
Carbon Source-Dependent Changes of the Structure of Streptococcus pneumoniae Capsular Polysaccharide with Serotype 6F

 , , ,
, , ,  and
and
Abstract
:1. Introduction
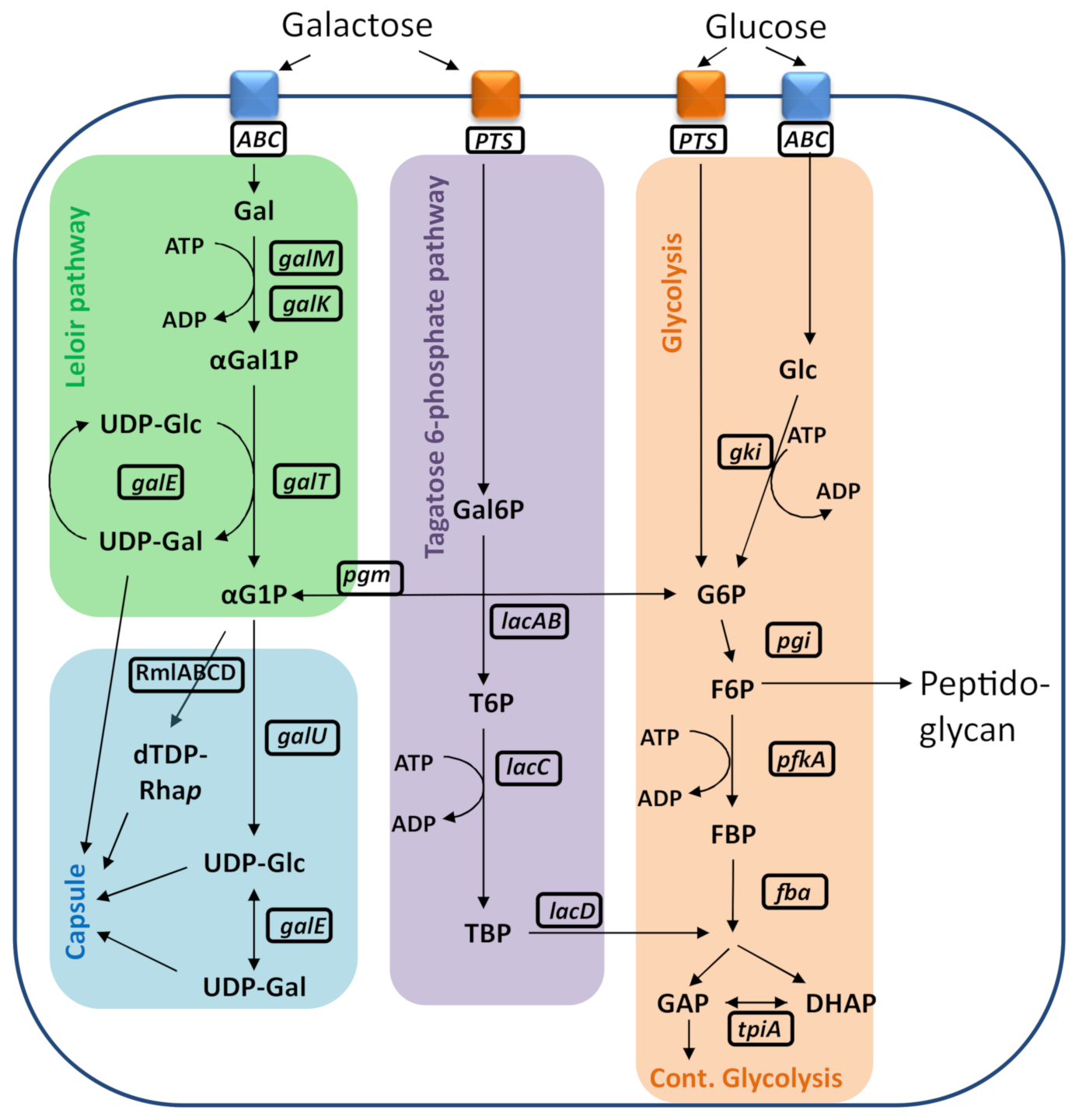
2. Results
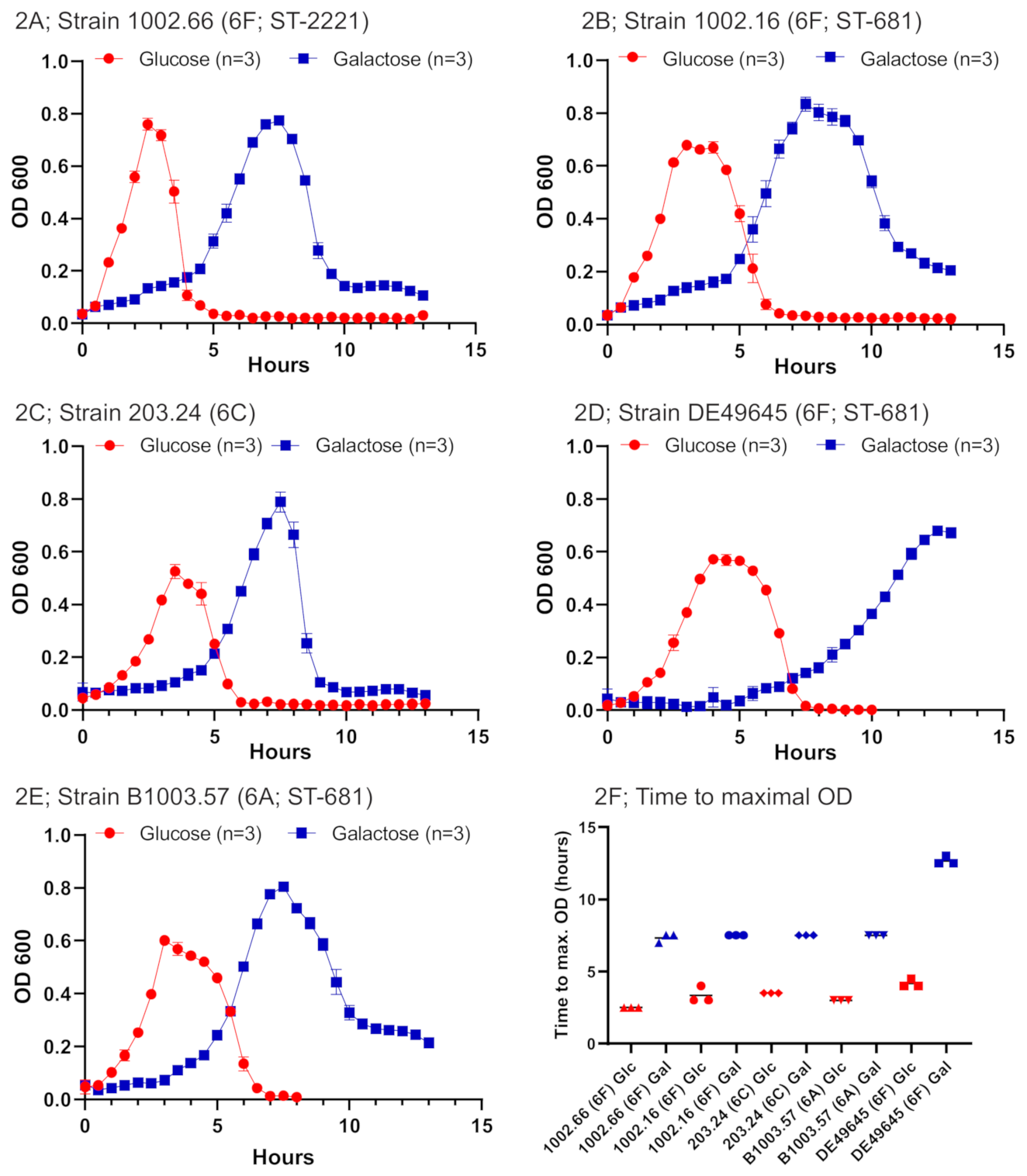
2.1. Bacterial Growth under Different Conditions for Serogroup 6 Strains
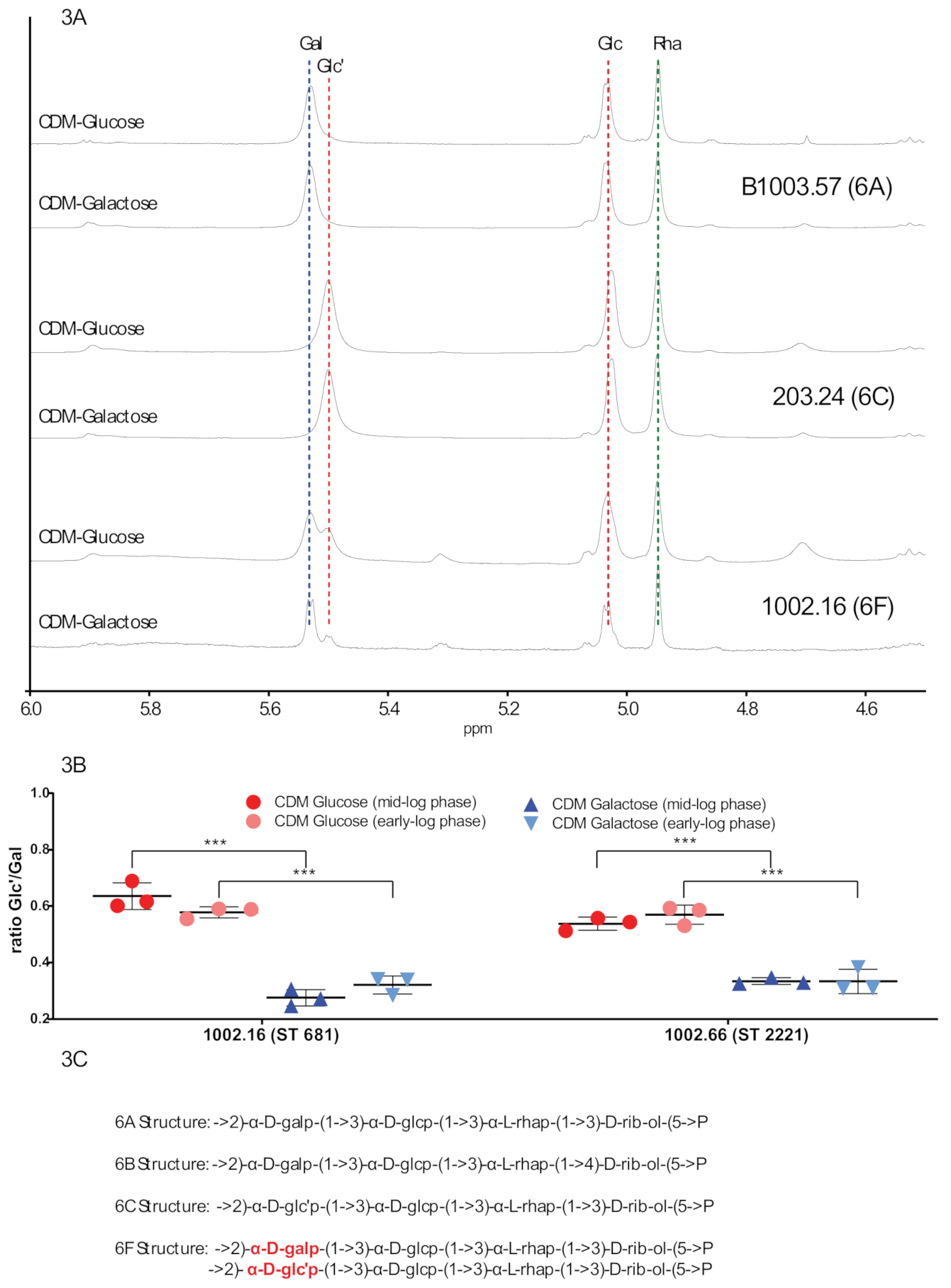
2.2. NMR Capsule Structure Analysis Indicates Differences within 6F Strains According to the Used Carbon Sources
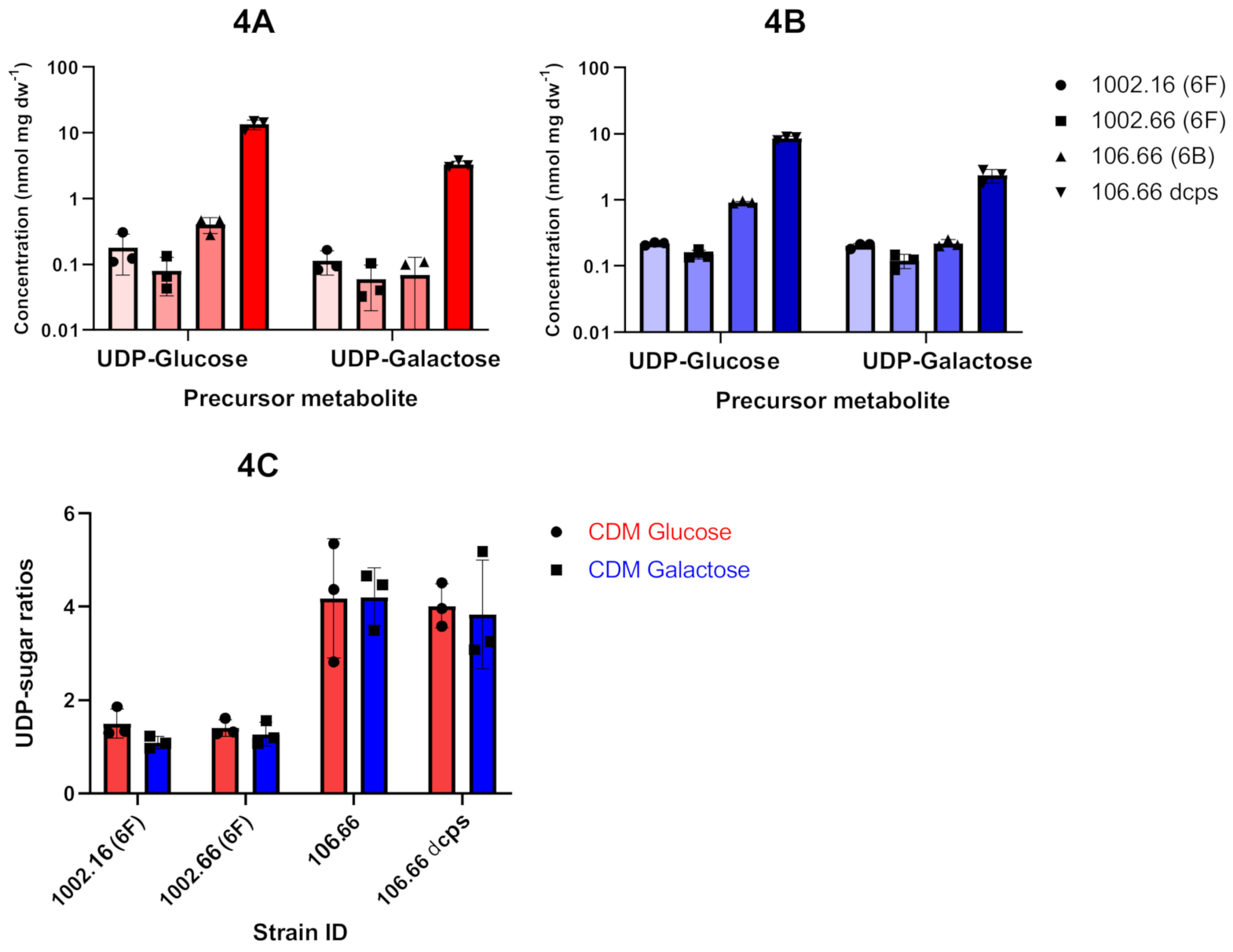
2.3. Analysis of Polysaccharide Capsule Metabolites by 31P NMR
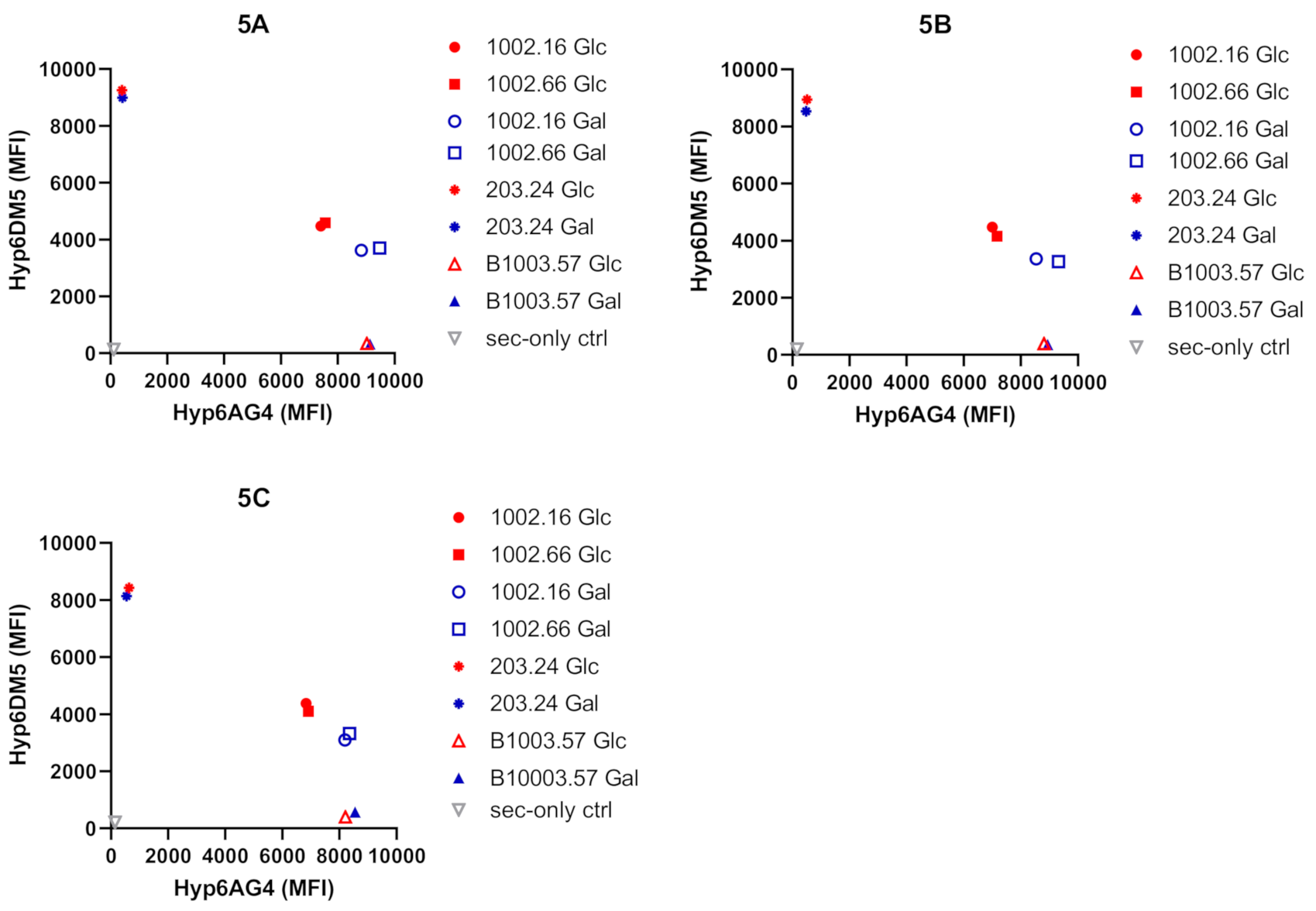
2.4. Flow Cytometry Analyses Show Different Bindings of Monoclonal Antibodies (mAbs)
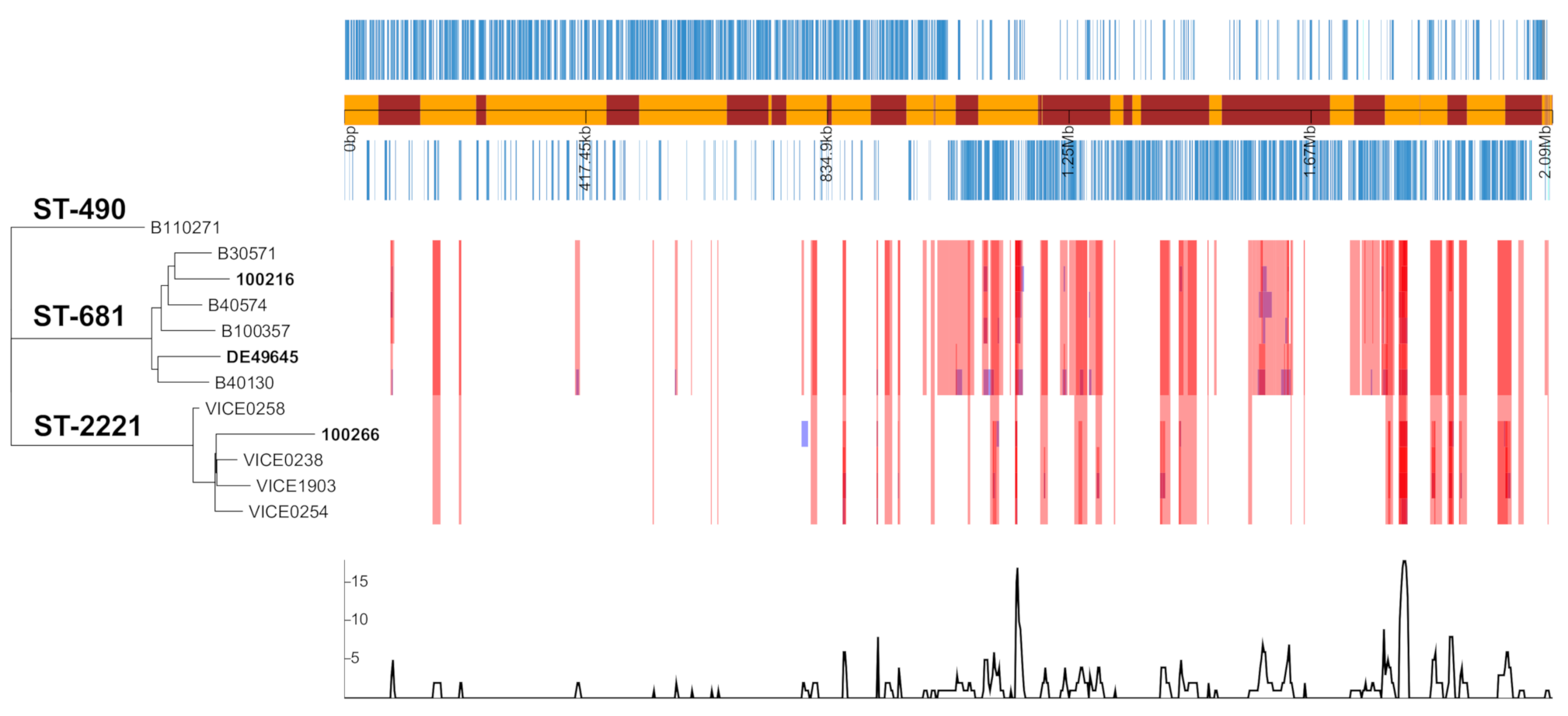
2.5. Analysis of Genome Assemblies of Serogroup 6 Strains
2.6. Evolutionary Events Leading to Serotype 6F Strains
3. Discussion
4. Materials and Methods
4.1. Selection of Bacterial Isolates
4.2. Bacterial Growth Conditions
4.3. Capsular Polysaccharide Extraction
4.4. H NMR Measurements of the Polysaccharide Capsule
4.5. Intracellular Metabolite Extraction, Identification and Quantification of Capsule Precursor Signals by 31P NMR
4.6. Fluorescence-Activated Cell Sorting (FACS) Experiments
4.7. DNA Library Preparation and Whole-Genome Sequencing (WGS)
4.8. Genomic Analysis and Assembly
4.9. Phylogenetic and Recombination Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Austrian, R. The Pneumococcus at the Millennium: Not Down, Not Out. J. Infect. Dis. 1999, 179 (Suppl. 2), S338–S341. [Google Scholar] [CrossRef] [PubMed]
