
A Growing Link between Circadian Rhythms, Type 2 Diabetes Mellitus and Alzheimer’s Disease


Abstract
:1. Introduction
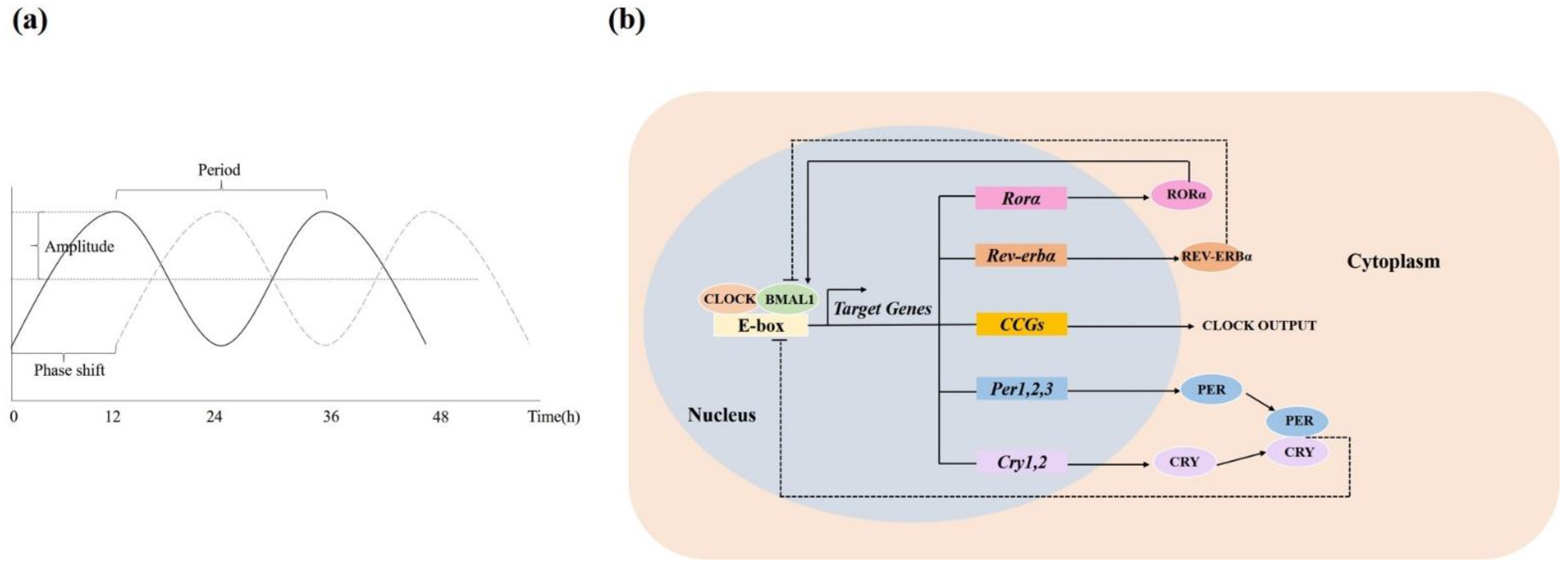
2. Circadian Rhythms
3. T2DM and Circadian Disruption
3.1. The Diurnal Rhythm of Glucose Metabolism
3.2. T2DM Animals and Patients Show Circadian Disruption
3.2.1. Circadian Disruption Occurs in Diabetic Animal Models
3.2.2. T2DM Patients Show Disruptive Circadian Rhythms
3.3. Circadian Disruption Contributes to T2DM
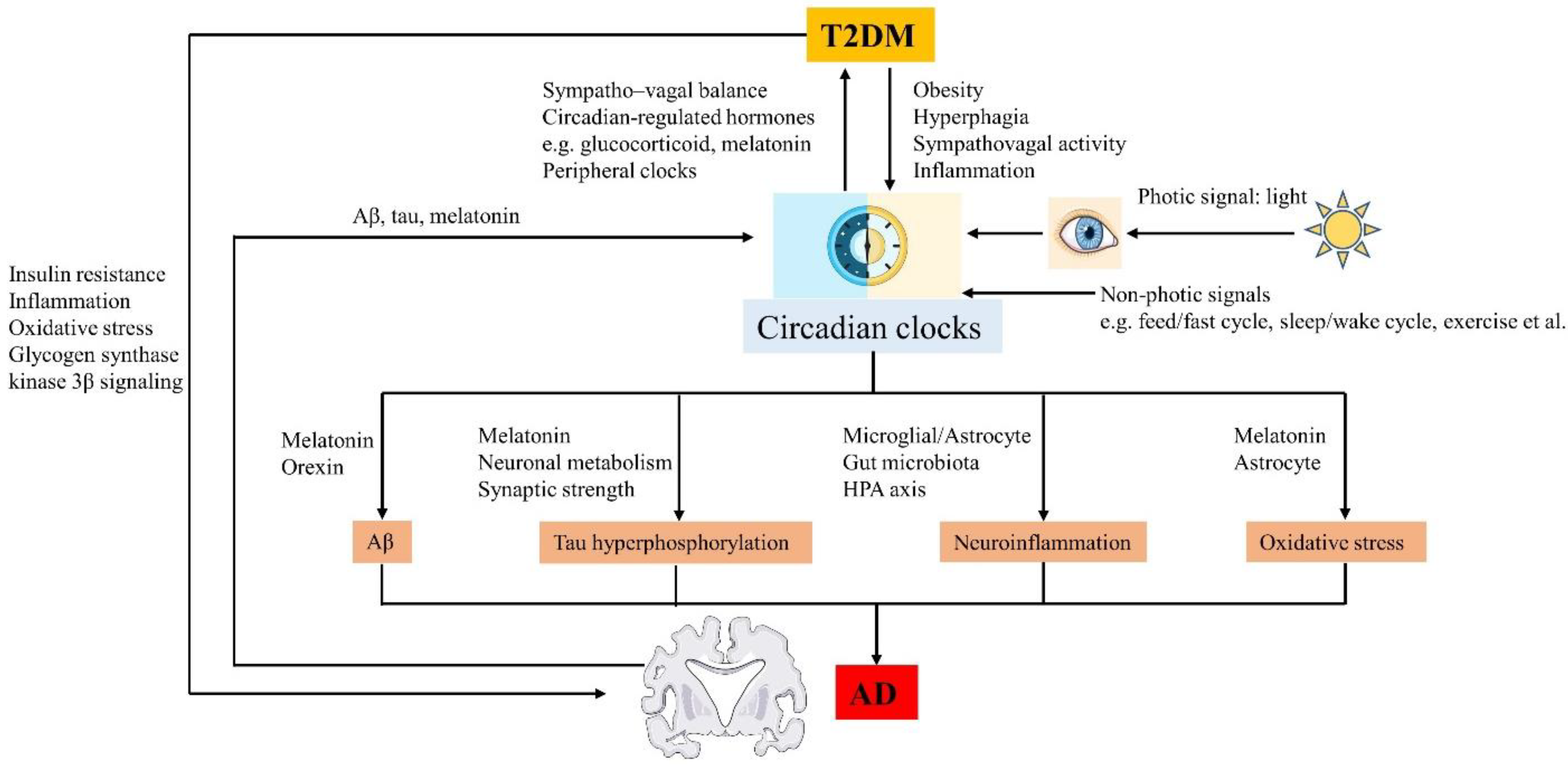
4. Circadian Disruption and AD
4.1. AD Presents Circadian Disruption
4.2. Effects of Circadian Disruption on AD
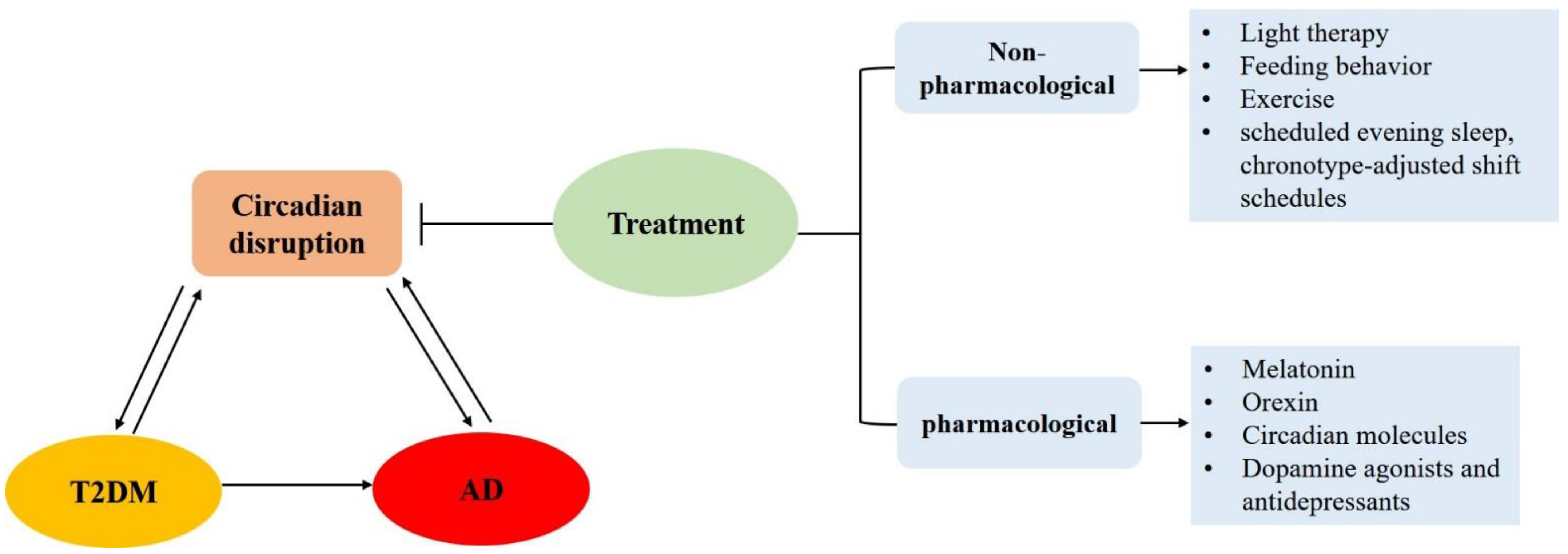
5. Treatment for T2DM and AD Targeting Circadian Rhythms
5.1. Non-Pharmacological Treatment
5.1.1. Light Therapy
5.1.2. Feeding Behavior
5.1.3. Exercise
5.1.4. Other Lifestyle Interventions
5.2. Pharmacological Treatment
5.2.1. Melatonin
5.2.2. Orexin
5.2.3. Circadian Molecules
5.2.4. Other Drugs
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prince, M.; Bryce, R.; Albanese, E.; Wimo, A.; Ribeiro, W.; Ferri, C.P. The global prevalence of dementia: A systematic review and metaanalysis. Alzheimer’s Dement. 2013, 9, 63–75.e62. [Google Scholar] [CrossRef]
- Scheltens, P.; Blennow, K.; Breteler, M.M.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet. 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.; Magliano, D.J.; Bennett, P.H. Diabetes mellitus statistics on prevalence and mortality: Facts and fallacies. Nat. Rev. Endocrinol. 2016, 12, 616–622. [Google Scholar] [CrossRef]
- Biessels, G.J.; Despa, F. Cognitive decline and dementia in diabetes mellitus: Mechanisms and clinical implications. Nat. Rev. Endocrinol. 2018, 14, 591–604. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, C.; Hua, S.; Liao, H.; Wang, M.; Xiong, Y.; Cao, F. An updated meta-analysis of cohort studies: Diabetes and risk of Alzheimer’s disease. Diabetes Res. Clin. Pract. 2017, 124, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, T. Epidemiological Evidence of the Relationship Between Diabetes and Dementia. Adv. Exp. Med. Biol. 2019, 1128, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; Thirumala, V.; Reddy, P.H. Is Alzheimer’s disease a Type 3 Diabetes? A critical appraisal. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1078–1089. [Google Scholar] [CrossRef]
- Maiese, K. Cognitive impairment with diabetes mellitus and metabolic disease: Innovative insights with the mechanistic target of rapamycin and circadian clock gene pathways. Expert Rev. Clin. Pharmacol. 2020, 13, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Loehfelm, A.; Boucsein, A.; Pretz, D.; Tups, A. Timing Matters: Circadian Effects on Energy Homeostasis and Alzheimer’s Disease. Trends Endocrinol. Metab. TEM 2019, 30, 132–143. [Google Scholar] [CrossRef]
- Schibler, U.; Sassone-Corsi, P. A web of circadian pacemakers. Cell 2002, 111, 919–922. [Google Scholar] [CrossRef] [Green Version]
- Arendt, J. Biological rhythms during residence in polar regions. Chronobiol. Int. 2012, 29, 379–394. [Google Scholar] [CrossRef] [Green Version]
