
Exosomal Vimentin from Adipocyte Progenitors Protects Fibroblasts against Osmotic Stress and Inhibits Apoptosis to Enhance Wound Healing

 ,
, 
Abstract
:1. Introduction
2. Results
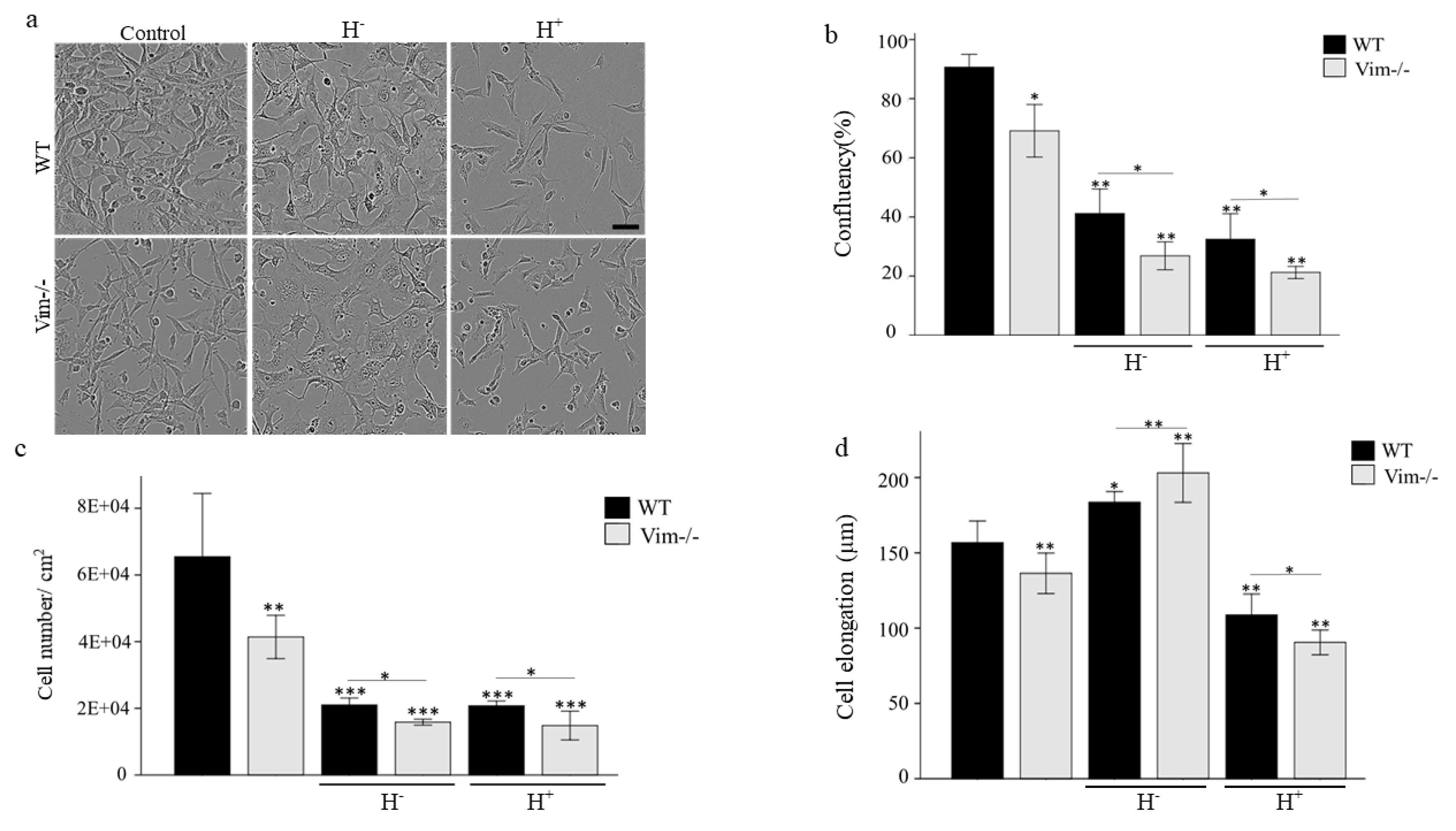
2.1. WT-APCs Tolerate Osmotic Stress Better than Vim−/−APCs
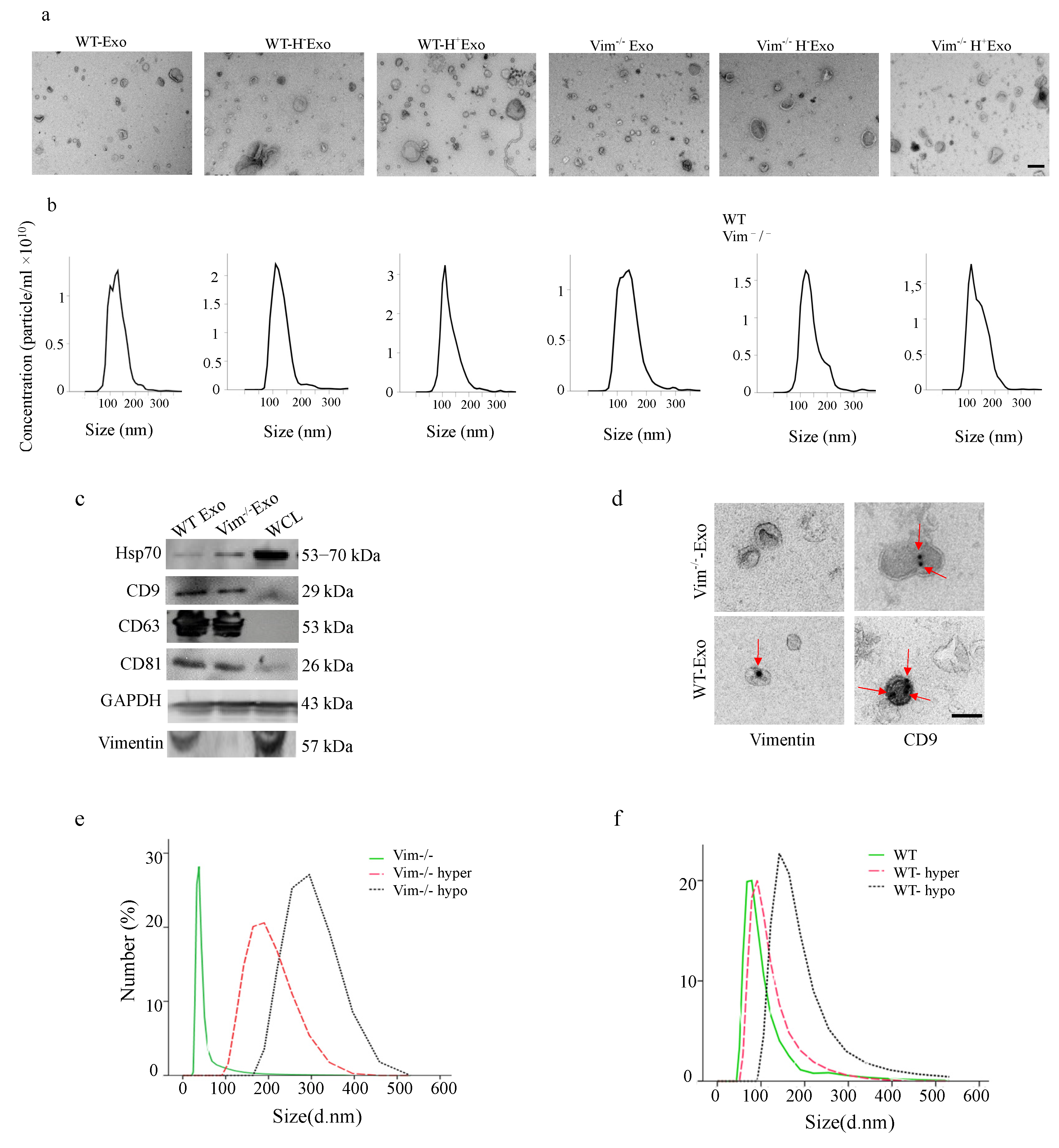
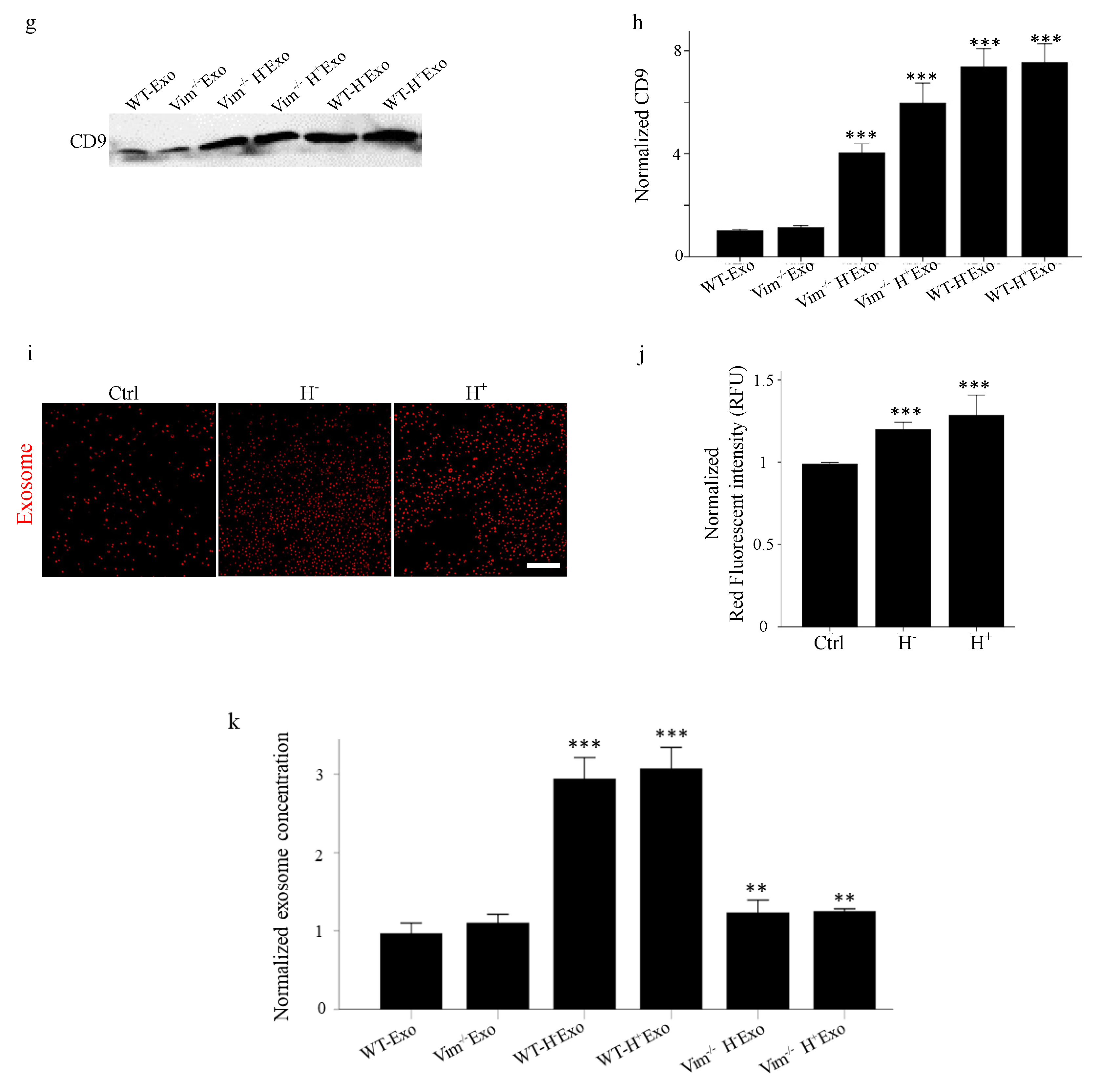
2.2. Osmotic Stress Significantly Increases the Exosome Secretion by WT and Vim−/−APCs
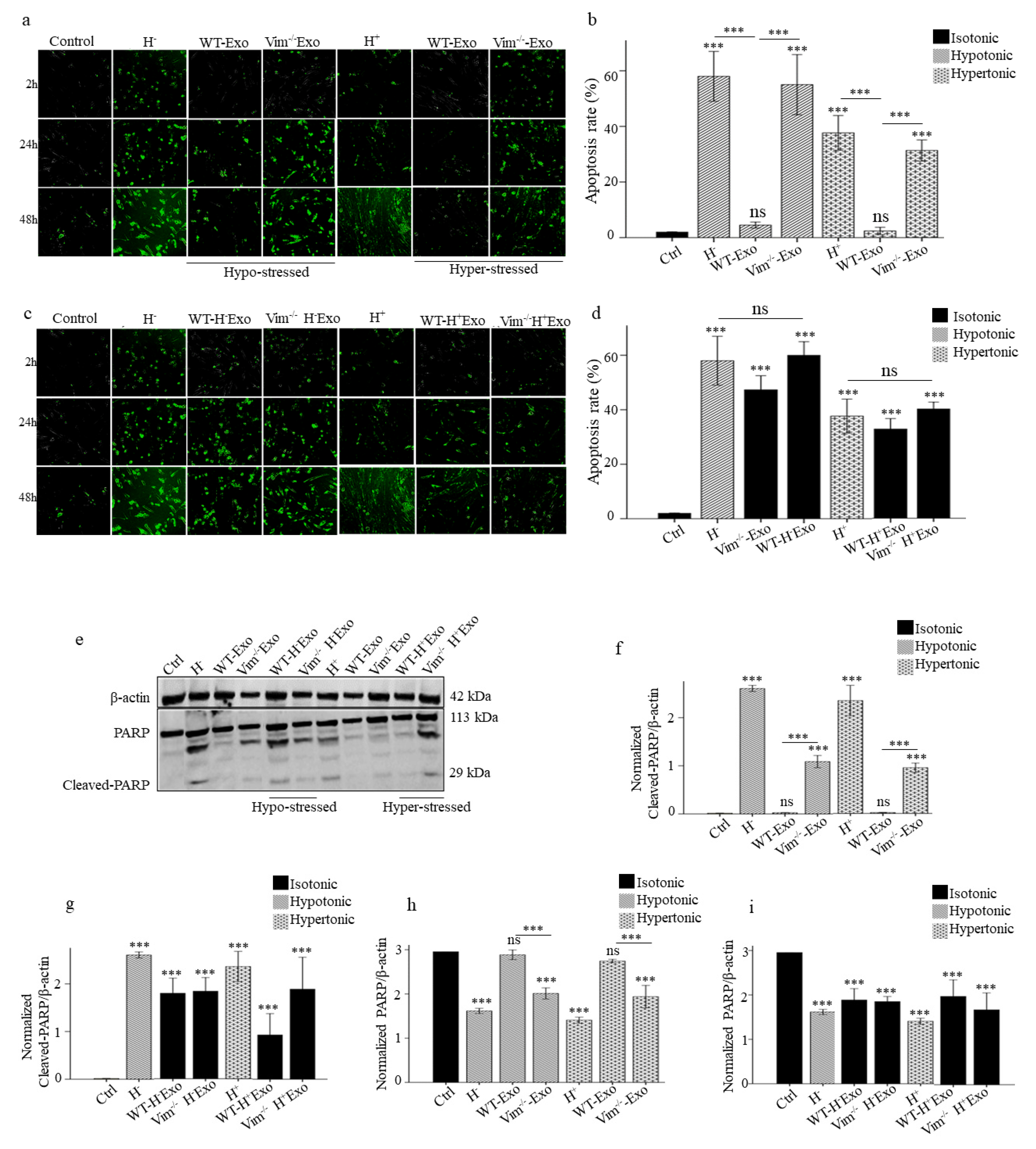
2.3. WT-Exos Promotes Proliferation and Prevents Apoptosis of Osmotic-Stressed HDFs
2.4. Osmotic-Stress Induced Exosomes Influence HDFs Proliferation and Apoptosis
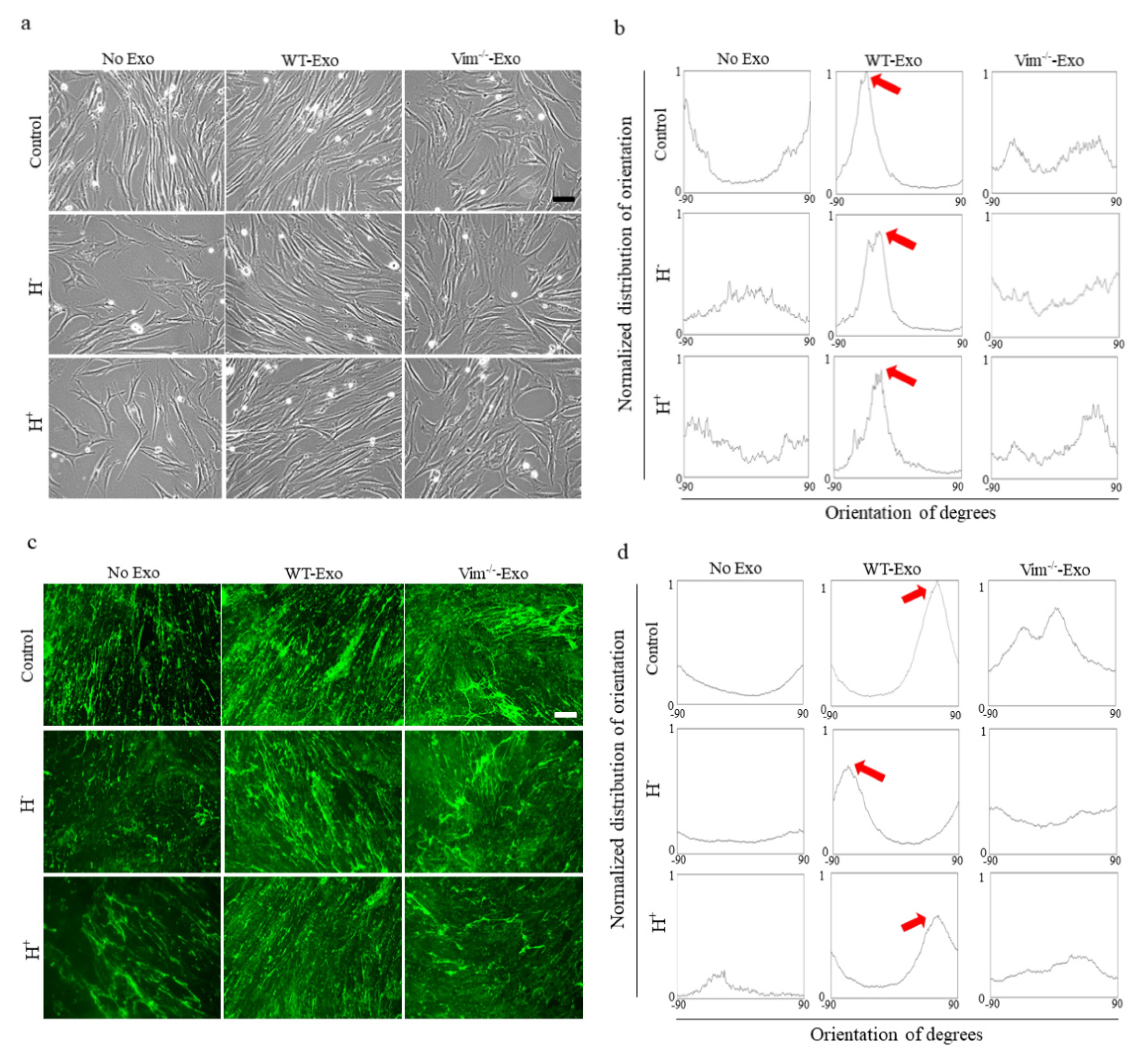
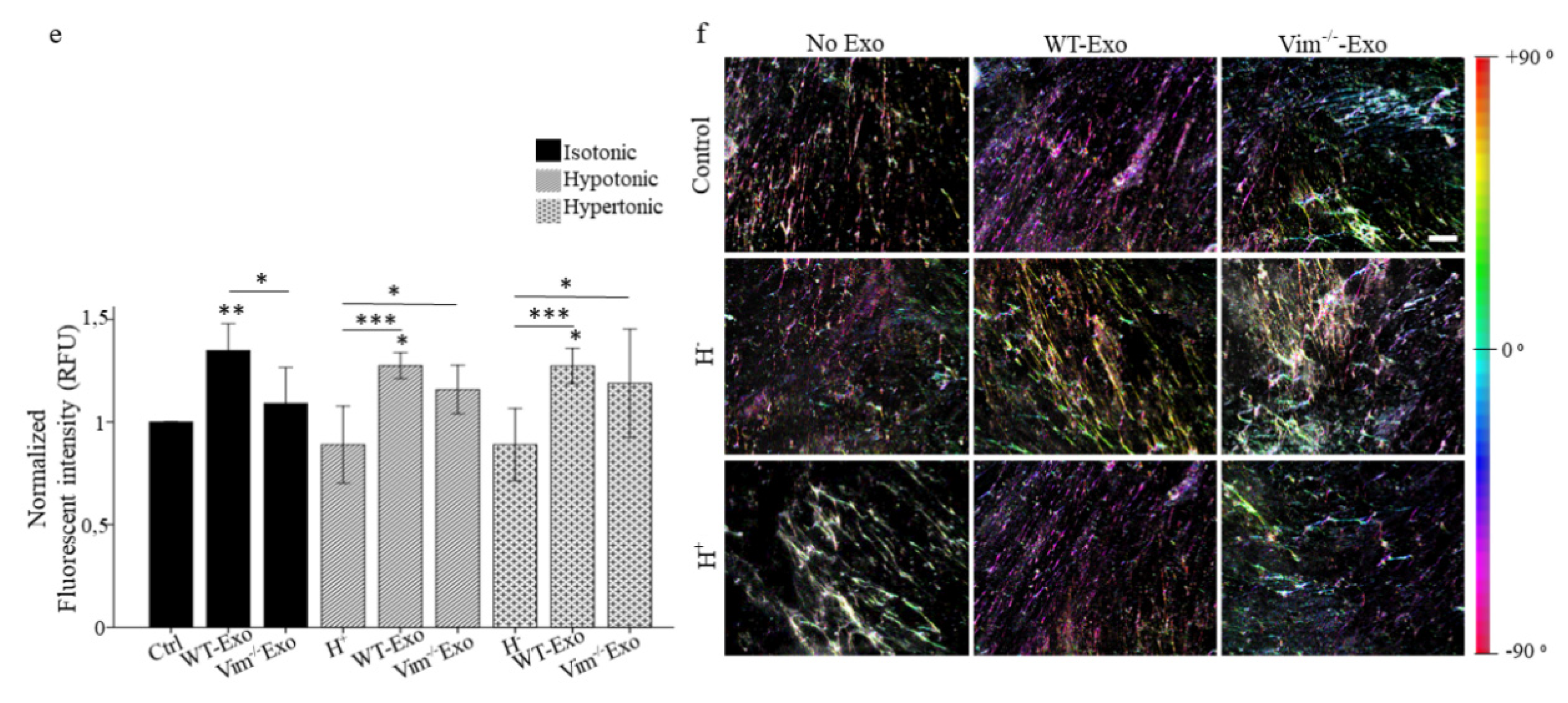
2.5. WT-APC-Exos Affect Collagen Fiber Orientation and Promote ECM Production by Osmotic Stressed HDFs
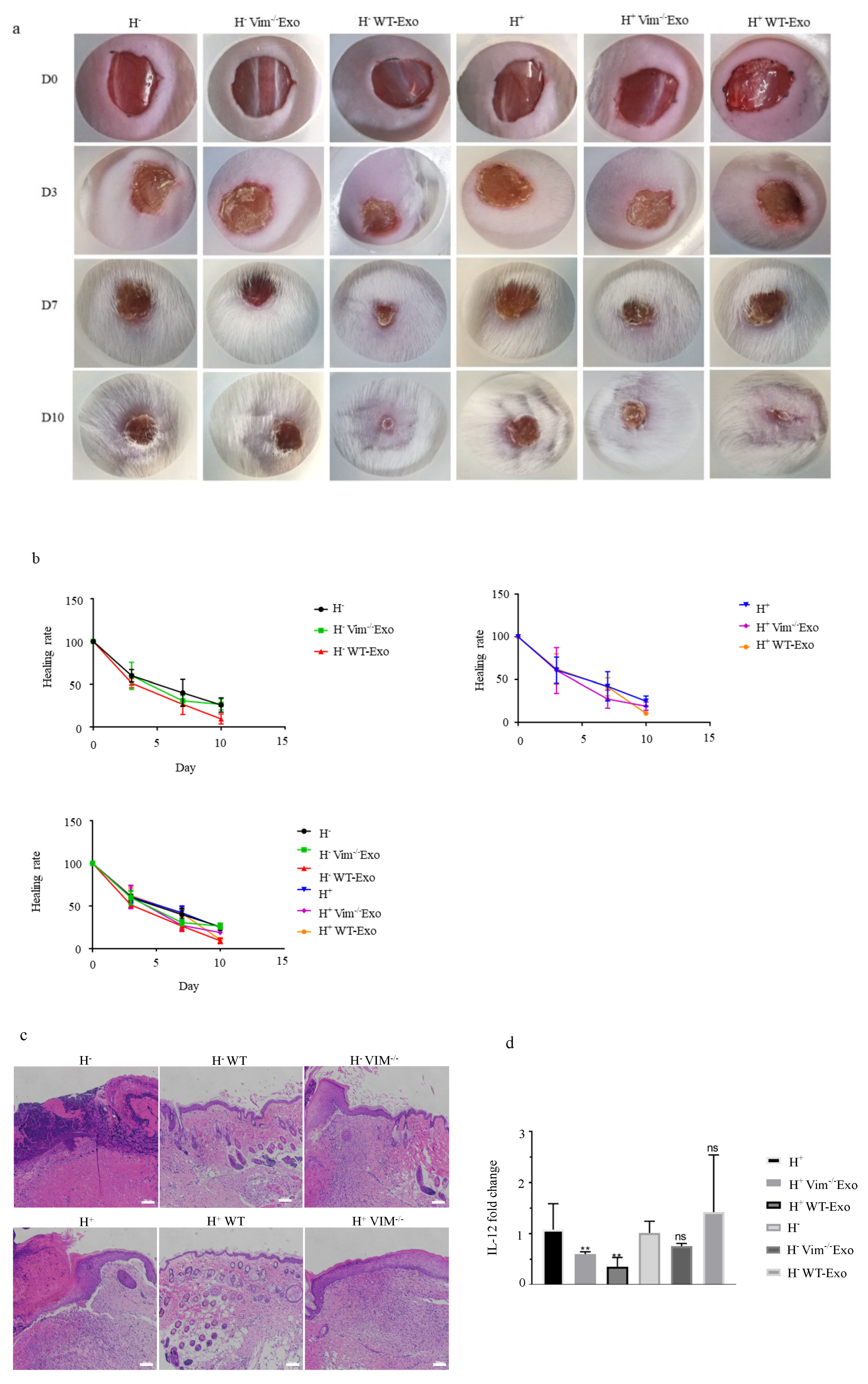
2.6. WT-APCs-Exos Promote Wound Healing In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Exosome Isolation
4.2. Exosome Characterization and Analysis
4.3. Exosome Labeling and Quantification
4.4. HDFs Proliferation Assay
4.5. Exosome Uptake by HDFs
4.6. Cell Apoptosis and Analysis
4.7. ECM Staining and Quantification
4.8. Mouse Skin Injury Model and Treatment
4.9. Histology
4.10. RNA Isolation and qPCR Analysis
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Rosińczuk, J.; Taradaj, J.; Dymarek, R.; Sopel, M. Mechanoregulation of Wound Healing and Skin Homeostasis. BioMed Res. Int. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, L.A.; Marshall, C.D.; Leavitt, T.; Hu, M.S.; Moore, A.L.; Gonzalez, J.G.; Longaker, M.T.; Gurtner, G.C. Mechanical Forces in Cutaneous Wound Healing: Emerging Therapies to Minimize Scar Formation. Adv. Wound Care 2018, 7, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Enyedi, B.; Niethammer, P. Mechanisms of epithelial wound detection. Trends Cell Biol. 2015, 25, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Brocker, C.; Thompson, D.C.; Vasiliou, V. The role of hyperosmotic stress in inflammation and disease. Biomol. Concepts 2012, 3, 345–364. [Google Scholar] [CrossRef]
- Nielsen, M.B.; Christensen, S.T.; Hoffmann, E.K. Effects of osmotic stress on the activity of MAPKs and PDGFR-β- mediated signal transduction in NIH-3T3 fibroblasts. Am. J. Physiol.-Cell Physiol. 2008, 294, 1046–1055. [Google Scholar] [CrossRef]
- Pietuch, A.; Brückner, B.R.; Janshoff, A. Membrane tension homeostasis of epithelial cells through surface area regulation in response to osmotic stress. Biochim. Biophys. Acta-Mol. Cell Res. 2013, 1833, 712–722. [Google Scholar] [CrossRef] [Green Version]
- Kruse, C.R.; Singh, M.; Sørensen, J.A.; Eriksson, E.; Nuutila, K. The effect of local hyperglycemia on skin cells in vitro and on wound healing in euglycemic rats. J. Surg. Res. 2016, 206, 418–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Kartenbeck, J.; Kabsch, K.; Mao, X.; Marqués, M.; Alonso, Á. Stress kinase p38 mediates EGFR transactivation by hyperosmolar concentrations of sorbitol. J. Cell. Physiol. 2002, 192, 234–243. [Google Scholar] [CrossRef]