- Tuomanen, E.I.; Austrian, R.; Masure, H.R. Pathogenesis of Pneumococcal Infection. N. Engl. J. Med. 1995, 332, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Geno, K.A.; Gilbert, G.L.; Song, J.Y.; Skovsted, I.C.; Klugman, K.P.; Jones, C.; Konradsen, H.B.; Nahm, M.H. Pneumococcal Capsules and Their Types: Past, Present, and Future. Clin. Microbiol. Rev. 2015, 28, 871–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meichtry, J.; Born, R.; Küffer, M.; Zwahlen, M.; Albrich, W.C.; Brugger, S.D.; Mühlemann, K.; Hilty, M. Serotype epidemiology of invasive pneumococcal disease in Swiss adults: A nationwide population-based study. Vaccine 2014, 32, 5185–5191. [Google Scholar] [CrossRef] [Green Version]
- Allemann, A.; Frey, P.M.; Brugger, S.D.; Hilty, M. Pneumococcal carriage and serotype variation before and after introduction of pneumococcal conjugate vaccines in patients with acute otitis media in Switzerland. Vaccine 2017, 35, 1946–1953. [Google Scholar] [CrossRef]
- Hauser, C.; Kronenberg, A.; Allemann, A.; Mühlemann, K.; Hilty, M. Serotype/serogroup-specific antibiotic non-susceptibility of invasive and non-invasive Streptococcus pneumoniae, Switzerland, 2004 to 2014. Eurosurveillance 2016, 21, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Càmara, J.; Grau, I.; González-Díaz, A.; Tubau, F.; Calatayud, L.; Cubero, M.; Domínguez, M.Á.; Liñares, J.; Yuste, J.; Pallarés, R.; et al. A historical perspective of MDR invasive pneumococcal disease in Spanish adults. J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef]