- Leng, Y.; Musiek, E.S.; Hu, K.; Cappuccio, F.P.; Yaffe, K. Association between circadian rhythms and neurodegenerative diseases. Lancet Neurol. 2019, 18, 307–318. [Google Scholar] [CrossRef]
- Albrecht, U. Timing to perfection: The biology of central and peripheral circadian clocks. Neuron 2012, 74, 246–260. [Google Scholar] [CrossRef] [Green Version]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Hood, S.; Amir, S. Neurodegeneration and the Circadian Clock. Front. Aging Neurosci. 2017, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilking, M.; Ndiaye, M.; Mukhtar, H.; Ahmad, N. Circadian rhythm connections to oxidative stress: Implications for human health. Antioxid. Redox Signal. 2013, 19, 192–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musiek, E.S.; Holtzman, D.M. Mechanisms linking circadian clocks, sleep, and neurodegeneration. Science. 2016, 354, 1004–1008. [Google Scholar] [CrossRef] [Green Version]
- Maury, E. Off the Clock: From Circadian Disruption to Metabolic Disease. Int. J. Mol. Sci. 2019, 20, 1597. [Google Scholar] [CrossRef] [Green Version]
- Javeed, N.; Matveyenko, A.V. Circadian Etiology of Type 2 Diabetes Mellitus. Physiology 2018, 33, 138–150. [Google Scholar] [CrossRef]
- Poggiogalle, E.; Jamshed, H.; Peterson, C.M. Circadian regulation of glucose, lipid, and energy metabolism in humans. Metab. Clin. Exp. 2018, 84, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Ramsey, K.M.; Marcheva, B.; Bass, J. Circadian rhythms, sleep, and metabolism. J. Clin. Investig. 2011, 121, 2133–2141. [Google Scholar] [CrossRef]
- Jarrett, R.J.; Keen, H. Diurnal variation of oral glucose tolerance: A possible pointer to the evolution of diabetes mellitus. Br. Med. J. 1969, 2, 341–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, K.F.; Nestel, P.J. Diurnal variation in glucose tolerance and in insulin secretion in man. Diabetes 1973, 22, 333–348. [Google Scholar] [CrossRef]
- Aparicio, N.J.; Puchulu, F.E.; Gagliardino, J.J.; Ruiz, M.; Llorens, J.M.; Ruiz, J.; Lamas, A.; De Miguel, R. Circadian variation of the blood glucose, plasma insulin and human growth hormone levels in response to an oral glucose load in normal subjects. Diabetes 1974, 23, 132–137. [Google Scholar] [CrossRef]
- Jarrett, R.J. Circadian variation in blood glucose levels, in glucose tolerance and in plasma immunoreactive insulin levels. Acta Diabetol. Lat. 1972, 9, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Van Cauter, E.; Polonsky, K.S.; Scheen, A.J. Roles of circadian rhythmicity and sleep in human glucose regulation. Endocr. Rev. 1997, 18, 716–738. [Google Scholar] [CrossRef] [Green Version]
- Pinkhasov, B.B.; Selyatinskaya, V.G.; Astrakhantseva, E.L.; Anufrienko, E.V. Circadian Rhythms of Carbohydrate Metabolism in Women with Different Types of Obesity. Bull. Exp. Biol. Med. 2016, 161, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Hulmán, A.; Færch, K.; Vistisen, D.; Karsai, J.; Nyári, T.A.; Tabák, A.G.; Brunner, E.J.; Kivimäki, M.; Witte, D.R. Effect of time of day and fasting duration on measures of glycaemia: Analysis from the Whitehall II Study. Diabetologia 2013, 56, 294–297. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Ader, M.; Bray, G.A.; Bergman, R.N. Diurnal variation in glucose tolerance. Cyclic suppression of insulin action and insulin secretion in normal-weight, but not obese, subjects. Diabetes 1992, 41, 750–759. [Google Scholar] [CrossRef]
