
Mapping and Characterization of HCMV-Specific Unconventional HLA-E-Restricted CD8 T Cell Populations and Associated NK and T Cell Responses Using HLA/Peptide Tetramers and Spectral Flow Cytometry

 ,
,
Abstract
:1. Introduction
2. Results
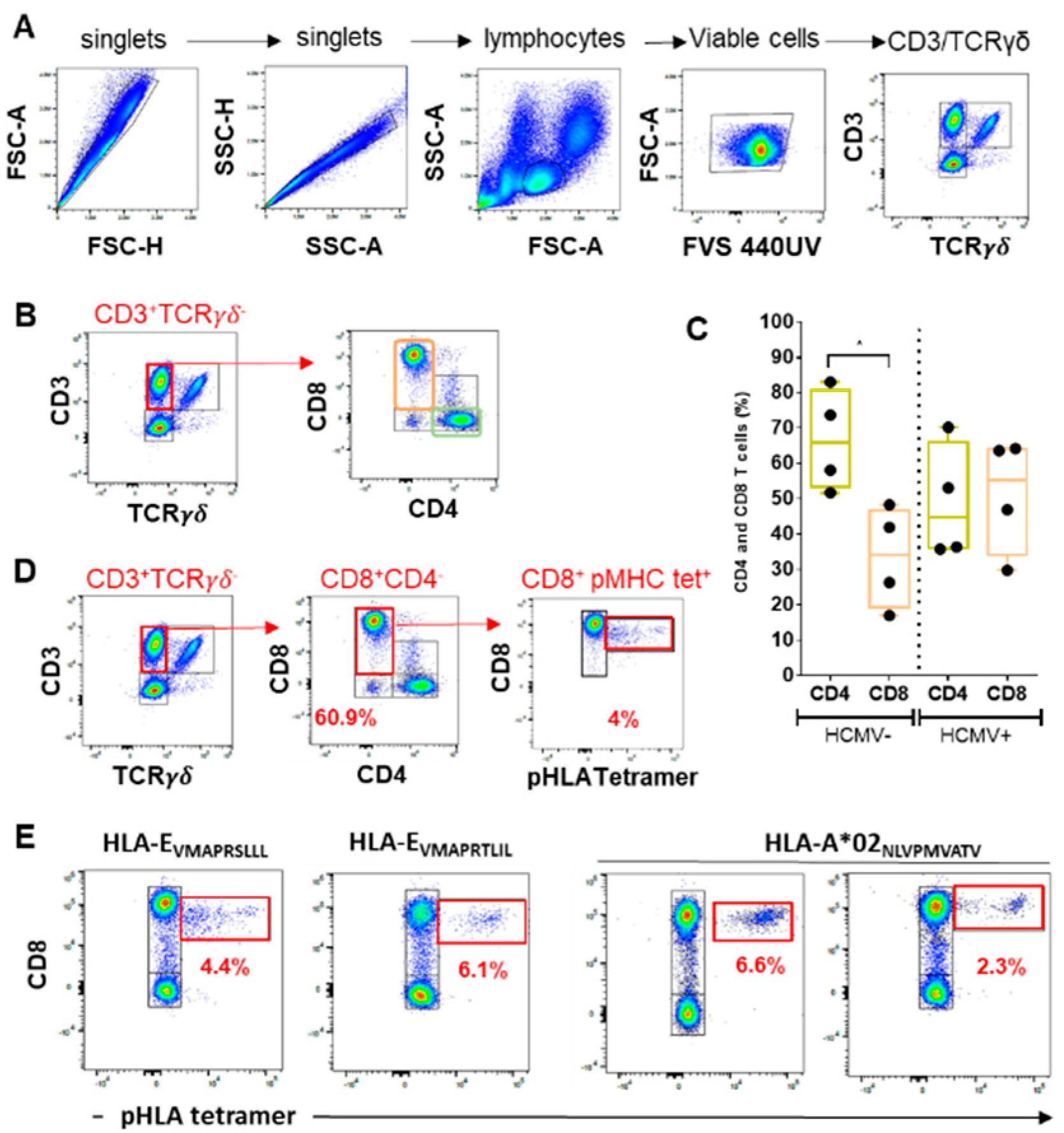
2.1. Quantitative Assessment of CD8 αβT Cells in Response to HCMV Infection: CD8 Inflation and Frequencies of Anti-HCMV Peptide-Specific CD8 T Cells
2.1.1. Lymphocyte Gating
2.1.2. CD4/CD8 Ratio
2.1.3. HCMV Peptide-Specific CD8 T Cell Responses
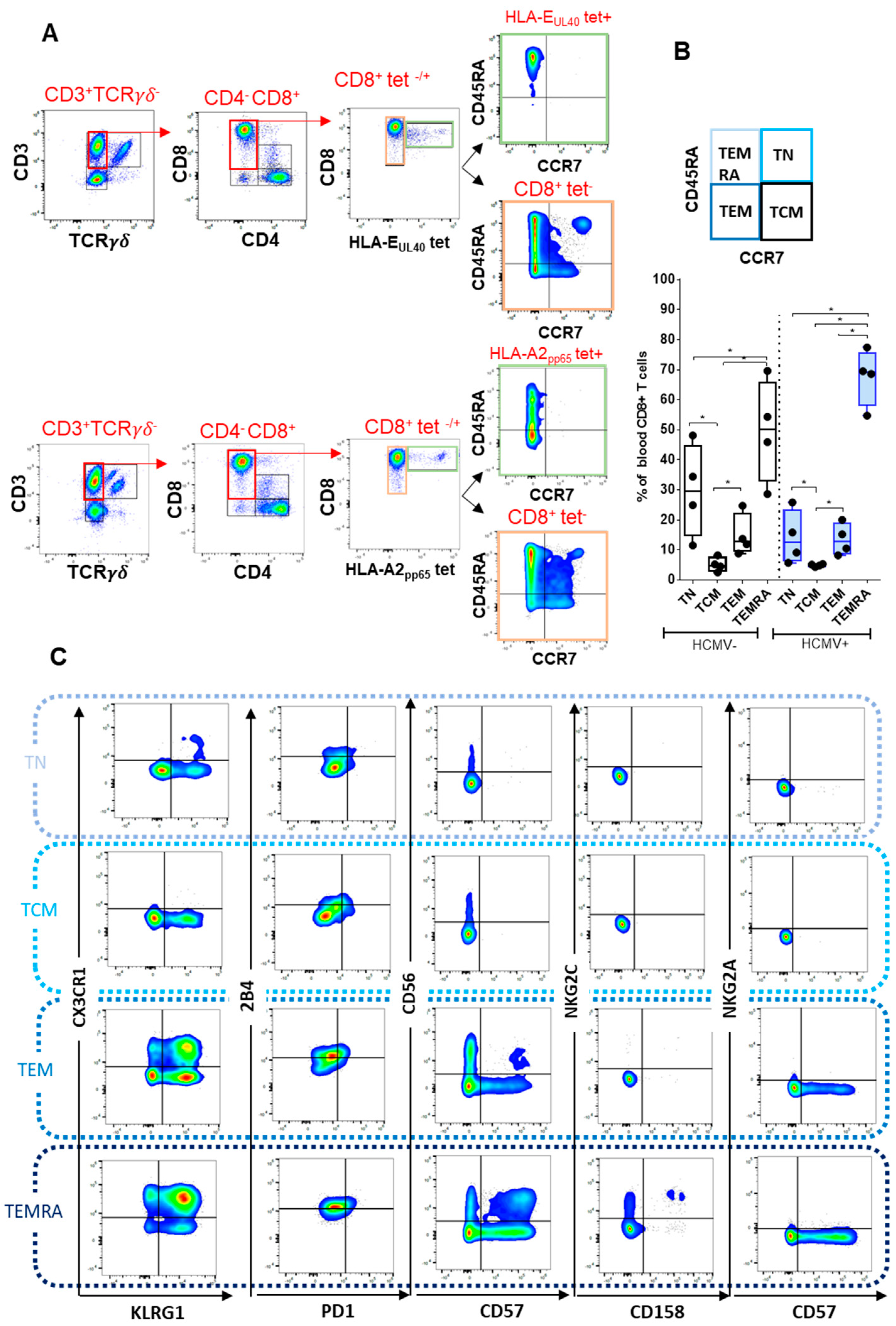
2.2. Immunophenotyping of CD8 T Cell Responses
2.2.1. CD8 T Cell Differentiation
2.2.2. Immunophenotyping of Naive, Central Memory, Effector Memory, and Terminally Differentiated CD8+ T Cell Subsets
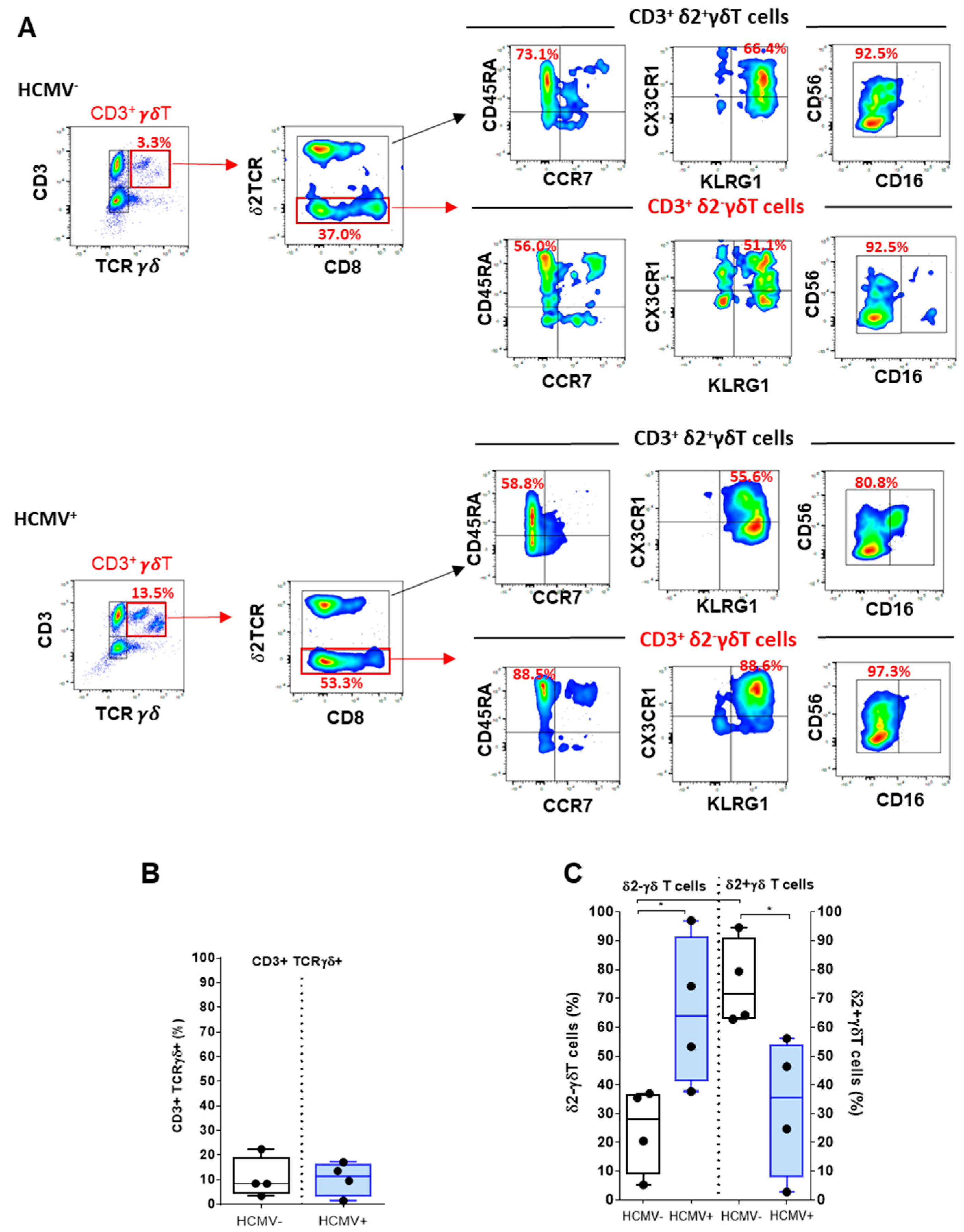
2.3. Deciphering γδT and Vδ2−γδT Cells upon HCMV Infection
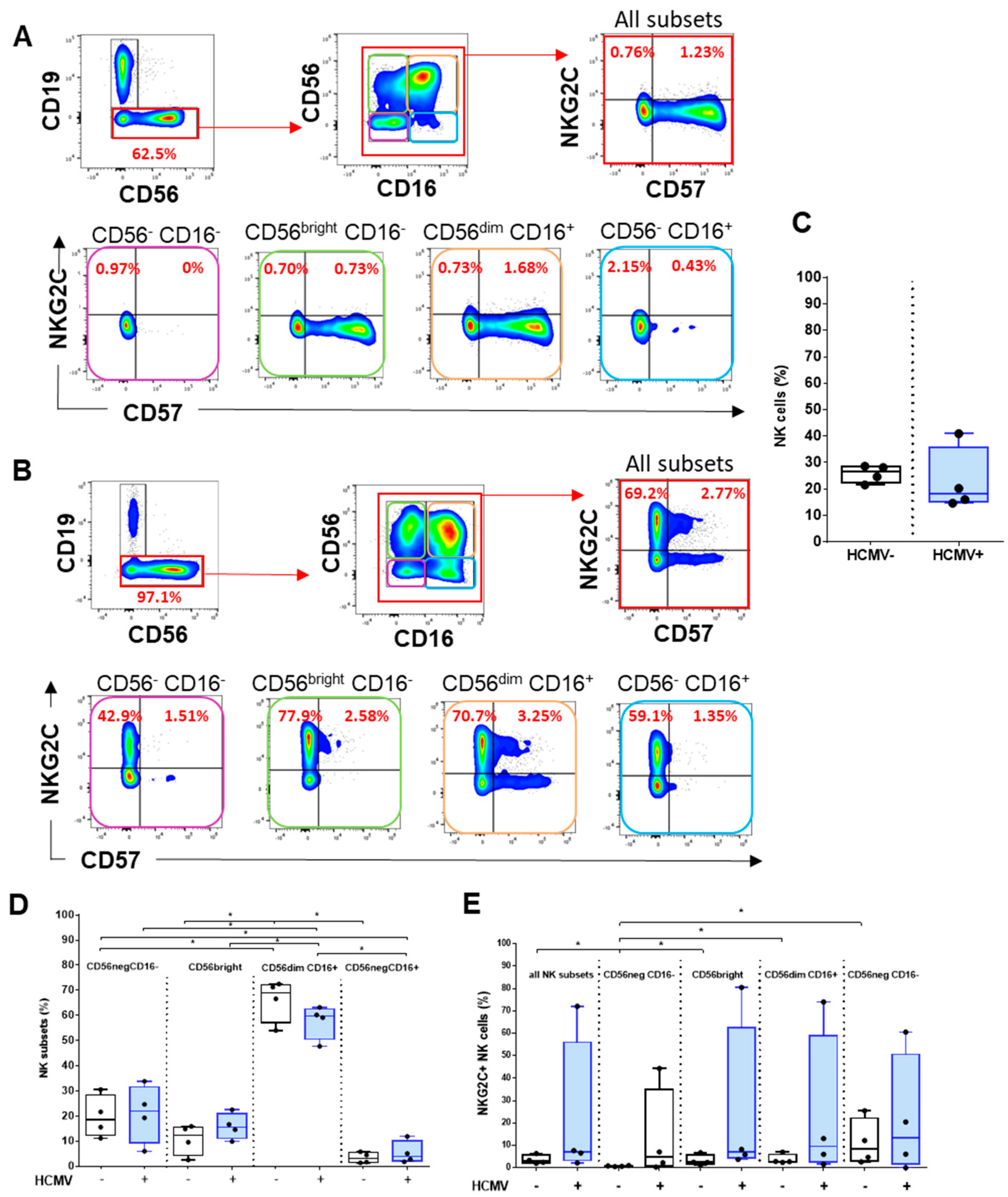
2.4. Analysis of HCMV-Induced NK Cell Subsets
3. Discussion
4. Materials and Methods
4.1. Samples and Reagents
4.2. HLA-EUL40 and HLA-A*02:01pp65 Tetramer Complexes
4.3. Spectral Flow Cytometry: Immunostaining, Acquisition and Post-Acquisition Data Analysis
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boeckh, M.; Geballe, A.P. Cytomegalovirus: Pathogen, paradigm, and puzzle. J. Clin. Investig. 2011, 121, 1673–1680. [Google Scholar] [CrossRef]
- Sylwester, A.W.; Mitchell, B.L.; Edgar, J.B.; Taormina, C.; Pelte, C.; Ruchti, F.; Sleath, P.R.; Grabstein, K.H.; Hosken, N.A.; Kern, F.; et al. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J. Exp. Med. 2005, 202, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, H.; Garrigue, I.; Couzi, L.; Taton, B.; Bachelet, T.; Moreau, J.F.; Dechanet-Merville, J.; Thiebaut, R.; Merville, P. Surveillance of gammadelta T Cells Predicts Cytomegalovirus Infection Resolution in Kidney Transplants. J. Am. Soc. Nephrol. 2016, 27, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Forte, E.; Zhang, Z.; Thorp, E.B.; Hummel, M. Cytomegalovirus Latency and Reactivation: An Intricate Interplay With the Host Immune Response. Front. Cell Infect. Microbiol. 2020, 10, 130. [Google Scholar] [CrossRef]