- Lee, C.; Choi, S.K.; Kim, R.K.; Kim, H.; Whang, Y.H.; Pham, T.-H.; Cheon, H.; Yoon, D.-Y.; Kim, C.W.; Baik, Y.O.; et al. Development of a new 15-valent pneumococcal conjugate vaccine (PCV15) and evaluation of its immunogenicity. Biologicals 2019, 61, 32–37. [Google Scholar] [CrossRef]
- Hobbs, J.K.; Pluvinage, B.; Boraston, A.B. Glycan-metabolizing enzymes in microbe-host interactions: The Streptococcus pneumoniae paradigm. FEBS Lett. 2018, 592, 3865–3897. [Google Scholar] [CrossRef]
- Aprianto, R.; Slager, J.; Holsappel, S.; Veening, J.-W. High-resolution analysis of the pneumococcal transcriptome under a wide range of infection-relevant conditions. Nucleic Acids Res. 2018, 46, 9990–10006. [Google Scholar] [CrossRef]
- Bidossi, A.; Mulas, L.; Decorosi, F.; Colomba, L.; Ricci, S.; Pozzi, G.; Deutscher, J.; Viti, C.; Oggioni, M.R. A Functional Genomics Approach to Establish the Complement of Carbohydrate Transporters in Streptococcus pneumoniae. PLoS ONE 2012, 7, e33320. [Google Scholar] [CrossRef] [Green Version]
- Brugger, S.D.; Troxler, L.J.; Rüfenacht, S.; Frey, P.M.; Morand, B.; Geyer, R.; Mühlemann, K.; Höck, S.; Thormann, W.; Furrer, J.; et al. Polysaccharide Capsule Composition of Pneumococcal Serotype 19A Subtypes Is Unaltered among Subtypes and Independent of the Nutritional Environment. Infect. Immun. 2016, 84, 3152–3160. [Google Scholar] [CrossRef] [Green Version]