- Morris, C.J.; Yang, J.N.; Garcia, J.I.; Myers, S.; Bozzi, I.; Wang, W.; Buxton, O.M.; Shea, S.A.; Scheer, F.A. Endogenous circadian system and circadian misalignment impact glucose tolerance via separate mechanisms in humans. Proc. Natl. Acad. Sci. USA 2015, 112, E2225–E2234. [Google Scholar] [CrossRef] [Green Version]
- Ruiter, M.; La Fleur, S.E.; van Heijningen, C.; van der Vliet, J.; Kalsbeek, A.; Buijs, R.M. The daily rhythm in plasma glucagon concentrations in the rat is modulated by the biological clock and by feeding behavior. Diabetes 2003, 52, 1709–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Lozano, M.; Mingomataj, E.L.; Wu, W.K.; Ridout, S.A.; Brubaker, P.L. Circadian secretion of the intestinal hormone GLP-1 by the rodent L cell. Diabetes 2014, 63, 3674–3685. [Google Scholar] [CrossRef] [Green Version]
- Mason, I.C.; Qian, J.; Adler, G.K.; Scheer, F. Impact of circadian disruption on glucose metabolism: Implications for type 2 diabetes. Diabetologia 2020, 63, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radziuk, J.; Pye, S. Diurnal rhythm in endogenous glucose production is a major contributor to fasting hyperglycaemia in type 2 diabetes. Suprachiasmatic deficit or limit cycle behaviour? Diabetologia 2006, 49, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosbellet, E.; Dumont, S.; Schuster-Klein, C.; Guardiola-Lemaitre, B.; Pevet, P.; Criscuolo, F.; Challet, E. Circadian phenotyping of obese and diabetic db/db mice. Biochimie 2016, 124, 198–206. [Google Scholar] [CrossRef]
- Su, W.; Guo, Z.; Randall, D.C.; Cassis, L.; Brown, D.R.; Gong, M.C. Hypertension and disrupted blood pressure circadian rhythm in type 2 diabetic db/db mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1634–H1641. [Google Scholar] [CrossRef] [Green Version]
- Hou, T.; Su, W.; Guo, Z.; Gong, M.C. A Novel Diabetic Mouse Model for Real-Time Monitoring of Clock Gene Oscillation and Blood Pressure Circadian Rhythm. J. Biol. Rhythm. 2019, 34, 51–68. [Google Scholar] [CrossRef]
- Senador, D.; Kanakamedala, K.; Irigoyen, M.C.; Morris, M.; Elased, K.M. Cardiovascular and autonomic phenotype of db/db diabetic mice. Exp. Physiol. 2009, 94, 648–658. [Google Scholar] [CrossRef] [Green Version]
- Laposky, A.D.; Bradley, M.A.; Williams, D.L.; Bass, J.; Turek, F.W. Sleep-wake regulation is altered in leptin-resistant (db/db) genetically obese and diabetic mice. Am. J. Physiology Regul. Integr. Comp. Physiol. 2008, 295, R2059–R2066. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Xie, Z.; Guo, Z.; Duncan, M.J.; Lutshumba, J.; Gong, M.C. Altered clock gene expression and vascular smooth muscle diurnal contractile variations in type 2 diabetic db/db mice. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H621–H633. [Google Scholar] [CrossRef] [Green Version]