- Dascalu, A.; Matithyou, A.; Oron, Y.; Korenstein, R. A Hyperosmotic Stimulus Elevates Intracellular Calcium and Inhibits Proliferation of a Human Keratinocyte Cell Line. J. Investig. Dermatol. 2000, 115, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Varela, D.; Held, N.; Linas, S. Overview of Cerebral Edema During Correction of Hyperglycemic Crises. Am. J. Case Rep. 2018, 19, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Jingi, A.M.; Tankeu, A.T.; Ateba, N.A.; Noubiap, J.J. Mechanism of worsening diabetic retinopathy with rapid lowering of blood glucose: The synergistic hypothesis. BMC Endocr. Disord. 2017, 17, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, M.B.; Cherry, G.; Heimburger, S.; Hay, M.; Haydel, H.; Cooley, L. The Effect of Edema and External Pressure on Wound Healing. Arch. Surg. 1967, 94, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Rowan, M.P.; Cancio, L.C.; Elster, E.A.; Burmeister, D.M.; Rose, L.F.; Natesan, S.; Chan, R.K.; Christy, R.J.; Chung, K.K. Burn wound healing and treatment: Review and advancements. Crit. Care 2015, 19, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, A.D.F.; Gomes, D.A. Stem Cell Extracellular Vesicles in Skin Repair. Bioengineering 2018, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, J.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Extracellular vesicles as modulators of wound healing. Adv. Drug Deliv. Rev. 2018, 129, 394–406. [Google Scholar] [CrossRef]
- Hedlund, M.; Nagaeva, O.; Kargl, D.; Baranov, V.; Mincheva-Nilsson, L. Thermal- and Oxidative Stress Causes Enhanced Release of NKG2D Ligand-Bearing Immunosuppressive Exosomes in Leukemia/Lymphoma T and B Cells. PLoS ONE 2011, 6, e16899. [Google Scholar] [CrossRef]