- Elberse, K.; Witteveen, S.; Van Der Heide, H.; Van De Pol, I.; Schot, C.; Van Der Ende, A.; Berbers, G.; Schouls, L. Sequence Diversity within the Capsular Genes of Streptococcus pneumoniae Serogroup 6 and 19. PLoS ONE 2011, 6, e25018. [Google Scholar] [CrossRef] [Green Version]
- Van Tonder, A.J.; Bray, J.E.; Quirk, S.J.; Haraldsson, G.; Jolley, K.A.; Maiden, M.C.J.; Hoffmann, S.; Bentley, S.D.; Haraldsson, A.; Erlendsdottir, H.; et al. Putatively novel serotypes and the potential for reduced vaccine effectiveness: Capsular locus diversity revealed among 5405 pneumococcal genomes. Microb. Genom. 2016, 2, 000090. [Google Scholar] [CrossRef] [Green Version]
- Kapatai, G.; Sheppard, C.L.; Troxler, L.J.; Litt, D.J.; Furrer, J.; Hilty, M.; Fry, N.K. Pneumococcal 23B Molecular Subtype Identified Using Whole Genome Sequencing. Genome Biol. Evol. 2017, 9, 2145–2158. [Google Scholar] [CrossRef]
- Oliver, M.B.; Jones, C.; Larson, T.R.; Calix, J.J.; Zartler, E.R.; Yother, J.; Nahm, M.H. Streptococcus pneumoniae Serotype 11D Has a Bispecific Glycosyltransferase and Expresses Two Different Capsular Polysaccharide Repeating Units. J. Biol. Chem. 2013, 288, 21945–21954. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.B.; van der Linden, M.P.G.; Küntzel, S.A.; Saad, J.S.; Nahm, M.H. Discovery of Streptococcus pneumoniae Serotype 6 Variants with Glycosyltransferases Synthesizing Two Differing Repeating Units. J. Biol. Chem. 2013, 288, 25976–25985. [Google Scholar] [CrossRef] [Green Version]