- Caton, P.W.; Kieswich, J.; Yaqoob, M.M.; Holness, M.J.; Sugden, M.C. Metformin opposes impaired AMPK and SIRT1 function and deleterious changes in core clock protein expression in white adipose tissue of genetically-obese db/db mice. Diabetes Obes. Metab. 2011, 13, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Akiyama, M.; Kuriyama, K.; Sudo, M.; Moriya, T.; Shibata, S. Night-time restricted feeding normalises clock genes and Pai-1 gene expression in the db/db mouse liver. Diabetologia 2004, 47, 1425–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branecky, K.L.; Niswender, K.D.; Pendergast, J.S. Disruption of Daily Rhythms by High-Fat Diet Is Reversible. PloS ONE 2015, 10, e0137970. [Google Scholar] [CrossRef]
- Pendergast, J.S.; Branecky, K.L.; Yang, W.; Ellacott, K.L.; Niswender, K.D.; Yamazaki, S. High-fat diet acutely affects circadian organisation and eating behavior. Eur. J. Neurosci. 2013, 37, 1350–1356. [Google Scholar] [CrossRef] [Green Version]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-fat diet disrupts behavioral and molecular circadian rhythms in mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, M.C.; Yang, S.C.; Tseng, H.L.; Hwang, L.L.; Chen, C.T.; Shieh, K.R. Abnormal expressions of circadian-clock and circadian clock-controlled genes in the livers and kidneys of long-term, high-fat-diet-treated mice. Int. J. Obes. 2010, 34, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Woodie, L.N.; Johnson, R.M.; Ahmed, B.; Fowler, S.; Haynes, W.; Carmona, B.; Reed, M.; Suppiramaniam, V.; Greene, M.W. Western diet-induced obesity disrupts the diurnal rhythmicity of hippocampal core clock gene expression in a mouse model. Brain Behav. Immun. 2020, 88, 815–825. [Google Scholar] [CrossRef]
- Yang, S.C.; Tseng, H.L.; Shieh, K.R. Circadian-clock system in mouse liver affected by insulin resistance. Chronobiol. Int. 2013, 30, 796–810. [Google Scholar] [CrossRef]
- Kuriyama, K.; Sasahara, K.; Kudo, T.; Shibata, S. Daily injection of insulin attenuated impairment of liver circadian clock oscillation in the streptozotocin-treated diabetic mouse. FEBS Lett. 2004, 572, 206–210. [Google Scholar] [CrossRef] [Green Version]
- Young, M.E.; Wilson, C.R.; Razeghi, P.; Guthrie, P.H.; Taegtmeyer, H. Alterations of the circadian clock in the heart by streptozotocin-induced diabetes. J. Mol. Cell. Cardiol. 2002, 34, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Bostwick, J.; Nguyen, D.; Cornélissen, G.; Halberg, F.; Hoogerwerf, W.A. Effects of acute and chronic STZ-induced diabetes on clock gene expression and feeding in the gastrointestinal tract. Mol. Cell. Biochem. 2010, 338, 203–213. [Google Scholar] [CrossRef]
- Herichová, I.; Zeman, M.; Stebelová, K.; Ravingerová, T. Effect of streptozotocin-induced diabetes on daily expression of per2 and dbp in the heart and liver and melatonin rhythm in the pineal gland of Wistar rat. Mol. Cell. Biochem. 2005, 270, 223–229. [Google Scholar] [CrossRef]
- Stebelová, K.; Herichová, I.; Zeman, M. Diabetes induces changes in melatonin concentrations in peripheral tissues of rat. Neuro Endocrinol. Lett. 2007, 28, 159–165. [Google Scholar]