- Aubertin, K.; Silva, A.K.; Luciani, N.; Espinosa, A.; Djemat, A.; Charue, D.; Gallet, F.; Blanc-Brude, O.; Wilhelm, C. Massive release of extracellular vesicles from cancer cells after photodynamic treatment or chemotherapy. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Parolini, I.; Federici, C.; Raggi, C.; Lugini, L.; Palleschi, S.; De Milito, A.; Coscia, C.; Iessi, E.; Logozzi, M.; Molinari, A.; et al. Microenvironmental pH Is a Key Factor for Exosome Traffic in Tumor Cells. J. Biol. Chem. 2009, 284, 34211–34222. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Guo, W.; Chen, M.; Huang, J.; Yuan, Z.; Zhang, Y.; Wang, M.; Li, P.; Peng, J.; Wang, A.; et al. Extracellular Vesicles and Autophagy in Osteoarthritis. BioMed Res. Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- King, H.W.; Michael, M.Z.; Gleadle, J.M. Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer 2012, 12, 421. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Su, C. Design strategies and application progress of therapeutic exosomes. Theranostics 2019, 9, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Villarroya-beltri, C.; Baixauli, F.; Gutiérrez-vázquez, C. Seminars in Cancer Biology Sorting it out: Regulation of exosome loading. Semin. Cancer Biol. 2014, 28, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Xu, Y.; Wan, Y.; Gao, J.; Chu, Y.; Li, J. Exosomes from adipose-derived mesenchymal stem cells prevent cardiomyocyte apoptosis induced by oxidative stress. Cell Death Discov. 2019, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T. MSC-derived exosomes protect against oxidative stress-induced skin injury via adaptive regulation of the NRF2 defense system. Biomaterials 2020, 257, 120264. [Google Scholar] [CrossRef] [PubMed]