- Klenerman, P.; Oxenius, A. T cell responses to cytomegalovirus. Nat. Rev. Immunol. 2016, 16, 367–377. [Google Scholar] [CrossRef]
- Khairallah, C.; Dechanet-Merville, J.; Capone, M. gammadelta T Cell-Mediated Immunity to Cytomegalovirus Infection. Front. Immunol. 2017, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Sylwester, A.; Nambiar, K.Z.; Caserta, S.; Klenerman, P.; Picker, L.J.; Kern, F. A new perspective of the structural complexity of HCMV-specific T-cell responses. Mech. Ageing Dev. 2016, 158, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, C.; Pietra, G.; Falco, M.; Millo, E.; Mazzarino, P.; Biassoni, R.; Moretta, A.; Moretta, L.; Mingari, M.C. Identification of HLA-E-specific alloreactive T lymphocytes: A cell subset that undergoes preferential expansion in mixed lymphocyte culture and displays a broad cytolytic activity against allogeneic cells. Proc. Natl. Acad. Sci. USA 2002, 99, 11328–11333. [Google Scholar] [CrossRef] [Green Version]
- Pietra, G.; Romagnani, C.; Mazzarino, P.; Falco, M.; Millo, E.; Moretta, A.; Moretta, L.; Mingari, M.C. HLA-E-restricted recognition of cytomegalovirus-derived peptides by human CD8+ cytolytic T lymphocytes. Proc. Natl. Acad. Sci. USA 2003, 100, 10896–10901. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, L.C.; Westall, G.P.; Widjaja, J.M.; Mifsud, N.A.; Nguyen, T.H.; Meehan, A.C.; Kotsimbos, T.C.; Brooks, A.G. The Presence of HLA-E-Restricted, CMV-Specific CD8+ T Cells in the Blood of Lung Transplant Recipients Correlates with Chronic Allograft Rejection. PLoS ONE 2015, 10, e0135972. [Google Scholar] [CrossRef] [Green Version]