- Lederbogen, F.; Hummel, J.; Fademrecht, C.; Krumm, B.; Kühner, C.; Deuschle, M.; Ladwig, K.H.; Meisinger, C.; Wichmann, H.E.; Lutz, H.; et al. Flattened circadian cortisol rhythm in type 2 diabetes. Exp. Clin. Endocrinol. Diabetes. 2011, 119, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Hygum, K.; Starup-Linde, J.; Harsløf, T.; Jørgensen, N.R.; Hartmann, B.; Holst, J.J.; Langdahl, B.L. The diurnal variation of bone formation is attenuated in adult patients with type 2 diabetes. Eur. J. Endocrinol. 2019, 181, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Dumpala, S.; Zele, A.J.; Feigl, B. Outer Retinal Structure and Function Deficits Contribute to Circadian Disruption in Patients With Type 2 Diabetes. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1870–1878. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, V.; Gandasi, N.R.; Sage, D.; Tengholm, A.; Barg, S.; Dibner, C. In pancreatic islets from type 2 diabetes patients, the dampened circadian oscillators lead to reduced insulin and glucagon exocytosis. Proc. Natl. Acad. Sci. USA. 2020, 117, 2484–2495. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Takamura, T.; Matsuzawa-Nagata, N.; Shima, K.R.; Eto, T.; Misu, H.; Shiramoto, M.; Tsuru, T.; Irie, S.; Fujimura, A.; et al. Clock gene expression in peripheral leucocytes of patients with type 2 diabetes. Diabetologia 2009, 52, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Tian, L.; Ding, Y.; Gao, Y.; Li, D.; Tang, Y. Correlation between inflammatory markers and impaired circadian clock gene expression in type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2019, 156, 107831. [Google Scholar] [CrossRef]
- Sinturel, F.; Makhlouf, A.M.; Meyer, P.; Tran, C.; Pataky, Z.; Golay, A.; Rey, G.; Howald, C.; Dermitzakis, E.T.; Pichard, C.; et al. Cellular circadian period length inversely correlates with HbA(1c) levels in individuals with type 2 diabetes. Diabetologia 2019, 62, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, J.A.; Olsson, A.H.; Nagorny, C.L.; Malmgren, S.; Dekker-Nitert, M.; Ling, C.; Mulder, H. Regulation of core clock genes in human islets. Metab. Clin. Exp. 2012, 61, 978–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schipper, S.B.J.; Van Veen, M.M.; Elders, P.J.M.; van Straten, A.; Van Der Werf, Y.D.; Knutson, K.L.; Rutters, F. Sleep disorders in people with type 2 diabetes and associated health outcomes: A review of the literature. Diabetologia 2021, 64, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and metabolic syndrome in circadian Clock mutant mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamia, K.A.; Storch, K.F.; Weitz, C.J. Physiological significance of a peripheral tissue circadian clock. Proc. Natl. Acad. Sci. USA 2008, 105, 15172–15177. [Google Scholar] [CrossRef] [Green Version]
- Jacobi, D.; Liu, S.; Burkewitz, K.; Kory, N.; Knudsen, N.H.; Alexander, R.K.; Unluturk, U.; Li, X.; Kong, X.; Hyde, A.L.; et al. Hepatic Bmal1 Regulates Rhythmic Mitochondrial Dynamics and Promotes Metabolic Fitness. Cell Metab. 2015, 22, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, M.S.; Li, R.; Liu, V.Y.; Fu, L.; Moore, D.D.; Ma, K.; Yechoor, V.K. Loss of Bmal1 leads to uncoupling and impaired glucose-stimulated insulin secretion in β-cells. Islets 2011, 3, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.E.; Liu, Y.; Dentin, R.; Pongsawakul, P.Y.; Liu, A.C.; Hirota, T.; Nusinow, D.A.; Sun, X.; Landais, S.; Kodama, Y.; et al. Cryptochrome mediates circadian regulation of cAMP signaling and hepatic gluconeogenesis. Nature Med. 2010, 16, 1152–1156. [Google Scholar] [CrossRef]
