membrane: A renal lesion in prediabetes? Mod. Pathol. 2004, 17, 1506–1512. [Google Scholar] [CrossRef] [PubMed]
- Al Shawaf, E.; Abu-Farha, M.; Devarajan, S.; Alsairafi, Z.; Al-Khairi, I.; Cherian, P.; Ali, H.; Mathur, A.; Al-Mulla, F.; Al Attar, A.; et al. ANGPTL4: A Predictive Marker for Diabetic Nephropathy. J. Diabetes Res. 2019, 2019, 4943191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakoshi, M.; Tanimoto, M.; Gohda, T.; Hagiwara, S.; Takagi, M.; Horikoshi, S.; Tomino, Y. Mindin: A novel marker for podocyte injury in diabetic nephropathy. Nephrol. Dial. Transpl. 2011, 26, 2153–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donegan, D.; Bale, L.K.; Conover, C.A. PAPP-A in normal human mesangial cells: Effect of inflammation and factors related to diabetic nephropathy. J. Endocrinol. 2016, 231, 71–80. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, Y.; Kelly, D.J.; Tan, C.Y.; Gill, A.; Cheng, D.; Braet, F.; Park, J.S.; Sue, C.M.; Pollock, C.A.; et al. Thioredoxin interacting protein (TXNIP) regulates tubular autophagy and mitophagy in diabetic nephropathy through the mTOR signaling pathway. Sci. Rep. 2016, 6, 29196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randhawa, P.K.; Rylova, S.; Heinz, J.Y.; Kiser, S.; Fried, J.H.; Dunworth, W.P.; Anderson, A.L.; Barber, A.T.; Chappell, J.C.; Roberts, D.M.; et al. The Ras activator RasGRP3 mediates diabetes-induced embryonic defects and affects endothelial cell migration. Circ. Res. 2011, 108, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Han, D.; Yu, P.; Huang, Q.; Ge, P. Genome-scale transcriptional analysis reveals key genes associated with the development of type II diabetes in mice. Exp. Ther. Med. 2017, 13, 1044–1150. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Soroush, F.; Long, J.; Hall, E.T.; Adishesha, P.K.; Bhattacharya, S.; Kiani, M.F.; Bhalla, V. Murine glomerular transcriptome links endothelial cell-specific molecule-1 deficiency with susceptibility to diabetic nephropathy. PLoS ONE 2017, 12, e0185250. [Google Scholar] [CrossRef]
- Doan, T.N.A.; Briffa, J.F.; Phillips, A.L.; Leemaqz, S.Y.; Burton, R.A.; Romano, T.; Wlodek, M.E.; Bianco-Miotto, T. Epigenetic mechanisms involved in intrauterine growth restriction and aberrant kidney development and function. J. Dev. Orig. Health Dis. 2020, 1–11. [Google Scholar] [CrossRef]
- Huang, B.; Liu, Z.; Vonk, A.; Zeng, Z.; Li, Z. Epigenetic regulation of kidney progenitor cells. Stem Cells Transl. Med. 2020, 9, 655–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Dahr, S.S.; Saifudeen, Z. Epigenetic regulation of renal development. Semin. Cell Dev. Biol. 2019, 91, 111–118. [Google Scholar] [CrossRef]
- Liu, H.; Hilliard, S.; Kelly, E.; Chen, C.H.; Saifudeen, Z.; El-Dahr, S.S. The polycomb proteins EZH1 and EZH2 co-regulate chromatin accessibility and nephron progenitor cell lifespan in mice. J. Biol. Chem. 2020, 295, 11542–11558. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Yao, X.; Li, Y.; Chen, C.H.; Liu, J.; Saifudeen, Z.; El-Dahr, S.S. Histone deacetylases 1 and 2 regulate the transcriptional programs of nephron progenitors and renal vesicles. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Ettou, S.; Khalid, M.; Taglienti, M.; Jain, D.; Jung, Y.L.; Seager, C.; Liu, Y.; Ng, K.H.; Park, P.J.; et al. EED, a member of the polycomb group, is required for nephron differentiation and the maintenance of nephron progenitor cells. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumpf, T.; Schiedel, M.; Karaman, B.; Roessler, C.; North, B.J.; Lehotzky, A.; Olah, J.; Ladwein, K.I.; Schmidtkunz, K.; Gajer, M.; et al. Selective Sirt2 inhibition by ligand-induced rearrangement of the active site. Nat. Commun. 2015, 6, 6263. [Google Scholar] [CrossRef]