- Jouand, N.; Bressollette-Bodin, C.; Gerard, N.; Giral, M.; Guerif, P.; Rodallec, A.; Oger, R.; Parrot, T.; Allarrd, M.; Cesbron-Gautier, A.; et al. HCMV triggers frequent and persistent UL40-specific unconventional HLA-E-restricted CD8 T-cell responses with potential autologous and allogeneic peptide recognition. PLoS Pathog. 2018, 14, e1007041. [Google Scholar] [CrossRef]
- Pietra, G.; Romagnani, C.; Falco, M.; Vitale, M.; Castriconi, R.; Pende, D.; Millo, E.; Anfossi, S.; Biassoni, R.; Moretta, L.; et al. The analysis of the natural killer-like activity of human cytolytic T lymphocytes revealed HLA-E as a novel target for TCR alpha/beta-mediated recognition. Eur. J. Immunol. 2001, 31, 3687–3693. [Google Scholar] [CrossRef]
- Allard, M.; Tonnerre, P.; Nedellec, S.; Oger, R.; Morice, A.; Guilloux, Y.; Houssaint, E.; Charreau, B.; Gervois, N. HLA-E-restricted cross-recognition of allogeneic endothelial cells by CMV-associated CD8 T cells: A potential risk factor following transplantation. PLoS ONE 2012, 7, e50951. [Google Scholar] [CrossRef] [Green Version]
- Altman, J.D.; Moss, P.A.; Goulder, P.J.; Barouch, D.H.; McHeyzer-Williams, M.G.; Bell, J.I.; McMichael, A.J.; Davis, M.M. Phenotypic analysis of antigen-specific T lymphocytes. Science 1996, 274, 94–96. [Google Scholar] [CrossRef]
- Hadrup, S.R.; Bakker, A.H.; Shu, C.J.; Andersen, R.S.; van Veluw, J.; Hombrink, P.; Castermans, E.; Straten, P.T.; Blank, C.; Haanen, J.B.; et al. Parallel detection of antigen-specific T-cell responses by multidimensional encoding of MHC multimers. Nat. Methods 2009, 6, 520–526. [Google Scholar] [CrossRef]
- Newell, E.W.; Klein, L.O.; Yu, W.; Davis, M.M. Simultaneous detection of many T-cell specificities using combinatorial tetramer staining. Nat. Methods 2009, 6, 497–499. [Google Scholar] [CrossRef]
- Newell, E.W.; Sigal, N.; Nair, N.; Kidd, B.A.; Greenberg, H.B.; Davis, M.M. Combinatorial tetramer staining and mass cytometry analysis facilitate T-cell epitope mapping and characterization. Nat. Biotechnol. 2013, 31, 623–629. [Google Scholar] [CrossRef]