- Qian, J.; Scheer, F. Circadian System and Glucose Metabolism: Implications for Physiology and Disease. Trends Endocrinol. Metab. 2016, 27, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Bano-Otalora, B.; Martial, F.; Harding, C.; Bechtold, D.A.; Allen, A.E.; Brown, T.M.; Belle, M.D.C.; Lucas, R.J. Bright daytime light enhances circadian amplitude in a diurnal mammal. Proc. Natl. Acad. Sci. USA 2021, 118, e2100094118. [Google Scholar] [CrossRef]
- Hubbard, J.; Kobayashi Frisk, M.; Ruppert, E.; Tsai, J.W.; Fuchs, F.; Robin-Choteau, L.; Husse, J.; Calvel, L.; Eichele, G.; Franken, P.; et al. Dissecting and modeling photic and melanopsin effects to predict sleep disturbances induced by irregular light exposure in mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2017364118. [Google Scholar] [CrossRef]
- Nankivell, V.A.; Tan, J.T.M.; Wilsdon, L.A.; Morrison, K.R.; Bilu, C.; Psaltis, P.J.; Zimmet, P.; Kronfeld-Schor, N.; Nicholls, S.J.; Bursill, C.A.; et al. Circadian disruption by short light exposure and a high energy diet impairs glucose tolerance and increases cardiac fibrosis in Psammomys obesus. Sci. Rep. 2021, 11, 9673. [Google Scholar] [CrossRef]
- Coomans, C.P.; van den Berg, S.A.; Houben, T.; van Klinken, J.B.; van den Berg, R.; Pronk, A.C.; Havekes, L.M.; Romijn, J.A.; van Dijk, K.W.; Biermasz, N.R.; et al. Detrimental effects of constant light exposure and high-fat diet on circadian energy metabolism and insulin sensitivity. FASEB J. 2013, 27, 1721–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obayashi, K.; Yamagami, Y.; Kurumatani, N.; Saeki, K. Bedroom lighting environment and incident diabetes mellitus: A longitudinal study of the HEIJO-KYO cohort. Sleep Med. 2020, 65, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J.; Purvis, T.E.; Mistretta, J.; Scheer, F.A. Effects of the Internal Circadian System and Circadian Misalignment on Glucose Tolerance in Chronic Shift Workers. J. Clin. Endocrinol. Metab. 2016, 101, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.; Schernhammer, E.S.; Sun, Q.; Hu, F.B. Rotating night shift work and risk of type 2 diabetes: Two prospective cohort studies in women. PLoS Med. 2011, 8, e1001141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashti, H.S.; Scheer, F.; Saxena, R.; Garaulet, M. Timing of Food Intake: Identifying Contributing Factors to Design Effective Interventions. Adv. Nutr. 2019, 10, 606–620. [Google Scholar] [CrossRef]
- Stenvers, D.J.; Scheer, F.; Schrauwen, P.; la Fleur, S.E.; Kalsbeek, A. Circadian clocks and insulin resistance. Nat. Rev. Endocrinol. 2019, 15, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, D.; Carter, J.R.; Van Cauter, E.; Leproult, R. Adverse Impact of Sleep Restriction and Circadian Misalignment on Autonomic Function in Healthy Young Adults. Hypertension 2016, 68, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, P.; Yeh, W.H.; Dumont, G.A.; Boivin, D.B. A circadian rhythm in heart rate variability contributes to the increased cardiac sympathovagal response to awakening in the morning. Chronobiol. Int. 2012, 29, 757–768. [Google Scholar] [CrossRef]