- Paixao, L.; Oliveira, J.; Verissimo, A.; Vinga, S.; Lourenco, E.C.; Ventura, M.R.; Kjos, M.; Veening, J.W.; Fernandes, V.E.; Andrew, P.W.; et al. Host glycan sugar-specific pathways in Streptococcus pneumoniae: Galactose as a key sugar in colonisation and infection. PLoS ONE 2015, 10, e0121042. [Google Scholar] [CrossRef] [Green Version]
- Paixão, L.; Caldas, J.; Kloosterman, T.G.; Kuipers, O.P.; Vinga, S.; Neves, A.R. Transcriptional and metabolic effects of glucose on Streptococcus pneumoniae sugar metabolism. Front. Microbiol. 2015, 6, 1041. [Google Scholar] [CrossRef]
- Quirk, S.J.; Haraldsson, G.; Hjálmarsdóttir, M.Á.; Van Tonder, A.J.; Hrafnkelsson, B.; Bentley, S.D.; Haraldsson, Á.; Erlendsdóttir, H.; Brueggemann, A.B.; Kristinsson, K.G. Vaccination of Icelandic Children with the 10-Valent Pneumococcal Vaccine Leads to a Significant Herd Effect among Adults in Iceland. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Troxler, L.J.; Werren, J.P.; Schaffner, T.O.; Mostacci, N.; Vermathen, P.; Vermathen, M.; Wüthrich, D.; Simillion, C.; Brugger, S.D.; Bruggmann, R.; et al. Carbon source regulates polysaccharide capsule biosynthesis in Streptococcus pneumoniae. J. Biol. Chem. 2019, 294, 17224–17238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiller, N.L.; Sá-Leão, R. Puzzling Over the Pneumococcal Pangenome. Front. Microbiol. 2018, 9, 2580. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Aanensen, D.M.; Mavroidi, A.; Saunders, D.; Rabbinowitsch, E.; Collins, M.; Donohoe, K.; Harris, D.; Murphy, L.; Quail, M.A.; et al. Genetic Analysis of the Capsular Biosynthetic Locus from All 90 Pneumococcal Serotypes. PLoS Genet. 