- Senger, J.; Melesina, J.; Marek, M.; Romier, C.; Oehme, I.; Witt, O.; Sippl, W.; Jung, M. Synthesis and Biological Investigation of Oxazole Hydroxamates as Highly Selective Histone Deacetylase 6 (HDAC6) Inhibitors. J. Med. Chem. 2016, 59, 1545–1555. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, Y.; Ito, A.; Niwa, H.; Okamura, M.; Fujiwara, T.; Hirano, T.; Handa, N.; Umehara, T.; Sonoda, T.; Ogawa, K.; et al. Identification of Cyproheptadine as an Inhibitor of SET Domain Containing Lysine Methyltransferase 7/9 (Set7/9) That Regulates Estrogen-Dependent Transcription. J. Med. Chem. 2016, 59, 3650–3660. [Google Scholar] [CrossRef] [PubMed]
- Barsyte-Lovejoy, D.; Li, F.; Oudhoff, M.J.; Tatlock, J.H.; Dong, A.; Zeng, H.; Wu, H.; Freeman, S.A.; Schapira, M.; Senisterra, G.A.; et al. (R)-PFI-2 is a potent and selective inhibitor of SETD7 methyltransferase activity in cells. Proc. Natl. Acad. Sci. USA 2014, 111, 12853–12858. [Google Scholar] [CrossRef] [Green Version]
- Roatsch, M.; Hoffmann, I.; Abboud, M.I.; Hancock, R.L.; Tarhonskaya, H.; Hsu, K.F.; Wilkins, S.E.; Yeh, T.L.; Lippl, K.; Serrer, K.; et al. The Clinically Used Iron Chelator Deferasirox Is an Inhibitor of Epigenetic JumonjiC Domain-Containing Histone Demethylases. ACS Chem. Biol. 2019, 14, 1737–1750. [Google Scholar] [CrossRef]
- Luo, X.; Liu, Y.; Kubicek, S.; Myllyharju, J.; Tumber, A.; Ng, S.; Che, K.H.; Podoll, J.; Heightman, T.D.; Oppermann, U.; et al. A selective inhibitor and probe of the cellular functions of Jumonji C domain-containing histone demethylases. J. Am. Chem. Soc. 2011, 133, 9451–9456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajer, J.M.; Furdas, S.D.; Grunder, A.; Gothwal, M.; Heinicke, U.; Keller, K.; Colland, F.; Fulda, S.; Pahl, H.L.; Fichtner, I.; et al. Histone acetyltransferase inhibitors block neuroblastoma cell growth in vivo. Oncogenesis 2015, 4, e137. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, V.C.; de Abreu da Silva, I.C.; Torres, E.J.; Caby, S.; Lancelot, J.; Vanderstraete, M.; Furdas, S.D.; Jung, M.; Pierce, R.J.; Fantappie, M.R. Epigenetic changes modulate schistosome egg formation and are a novel target for reducing transmission of schistosomiasis. PLoS Pathog. 2014, 10, e1004116. [Google Scholar] [CrossRef] [PubMed]
- Willmann, D.; Lim, S.; Wetzel, S.; Metzger, E.; Jandausch, A.; Wilk, W.; Jung, M.; Forne, I.; Imhof, A.; Janzer, A.; et al. Impairment of prostate cancer cell growth by a selective and reversible lysine-specific demethylase 1 inhibitor. Int. J. Cancer 2012, 131, 2704–2709. [Google Scholar] [CrossRef]