- Perfetto, S.P.; Chattopadhyay, P.K.; Roederer, M. Seventeen-colour flow cytometry: Unravelling the immune system. Nat. Rev. Immunol. 2004, 4, 648–655. [Google Scholar] [CrossRef]
- Nolan, J.P.; Condello, D. Spectral flow cytometry. Curr. Protoc. Cytom. 2013, 1, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Karrer, U.; Sierro, S.; Wagner, M.; Oxenius, A.; Hengel, H.; Koszinowski, U.H.; Phillips, R.E.; Klenerman, P. Memory inflation: Continuous accumulation of antiviral CD8+ T cells over time. J. Immunol. 2003, 170, 2022–2029. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Olson, D.; Simons, B.C.; Conrad, J.A.; Smith, R.M.; Barnett, L.; Lorey, S.L.; Duncan, C.B.; Ramalingam, R.; Kalams, S.A. Clonal expansion and TCR-independent differentiation shape the HIV-specific CD8+ effector-memory T-cell repertoire in vivo. Blood 2010, 116, 396–405. [Google Scholar] [CrossRef]
- O’Hara, G.A.; Welten, S.P.; Klenerman, P.; Arens, R. Memory T cell inflation: Understanding cause and effect. Trends Immunol. 2012, 33, 84–90. [Google Scholar] [CrossRef]
- Van den Berg, S.P.H.; Pardieck, I.N.; Lanfermeijer, J.; Sauce, D.; Klenerman, P.; van Baarle, D.; Arens, R. The hallmarks of CMV-specific CD8 T-cell differentiation. Med. Microbiol. Immunol. 2019, 208, 365–573. [Google Scholar] [CrossRef] [Green Version]
- Derhovanessian, E.; Maier, A.B.; Hahnel, K.; Beck, R.; de Craen, A.J.M.; Slagboom, E.P.; Westendorp, R.G.J.; Pawelec, G. Infection with cytomegalovirus but not herpes simplex virus induces the accumulation of late-differentiated CD4+ and CD8+ T-cells in humans. J. Gen. Virol. 2011, 92, 2746–2756. [Google Scholar] [CrossRef]
- Dechanet, J.; Merville, P.; Berge, F.; Bone-Mane, G.; Taupin, J.L.; Michel, P.; Joly, P.; Bonneville, M.; Potaux, L.; Moreau, J.