- Stamatakis, K.A.; Punjabi, N.M. Effects of sleep fragmentation on glucose metabolism in normal subjects. Chest 2010, 137, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Andrews, R.C.; Walker, B.R. Glucocorticoids and insulin resistance: Old hormones, new targets. Clin. Sci. 1999, 96, 513–523. [Google Scholar] [CrossRef]
- Ambrée, O.; Touma, C.; Görtz, N.; Keyvani, K.; Paulus, W.; Palme, R.; Sachser, N. Activity changes and marked stereotypic behavior precede Abeta pathology in TgCRND8 Alzheimer mice. Neurobiol. Aging 2006, 27, 955–964. [Google Scholar] [CrossRef]
- Gorman, M.R.; Yellon, S. Lifespan daily locomotor activity rhythms in a mouse model of amyloid-induced neuropathology. Chronobiol. Int. 2010, 27, 1159–1177. [Google Scholar] [CrossRef]
- Wisor, J.P.; Edgar, D.M.; Yesavage, J.; Ryan, H.S.; McCormick, C.M.; Lapustea, N.; Murphy, G.M., Jr. Sleep and circadian abnormalities in a transgenic mouse model of Alzheimer’s disease: A role for cholinergic transmission. Neuroscience 2005, 131, 375–385. [Google Scholar] [CrossRef]
- Buhl, E.; Higham, J.P.; Hodge, J.J.L. Alzheimer’s disease-associated tau alters Drosophila circadian activity, sleep and clock neuron electrophysiology. Neurobiol. Dis. 2019, 130, 104507. [Google Scholar] [CrossRef] [PubMed]
- Sterniczuk, R.; Dyck, R.H.; Laferla, F.M.; Antle, M.C. Characterization of the 3 × Tg-AD mouse model of Alzheimer’s disease: Part 1. Circadian changes. Brain Res. 2010, 1348, 139–148. [Google Scholar] [CrossRef]
- Zhou, L.; Gao, Q.; Nie, M.; Gu, J.L.; Hao, W.; Wang, L.; Cao, J.M. Degeneration and energy shortage in the suprachiasmatic nucleus underlies the circadian rhythm disturbance in ApoE (-/-) mice: Implications for Alzheimer’s disease. Sci. Rep. 2016, 6, 36335. [Google Scholar] [CrossRef] [Green Version]
- Videnovic, A.; Lazar, A.S.; Barker, R.A.; Overeem, S. ‘The clocks that time us’—Circadian rhythms in neurodegenerative disorders. Nat. Rev. Neurol. 2014, 10, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, C.F.; Herbert, J.; van Someren, E.J.; Hodges, J.R.; Hastings, M.H. Disrupted daily activity/rest cycles in relation to daily cortisol rhythms of home-dwelling patients with early Alzheimer’s dementia. Brain A J. Neurol. 2004, 127, 1061–1074. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.H.; Fischer, D.F.; Kalsbeek, A.; Garidou-Boof, M.L.; van der Vliet, J.; van Heijningen, C.; Liu, R.Y.; Zhou, J.N.; Swaab, D.F. Pineal clock gene oscillation is disturbed in Alzheimer’s disease, due to functional disconnection from the “master clock”. FASEB J. 2006, 20, 1874–1876. [Google Scholar] [CrossRef] [Green Version]
- Bonanni, E.; Maestri, M.; Tognoni, G.; Fabbrini, M.; Nucciarone, B.; Manca, M.L.; Gori, S.; Iudice, A.; Murri, L. Daytime sleepiness in mild and moderate Alzheimer’s disease and its relationship with cognitive impairment. J. Sleep Res. 2005, 14, 311–317. [Google Scholar] [CrossRef]