- Schmitt, M.L.; Hauser, A.T.; Carlino, L.; Pippel, M.; Schulz-Fincke, J.; Metzger, E.; Willmann, D.; Yiu, T.; Barton, M.; Schule, R.; et al. Nonpeptidic propargylamines as inhibitors of lysine specific demethylase 1 (LSD1) with cellular activity. J. Med. Chem. 2013, 56, 7334–7342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimburg, T.; Chakrabarti, A.; Lancelot, J.; Marek, M.; Melesina, J.; Hauser, A.T.; Shaik, T.B.; Duclaud, S.; Robaa, D.; Erdmann, F.; et al. Structure-Based Design and Synthesis of Novel Inhibitors Targeting HDAC8 from Schistosoma mansoni for the Treatment of Schistosomiasis. J. Med. Chem. 2016, 59, 2423–2435. [Google Scholar] [CrossRef] [Green Version]
- Tatamiya, T.; Saito, A.; Sugawara, T.; Nakanishi, O. Isozyme-Selective Activity of the HDAC inhibitor MS-275; AACR: Phila-delphia, PA, USA, 2004. [Google Scholar]
- Sharma, N.; Sankrityayan, H.; Kale, A.; Gaikwad, A.B. Role of SET7/9 in the progression of ischemic renal injury in diabetic and non-diabetic rats. Biochem. Biophys. Res. Commun. 2020, 528, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Fujiwara, T.; Niwa, H.; Hirano, M.; Ohira, K.; Okazaki, Y.; Sato, S.; Umehara, T.; Maemoto, Y.; Ito, A.; et al. Development of Novel Inhibitors for Histone Methyltransferase SET7/9 based on Cyproheptadine. ChemMedChem 2018, 13, 1530–1540. [Google Scholar] [CrossRef]
- Keijzer-Veen, M.G.; Schrevel, M.; Finken, M.J.; Dekker, F.W.; Nauta, J.; Hille, E.T.; Frolich, M.; van der Heijden, B.J.; Dutch, P.-C.S.G. Microalbuminuria and lower glomerular filtration rate at young adult age in subjects born very premature and after intrauterine growth retardation. J. Am. Soc. Nephrol. 2005, 16, 2762–2768. [Google Scholar] [CrossRef] [Green Version]
- Hurtado Del Pozo, C.; Garreta, E.; Izpisua Belmonte, J.C.; Montserrat, N. Modeling epigenetic modifications in renal development and disease with organoids and genome editing. Dis. Model Mech. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Nurtjahja-Tjendraputra, E.; Fu, D.; Phang, J.M.; Richardson, D.R. Iron chelation regulates cyclin D1 expression via the proteasome: A link to iron deficiency-mediated growth suppression. Blood 2007, 109, 4045–4054. [Google Scholar] [CrossRef] [PubMed]
- Swali, A.; McMullen, S.; Hayes, H.; Gambling, L.; McArdle, H.J.; Langley-Evans, S.C. Processes underlying the nutritional programming of embryonic development by iron deficiency in the rat. PLoS ONE 2012, 7, e48133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandala, A.; Armstrong, A.; Girresch, B.; Zhu, J.; Chilakala, A.; Chavalmane, S.; Chaudhary, K.; Biswas, P.; Ogilvie, J.; Gnana-Prakasam, J.P. Fenofibrate prevents iron induced activation of canonical Wnt/beta-catenin and oxidative stress signaling in the retina. NPJ Aging Mech. Dis. 2020, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Christova, T.; Perusini, S.; Alizadeh, S.; Bao, R.Y.; Miller, B.W.; Hurren, R.; Jitkova, Y.; Gronda, M.; Isaac, M.; et al. Wnt inhibitor screen reveals iron dependence of beta-catenin signaling in cancers. Cancer Res. 2011, 71, 7628–7639. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.-P.; Liao, M.-C.; Chang, S.-Y.; Abdo, S.; Aliou, Y.; Chenier, I.; Ingelfinger, J.R.; Zhang, S.-L. Maternal diabetes modulates kidney formation in murine progeny: The role of hedgehog interacting protein (HHIP). Diabetologia 2014, 57, 1986–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.-Y.; Chen, Y.-W.; Zhao, X.-P.; Chenier, I.; Tran, S.; Sauvé, A.; Ingelfinger, J.R.; Zhang, S.