-F.; et al. Major expansion of gammadelta T lymphocytes following cytomegalovirus infection in kidney allograft recipients. J. Infect. Dis. 1999, 179, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Stankovic, S.; Davey, M.S.; Shaw, E.M.; von Borstel, A.; Cristiano, Y.; Levvey, B.J.; Rossjohn, J.; Westall, G.P.; Snell, G.I.; Brooks, A.G.; et al. Cytomegalovirus replication is associated with enrichment of distinct gammadelta T cell subsets following lung transplantation: A novel therapeutic approach? J. Heart Lung Transpl. 2020, 39, 1300–1312. [Google Scholar] [CrossRef]
- Alejenef, A.; Pachnio, A.; Halawi, M.; Christmas, S.E.; Moss, P.A.; Khan, N. Cytomegalovirus drives Vdelta2neg gammadelta T cell inflation in many healthy virus carriers with increasing age. Clin. Exp. Immunol. 2014, 176, 418–428. [Google Scholar] [CrossRef]
- Couzi, L.; Pitard, V.; Moreau, J.F.; Merville, P.; Dechanet-Merville, J. Direct and Indirect Effects of Cytomegalovirus-Induced gammadelta T Cells after Kidney Transplantation. Front. Immunol. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Della Chiesa, M.; Marcenaro, E.; Sivori, S.; Carlomagno, S.; Pesce, S.; Moretta, A. Human NK cell response to pathogens. Semin. Immunol. 2014, 26, 152–160. [Google Scholar] [CrossRef]
- Cichicki, F.; Schlums, H.; Theorell, J.; Tesi, B.; Miller, J.S.; Ljunggren, H.G.; Bryceson, Y.T. Diversification and Functional Specialization of Human NK Cell Subsets. Curr. Top. Microbiol. Immunol. 2016, 395, 63–94. [Google Scholar]
- Freud, A.G.; Mundy-Bosse, B.L.; Yu, J.; Caligiuri, M.A. The Broad Spectrum of Human Natural Killer Cell Diversity. Immunity 2017, 47, 820–833. [Google Scholar] [CrossRef] [Green Version]
- Roberto, A.; Di Vito, C.; Zaghi, E.; Mazza, E.M.C.; Capucetti, A.; Calvi, M.; Tentorio, P.; Zanon, V.; Sarina, B.; Mariotti, J.; et al. The early expansion of anergic NKG2A(pos)/CD56(dim)/CD16(neg) natural killer represents a therapeutic target in haploidentical hematopoietic stem cell transplantation. Haematologica 2018, 103, 1390–1402. [Google Scholar] [CrossRef] [Green Version]
- Di Vito, C.; Mikulak, J.; Mavilio, D. On the Way to Become a Natural Killer Cell. Front. Immunol. 2019, 10, 1812. [Google Scholar] [CrossRef]