- Kucharzewska, P.; Belting, M. Emerging roles of extracellular vesicles in the adaptive response of tumour cells to microenvironmental stress. J. Extracell. Vesicles 2013, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Buchmaier, B.S.; Bibi, A.; Müller, G.A.; Dihazi, G.H.; Eltoweissy, M.; Kruegel, J.; Dihazi, H. Renal Cells Express Different Forms of Vimentin: The Independent Expression Alteration of these Forms is Important in Cell Resistance to Osmotic Stress and Apoptosis. PLoS ONE 2013, 8, e68301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gao, W.; Zhang, Y.; Cheng, F.; Eriksson, J.E.; Etienne-Manneville, S.; Jiu, Y. Engagement of vimentin intermediate filaments in hypotonic stress. J. Cell. Biochem. 2019, 120, 13168–13176. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, M.; Russell, D.; Morley, S.M.; Davies, A.M.; Lane, E.B. Keratin mutations of epidermolysis bullosa simplex alter the kinetics of stress response to osmotic shock. J. Cell Sci. 2002, 115, 4341–4351. [Google Scholar] [CrossRef] [Green Version]
- Parvanian, S.; Yan, F.; Su, D.; Coelho-Rato, L.S.; Venu, A.P.; Yang, P.; Zou, X.; Jiu, Y.; Chen, H.; Eriksson, J.E.; et al. Exosomal vimentin from adipocyte progenitors accelerates wound healing. Cytoskeleton 2020, 77, 399–413. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, L.; Cui, Y.; Zhou, Y.; Yin, X.; Guo, J.; Zhang, G.; Wang, T.; He, Q.-Y. Cytoskeleton-centric protein transportation by exosomes transforms tumor-favorable macrophages. Oncotarget 2016, 7, 67387–67402. [Google Scholar] [CrossRef] [Green Version]