2006, 2, e31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slotved, H.-C.; Satzke, C. In vitro growth of pneumococcal isolates representing 23 different serotypes. BMC Res. Notes 2013, 6, 208. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.M.; Kuipers, O.P.; Neves, A.R. Environmental and Nutritional Factors That Affect Growth and Metabolism of the Pneumococcal Serotype 2 Strain D39 and Its Nonencapsulated Derivative Strain R6. PLoS ONE 2013, 8, e58492. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.M.; Kloosterman, T.G.; Kuipers, O.P.; Neves, A.R. CcpA Ensures Optimal Metabolic Fitness of Streptococcus pneumoniae. PLoS ONE 2011, 6, e26707. [Google Scholar] [CrossRef] [Green Version]
- Park, I.H.; Park, S.; Hollingshead, S.K.; Nahm, M.H. Genetic Basis for the New Pneumococcal Serotype, 6C. Infect. Immun. 2007, 75, 4482–4489. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Adams, M.H.; Roe, A.S. A Partially Defined Medium for Cultivation of Pneumococcus. J. Bacteriol. 1945, 49, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Lacks, S. Integration efficiency and genetic recombination in pneumococcal transformation. Genetics 1966, 53, 207–235. [Google Scholar] [CrossRef]
- Lacks, S.; Hotchkiss, R.D. A study of the genetic material determining an enzyme activity in Pneumococcus. Biochim. Biophys. Acta Bioenerg. 1960, 39, 508–518. [Google Scholar] [CrossRef]