- Ferrer-Font, L.; Mayer, J.U.; Old, S.; Hermans, I.F.; Irish, J.; Price, K.M. High-Dimensional Data Analysis Algorithms Yield Comparable Results for Mass Cytometry and Spectral Flow Cytometry Data. Cytom. A 2020, 97, 824–831. [Google Scholar] [CrossRef]
- Ferrer-Font, L.; Pellefigues, C.; Mayer, J.U.; Small, S.J.; Jaimes, M.C.; Price, K.M. Panel Design and Optimization for High-Dimensional Immunophenotyping Assays Using Spectral Flow Cytometry. Curr. Protoc. Cytom. 2020, 92, e70. [Google Scholar] [CrossRef]
- Braud, V.M.; Allan, D.S.; O’Callaghan, C.A.; Soderstrom, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef]
- Arlettaz, L.; Villard, J.; de Rham, C.; Degermann, S.; Chapuis, B.; Huard, B.; Roosnek, E. Activating CD94:NKG2C and inhibitory CD94:NKG2A receptors are expressed by distinct subsets of committed CD8+ TCR alphabeta lymphocytes. Eur. J. Immunol. 2004, 34, 3456–3464. [Google Scholar] [CrossRef]
- Jiang, H.; Canfield, S.M.; Gallagher, M.P.; Jiang, H.H.; Jiang, Y.; Zheng, Z.; Chess, L. HLA-E-restricted regulatory CD8(+) T cells are involved in development and control of human autoimmune type 1 diabetes. J. Clin. Investig. 2010, 120, 3641–3650. [Google Scholar] [CrossRef] [PubMed]
- La Manna, M.P.; Orlando, V.; Prezzemolo, T.; Di Carlo, P.; Cascio, A.; Delogu, G.; Poli, G.; Sullivan, L.C.; Brooks, A.G.; Dieli, F.; et al. HLA-E-restricted CD8(+) T Lymphocytes Efficiently Control Mycobacterium tuberculosis and HIV-1 Coinfection. Am. J. Respir. Cell Mol. Biol. 2020, 62, 430–439. [Google Scholar] [CrossRef]
- Coupel, S.; Moreau, A.; Hamidou, M.; Horejsi, V.; Soulillou, J.P.; Charreau, B. Expression and release of soluble HLA-E is an immunoregulatory feature of endothelial cell activation. Blood 2007, 109, 2806–2814. [Google Scholar] [CrossRef] [Green Version]