-L. Catalase prevents maternal diabetes–induced perinatal programming via the Nrf2–HO-1 defense system. Diabetes 2012, 61, 2565–2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-W.; Chenier, I.; Tran, S.; Scotcher, M.; Chang, S.-Y.; Zhang, S.-L. Maternal diabetes programs hypertension and kidney injury in offspring. Pediatric Nephrol. 2010, 25, 1319–1329. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Chenier, I.; Chang, S.-Y.; Tran, S.; Ingelfinger, J.R.; Zhang, S.-L. High glucose promotes nascent nephron apoptosis via NF-κB and p53 pathways. Am. J. Physiol. Ren. Physiol. 2010, 300, F147–F156. [Google Scholar] [CrossRef]
- Nehiri, T.; Van Huyen, J.-P.D.; Viltard, M.; Fassot, C.; Heudes, D.; Freund, N.; Deschênes, G.; Houillier, P.; Bruneval, P.; Lelièvre-Pégorier, M. Exposure to maternal diabetes induces salt-sensitive hypertension and impairs renal function in adult rat offspring. Diabetes 2008, 57, 2167–2175. [Google Scholar] [CrossRef] [Green Version]
- França-Silva, N.; Oliveira, N.D.G.; Balbi, A.P.C. Morphofunctional renal alterations in rats induced by intrauterine hyperglycemic environment. Arch. Med. Sci. AMS 2016, 12, 243. [Google Scholar] [CrossRef] [Green Version]
- Pani, L.; Horal, M.; Loeken, M.R. Polymorphic susceptibility to the molecular causes of neural tube defects during diabetic embryopathy. Diabetes 2002, 51, 2871–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, T.A.; Denno, K.M.; Sipos, G.F.; Sadler, T.W. Diabetic teratogenesis. In vitro evidence for a multifactorial etiology with little contribution from glucose per se. Diabetes 1994, 43, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Wentzel, P.; Eriksson, U.J. Insulin treatment fails to abolish the teratogenic potential of serum from diabetic rats. Eur. J. Endocrinol. 1996, 134, 459–466. [Google Scholar] [CrossRef]
- El Hajj, N.; Pliushch, G.; Schneider, E.; Dittrich, M.; Muller, T.; Korenkov, M.; Aretz, M.; Zechner, U.; Lehnen, H.; Haaf, T. Metabolic programming of MEST DNA methylation by intrauterine exposure to gestational diabetes mellitus. Diabetes 2013, 62, 1320–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anguita-Ruiz, A.; Mendez-Gutierrez, A.; Ruperez, A.I.; Leis, R.; Bueno, G.; Gil-Campos, M.; Tofe, I.; Gomez-Llorente, C.; Moreno, L.A.; Gil, A.; et al. The protein S100A4 as a novel marker of insulin resistance in prepubertal and pubertal children with obesity. Metabolism 2020, 105, 154187. [Google Scholar] [CrossRef] [PubMed]
- Hjort, L.; Martino, D.; Grunnet, L.G.; Naeem, H.; Maksimovic, J.; Olsson, A.H.; Zhang, C.; Ling, C.; Olsen, S.F.; Saffery, R.; et al. Gestational diabetes and maternal obesity are associated with epigenome-wide methylation changes in children. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Quilter, C.R.; Cooper, W.N.; Cliffe, K.M.; Skinner, B.M.; Prentice, P.M.; Nelson, L.; Bauer, J.; Ong, K.K.; Constancia, M.; Lowe, W.L.; et al. Impact on offspring methylation patterns of maternal gestational diabetes mellitus and intrauterine growth restraint suggest common genes and pathways linked to subsequent type 2 diabetes risk. FASEB J. 2014, 28, 4868–4879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilliard, S.A.; El-Dahr, S.S. Epigenetics mechanisms in renal development. Pediatric Nephrol. 2016, 31, 1055–1060. [Google Scholar] [CrossRef]