- Hathaway, L.J.; Brugger, S.D.; Morand, B.; Bangert, M.; Rotzetter, J.U.; Hauser, C.; Graber, W.A.; Gore, S.; Kadioglu, A.; Mühlemann, K. Capsule Type of Streptococcus pneumoniae Determines Growth Phenotype. PLoS Pathog. 2012, 8, e1002574. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Snippy: Rapid haploid variant calling and core genome alignment. 2014. [Google Scholar]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; AbuDahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MLST | Serotype | Year of Isolation | Isolate | Country | Genome Size (bp) | Reference Accession Number | |
|---|---|---|---|---|---|---|---|
| B100357 | 681 | 6A | 2011 | Blood | Switzerland | 2,159,962 ** | This study; SRR11550454 |
| B110271 | 490 | 6A | 2011 | Blood | Switzerland | 2,091,771 ** | This study; SRR11550453 |
| B30571 | 681 | 6A | 2004 | Blood | Switzerland | 2,141,354 ** | This study; SRR11550452 |
| B40130 | 681 | 6A | 2005 | Blood | Switzerland | 2,116,757 ** | This study; SRR11550451 |
| B40574 | 681 | 6A | 2005 | Blood | Switzerland | 2,167,664 ** | This study; SRR11550459 |
| VICE0238 * | 2221 | 6A | 2011 | Blood | Iceland | 2,132,191 ** | [20]; ERR388909 |
| VICE0258 * | 2221 | 6A | 2012 | Blood | Iceland | 2,130,915 ** | [20]; ERR388920 |
| VICE0254 * | 2221 | 6A | 2012 | Blood | Iceland | 2,132,423 ** | [20]; ERR388919 |
| VICE1903 * | 2221 | 6A | 2014 | Blood | Iceland | 2,137,293 ** | [20]; ENA: ERR755620 |
| 106.66 | 2244 | 6B | NP | Switzerland | 2,130,915 ** | [21]; ENA: ERR388920 | |
| 106.66 Δcps Janus | 2244 | nt | NP | Switzerland | n.a. | [21] | |
| 203.24 | n.a. | 6C | NP | Switzerland | n.a. | [21] | |