- Holderried, T.A.; Lang, P.A.; Kim, H.J.; Cantor, H. Genetic disruption of CD8+ Treg activity enhances the immune response to viral infection. Proc. Natl. Acad. Sci. USA 2013, 110, 21089–21094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Verges, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.-M.; Norris, P.J.; et al. Expansion of a unique CD57(+)NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratama, J.W.; Boeckh, M.; Nakamura, R.; Cornelissen, J.J.; Brooimans, R.A.; Zaia, J.A.; Forman, S.J.; Gaal, K.; Bray, K.R.; Gasior, G.H.; et al. Immune monitoring with iTAg MHC Tetramers for prediction of recurrent or persistent cytomegalovirus infection or disease in allogeneic hematopoietic stem cell transplant recipients: A prospective multicenter study. Blood 2010, 116, 1655–1662. [Google Scholar] [CrossRef]
- Jarque, M.; Crespo, E.; Melilli, E.; Gutierrez, A.; Moreso, F.; Guirado, L.; Revuelta, I.; Montero, N.; Torras, J.; Reira, L.; et al. Cellular Immunity to Predict the Risk of Cytomegalovirus Infection in Kidney Transplantation: A Prospective, Interventional, Multicenter Clinical Trial. Clin. Infect. Dis. 2020, 71, 2375–2385. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Λ Excitation (nm) | Fluorophore | Λ Emission (nm) | Cytometer Detector | Antigen | Antibody Clone | Dilution | Source |
|---|---|---|---|---|---|---|---|
| 355 | BUV395 | 395 | UV2 | CD45RA | 5H9 | 1/80e | BD Bioscience |
| Fixable Viability Dye UV440 | 436 | UV6 | Viability | / | 1/2000e | BD Bioscience | |
| BUV496 | 496 | UV7 | CD16 | 3G8 | 1/160e | BD Bioscience | |
| BUV563 | 564 | UV9 | NKG2C | 134591 | 1/20e | BD Bioscience | |
| BUV737 | 735 | UV14 | CD56 | NCAM16.2 | 1/20e | BD Bioscience | |
| BUV805 | 803 | UV16 | CD8 | SK1 | 1/80e | BD Bioscience | |
| 405 | BV421 | 421 | V1 | CCR7 | G043H7 | 1/20e | Biolegend |
| VioBlue | 452 | V3 | KLRG1 | REA261 | 1/50e | Myltenyi | |
| BV510 | 510 | V7 | CD3 | OKT3 | 1/20e | Biolegend | |
| BV570 | 570 | V8 | CD4 | RPA-T4 | 1/80e | Biolegend | |
| BV605 | 603 | V10 | 2B4 | C1.7 | 1/80e | Biolegend | |