- AAdolf, A.; Rohrbeck, A.; Münster-Wandowski, A.; Johansson, M.; Kuhn, H.-G.; Kopp, M.A.; Brommer, B.; Schwab, J.M.; Just, I.; Ahnert-Hilger, G.; et al. Release of astroglial vimentin by extracellular vesicles: Modulation of binding and internalization of C3 transferase in astrocytes and neurons. Glia 2018, 67, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Liu, W.; Perincheri, S.; Gunasekaran, M.; Mohanakumar, T. Exosomes expressing the self-antigens myosin and vimentin play an important role in syngeneic cardiac transplant rejection induced by antibodies to cardiac myosin. Am. J. Transplant. 2018, 18, 1626–1635. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Barger, J.F.; Lovat, F.; Gao, M.; Otterson, G.A.; Nana-Sinkam, P. Lung cancer exosomes as drivers of epithelial mesenchymal transition. Oncotarget 2016, 7, 54852–54866. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.L.; Bleaken, B.M.; Romisher, A.R.; Alnwibit, A.A.; Menko, A.S. In wound repair vimentin mediates the transition of mesenchymal leader cells to a myofibroblast phenotype. Mol. Biol. Cell 2018, 29, 1555–1570. [Google Scholar] [CrossRef] [PubMed]
- Eduardsen, K.; Larsen, S.L.; Novak, I.; Lambert, I.H.; Hoffmann, E.K.; Pedersen, S.F. Cell Volume Regulation and Signaling in 3T3-L1 Pre-adipocytes and Adipocytes: On the Possible Roles of Caveolae, Insulin Receptors, FAK and ERK1/2. Cell. Physiol. Biochem. 2011, 28, 1231–1246. [Google Scholar] [CrossRef]
- Chen, B.S.; Li, C.W. Big Mechanisms in System Biology, Chapter 6—Big Mechanism of Information Flow in Cellular Systems in Response to Environmental Stress Signals via System Identification and Data Mining; Academic Press: Cambridge, MA, USA, 2017; pp. 155–248. ISBN 9780128094792. [Google Scholar]