- Wanner, N.; Bechtel-Walz, W. Epigenetics of kidney disease. Cell Tissue Res. 2017, 369, 75–92. [Google Scholar] [CrossRef]
- Bechtel, W.; McGoohan, S.; Zeisberg, E.M.; Muller, G.A.; Kalbacher, H.; Salant, D.J.; Muller, C.A.; Kalluri, R.; Zeisberg, M. Methylation determines fibroblast activation and fibrogenesis in the kidney. Nat. Med. 2010, 16, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Yao, X.; Li, Y.; Saifudeen, Z.; Bachvarov, D.; El-Dahr, S.S. Histone deacetylase 1 and 2 regulate Wnt and p53 pathways in the ureteric bud epithelium. Development 2015, 142, 1180–1192. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, S.L.; Chen, S.; McLaughlin, N.; El-Dahr, S.S. Regulation of kidney development by histone deacetylases. Pediatric Nephrol. 2011, 26, 1445–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, A.; Valerius, M.T.; Mugford, J.W.; Carroll, T.J.; Self, M.; Oliver, G.; McMahon, A.P. Six2 defines and regulates a multipotent self-renewing nephron progenitor population throughout mammalian kidney development. Cell Stem Cell 2008, 3, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Kumaki, Y.; Oda, M.; Okano, M. QUMA: Quantification tool for methylation analysis. Nucleic Acids Res. 2008, 36, W170–W175. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Cohen, C.D.; Frach, K.; Schlondorff, D.; Kretzler, M. Quantitative gene expression analysis in renal biopsies: A novel protocol for a high-throughput multicenter application. Kidney Int. 2002, 61, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, C.D.; Klingenhoff, A.; Boucherot, A.; Nitsche, A.; Henger, A.; Brunner, B.; Schmid, H.; Merkle, M.; Saleem, M.A.; Koller, K.P.; et al. Comparative promoter analysis allows de novo identification of specialized cell junction-associated proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 5682–5687. [Google Scholar] [CrossRef] [Green Version]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [Green Version]
- Mamo, S.; Gal, A.B.; Bodo, S.; Dinnyes, A. Quantitative evaluation and selection of reference genes in mouse oocytes and embryos cultured in vivo and in vitro. BMC Dev Biol. 2007, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuhrmann, L.; Lindner, S.; Hauser, A.-T.; Höse, C.; Kretz, O.; Cohen, C.D.; Lindenmeyer, M.T.; Sippl, W.; Jung, M.; Huber, T.B.; et al. Effects of Environmental Conditions on Nephron Number: Modeling Maternal Disease and Epigenetic Regulation in Renal Development. Int. J. Mol. Sci. 2021, 22, 4157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084157
Fuhrmann L, Lindner S, Hauser A-T, Höse C, Kretz O, Cohen CD, Lindenmeyer MT, Sippl W, Jung M, Huber TB, et al. Effects of Environmental Conditions on Nephron Number: Modeling Maternal Disease and Epigenetic Regulation in Renal Development. International Journal of Molecular Sciences. 2021; 22(8):4157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084157
Chicago/Turabian StyleFuhrmann, Lars, Saskia Lindner, Alexander-Thomas Hauser, Clemens Höse, Oliver Kretz, Clemens D. Cohen, Maja T. Lindenmeyer, Wolfgang Sippl, Manfred Jung, Tobias B. Huber, and et al. 2021. "Effects of Environmental Conditions on Nephron Number: Modeling Maternal Disease and Epigenetic Regulation in Renal Development" International Journal of Molecular Sciences 22, no. 8: 4157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084157