| BV785 | 785 | V15 | PD-1 | EH12.2H7 | 1/10e | Biolegend | |
| 488 | FITC | 520 | B2 | CD57 | HNK-1 | 1/160e | Biolegend |
| PerCPeFluor710 | 710 | B10 | TCRgd | B1.1 | 1/20e | Thermo Fisher | |
| 561 | PE | 576 | YG1 | CD158 | HP-MA4 | 1/40e | Biolegend |
| AlexaFluor594 | 617 | YG3 | NKG2A | 131411 | 1/20e | Bio-Techne | |
| PE-Cy7 | 781 | YG9 | CX3CR1 | 2A9-1 | 1/20e | Biolegend | |
| 640 | SparkNir685 | 685 | R3 | CD19 | HIB19 | 1/80e | Biolegend |
| APC Fire750 APC | 787 660 | R7 R1 | TCRgd2 Streptavidin HLApeptide | B6 / | 1/160e 1/11e | Biolegend BD Bioscience | |
| / | / | / | / | CD94 | HP-3D9 | 30 µg/mL | BD Bioscience |
| / | / | / | / | Fc block | Fc1.3216 | 1/100e | BD Bioscience |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rousselière, A.; Delbos, L.; Bressollette, C.; Berthaume, M.; Charreau, B. Mapping and Characterization of HCMV-Specific Unconventional HLA-E-Restricted CD8 T Cell Populations and Associated NK and T Cell Responses Using HLA/Peptide Tetramers and Spectral Flow Cytometry. Int. J. Mol. Sci. 2022, 23, 263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010263
Rousselière A, Delbos L, Bressollette C, Berthaume M, Charreau B. Mapping and Characterization of HCMV-Specific Unconventional HLA-E-Restricted CD8 T Cell Populations and Associated NK and T Cell Responses Using HLA/Peptide Tetramers and Spectral Flow Cytometry. International Journal of Molecular Sciences. 2022; 23(1):263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010263
Chicago/Turabian StyleRousselière, Amélie, Laurence Delbos, Céline Bressollette, Maïlys Berthaume, and Béatrice Charreau. 2022. "Mapping and Characterization of HCMV-Specific Unconventional HLA-E-Restricted CD8 T Cell Populations and Associated NK and T Cell Responses Using HLA/Peptide Tetramers and Spectral Flow Cytometry" International Journal of Molecular Sciences 23, no. 1: 263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010263