- Sachs, F.; Sivaselvan, M.V. Cell volume control in three dimensions: Water movement without solute movement. J. Gen. Physiol. 2015, 145, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Fader, C.M.; Sánchez, D.; Furlán, M.; Colombo, M.I. Induction of autophagy promotes fusion of multivesicular bodies with autophagic vacuoles in K562 cells. Traffic 2008, 9, 230–250. [Google Scholar] [CrossRef]
- Soria, F.N.; Pampliega, O.; Bourdenx, M.; Meissner, W.G.; Bezard, E.; Dehay, B. Exosomes, an unmasked culprit in neurodegenerative diseases. Front. Neurosci. 2017, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- de Jong, O.G.; Verhaar, M.C.; Chen, Y.; Vader, P.; Gremmels, H.; Posthuma, G.; Schiffelers, R.M.; Gucek, M.; van Balkom, B.W.M. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J. Extracell. Vesicles 2012, 1, 18396. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, L.; Simonsen, U.; Eskildsen-Helmond, Y.; Vorum, H.; Uldbjerg, N.; Honoré, B.; Mulvany, M.J. Proteomics reveals lowering oxygen alters cytoskeletal and endoplasmatic stress proteins in human endothelial cells. Proteomics 2009, 9, 4457–4467. [Google Scholar] [CrossRef] [PubMed]
- Eckes, B.; Colucci-Guyon, E.; Smola, H.; Nodder, S.; Babinet, C.; Krieg, T.; Martin, P. Impaired wound healing in embryonic and adult mice lacking vimentin. J. Cell Sci. 2000, 113, 2455–2462. [Google Scholar] [CrossRef] [PubMed]
- Sung, B.H.; Ketova, T.; Hoshino, D.; Zijlstra, A.; Weaver, A.M. Directional cell movement through tissues is controlled by exosome secretion. Nat. Commun. 2015, 6, 7164. [Google Scholar] [CrossRef] [Green Version]