| 1002.16 | 681 | 6F | 2010 | Blood | Switzerland | 2,238,890 *** | This study; SRR9943963; SRR9960027 |
| 1002.66 | 2221 | 6F | 2011 | Blood | Switzerland | 2,184,396 *** | This study; SRR9943964; SRR9960028 |
| DE49645 | 681 | 6F | 2012 | Blood | Germany | 2,151,471 *** | [17]; SRR11550458; SRR11550455 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Werren, J.P.; Troxler, L.J.; Oyewole, O.R.-A.; Ramette, A.; Brugger, S.D.; Bruggmann, R.; van der Linden, M.; Nahm, M.H.; Gjuroski, I.; Casanova, C.; et al. Carbon Source-Dependent Changes of the Structure of Streptococcus pneumoniae Capsular Polysaccharide with Serotype 6F. Int. J. Mol. Sci. 2021, 22, 4580. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094580
Werren JP, Troxler LJ, Oyewole OR-A, Ramette A, Brugger SD, Bruggmann R, van der Linden M, Nahm MH, Gjuroski I, Casanova C, et al. Carbon Source-Dependent Changes of the Structure of Streptococcus pneumoniae Capsular Polysaccharide with Serotype 6F. International Journal of Molecular Sciences. 2021; 22(9):4580. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094580
Chicago/Turabian StyleWerren, Joel P., Lukas J. Troxler, Oluwaseun Rume-Abiola Oyewole, Alban Ramette, Silvio D. Brugger, Rémy Bruggmann, Mark van der Linden, Moon H. Nahm, Ilche Gjuroski, Carlo Casanova, and et al. 2021. "Carbon Source-Dependent Changes of the Structure of Streptococcus pneumoniae Capsular Polysaccharide with Serotype 6F" International Journal of Molecular Sciences 22, no. 9: 4580. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094580