- Kishi, C.; Minematsu, T.; Huang, L.; Mugita, Y.; Kitamura, A.; Nakagami, G.; Yamane, T.; Yoshida, M.; Noguchi, H.; Funakubo, M.; et al. Hypo-osmotic Shock-Induced Subclinical Inflammation of Skin in a Rat Model of Disrupted Skin Barrier Function. Biol. Res. Nurs. 2014, 17, 135–141. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence 5′→3′ | Reverse Primer Sequence 5′→3′ |
|---|---|---|
| Mouse-β-Actin | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
| Mouse-TNF-α | TCTCATCAGTTCTATGGCCC | GGGAGTAGACAAGGTACAAC |
| Mouse-Granzyme B | TCGACCCTACATGGCCTTAC | TGGGGAATGCATTTTACCAT |
| Mouse-IL-12 | CAGCATGTGTCAATCACGCTAC | TGTGGTCTTCAGCAGGTTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parvanian, S.; Zha, H.; Su, D.; Xi, L.; Jiu, Y.; Chen, H.; Eriksson, J.E.; Cheng, F. Exosomal Vimentin from Adipocyte Progenitors Protects Fibroblasts against Osmotic Stress and Inhibits Apoptosis to Enhance Wound Healing. Int. J. Mol. Sci. 2021, 22, 4678. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094678
Parvanian S, Zha H, Su D, Xi L, Jiu Y, Chen H, Eriksson JE, Cheng F. Exosomal Vimentin from Adipocyte Progenitors Protects Fibroblasts against Osmotic Stress and Inhibits Apoptosis to Enhance Wound Healing. International Journal of Molecular Sciences. 2021; 22(9):4678. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094678
Chicago/Turabian StyleParvanian, Sepideh, Hualian Zha, Dandan Su, Lifang Xi, Yaming Jiu, Hongbo Chen, John E. Eriksson, and Fang Cheng. 2021. "Exosomal Vimentin from Adipocyte Progenitors Protects Fibroblasts against Osmotic Stress and Inhibits Apoptosis to Enhance Wound Healing" International Journal of Molecular Sciences 22, no. 9: 4678. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094678