- Lim, M.M.; Gerstner, J.R.; Holtzman, D.M. The sleep-wake cycle and Alzheimer’s disease: What do we know? Neurodegener. Dis. Manag. 2014, 4, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skene, D.J.; Swaab, D.F. Melatonin rhythmicity: Effect of age and Alzheimer’s disease. Exp. Gerontol. 2003, 38, 199–206. [Google Scholar] [CrossRef]
- Coogan, A.N.; Schutová, B.; Husung, S.; Furczyk, K.; Baune, B.T.; Kropp, P.; Häßler, F.; Thome, J. The circadian system in Alzheimer’s disease: Disturbances, mechanisms, and opportunities. Biol. Psychiatry 2013, 74, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Van Someren, E.J.; Shea, S.A.; Scheer, F.A. Reduction of scale invariance of activity fluctuations with aging and Alzheimer’s disease: Involvement of the circadian pacemaker. Proc. Natl. Acad. Sci. USA 2009, 106, 2490–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cermakian, N.; Lamont, E.W.; Boudreau, P.; Boivin, D.B. Circadian clock gene expression in brain regions of Alzheimer’s disease patients and control subjects. J. Biol. Rhythm. 2011, 26, 160–170. [Google Scholar] [CrossRef]
- Saeed, Y.; Abbott, S.M. Circadian Disruption Associated with Alzheimer’s Disease. Curr. Neurol. Neurosci. Rep. 2017, 17, 29. [Google Scholar] [CrossRef]
- Chauhan, R.; Chen, K.F.; Kent, B.A.; Crowther, D.C. Central and peripheral circadian clocks and their role in Alzheimer’s disease. Dis. Models Mech. 2017, 10, 1187–1199. [Google Scholar] [CrossRef] [Green Version]
- Karatsoreos, I.N.; Bhagat, S.; Bloss, E.B.; Morrison, J.H.; McEwen, B.S. Disruption of circadian clocks has ramifications for metabolism, brain, and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 1657–1662. [Google Scholar] [CrossRef] [Green Version]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef] [Green Version]
- Kress, G.J.; Liao, F.; Dimitry, J.; Cedeno, M.R.; FitzGerald, G.A.; Holtzman, D.M.; Musiek, E.S. Regulation of amyloid-β dynamics and pathology by the circadian clock. J. Exp. Med. 2018, 215, 1059–1068. [Google Scholar] [CrossRef]
- Lin, L.; Huang, Q.X.; Yang, S.S.; Chu, J.; Wang, J.Z.; Tian, Q. Melatonin in Alzheimer’s disease. Int. J. Mol. Sci. 2013, 14, 14575–14593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, J.; Wan, J.; Liu, A.; Sun, J. Melatonin regulates Aβ production/clearance balance and Aβ neurotoxicity: A potential therapeutic molecule for Alzheimer’s disease. Biomed. Pharmacother. 2020, 132, 110887. [Google Scholar] [CrossRef]
- Roh, J.H.; Jiang, H.; Finn, M.B.; Stewart, F.R.; Mahan, T.E.; Cirrito, J.R.; Heda, A.; Snider, B.J.; Li, M.; Yanagisawa, M.; et al. Potential role of orexin and sleep modulation in the pathogenesis of Alzheimer’s disease. J. Exp. Med. 2014, 211, 2487–2496. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.E.; Lim, M.M.; Bateman, R.J.; Lee, J.J.; Smyth, L.P.; Cirrito, J.R.; Fujiki, N.; Nishino, S.; Holtzman, D.M. Amyloid-beta dynamics are regulated by orexin and the sleep-wake cycle. Science 2009, 326, 1005–1007. [Google Scholar] [CrossRef] [Green Version]
- Holth, J.K.; Fritschi, S.K.; Wang, C.; Pedersen, N.P.; Cirrito, J.R.; Mahan, T.E.; Finn, M.B.; Manis, M.; Geerling, J.C.; Fuller, P.M.; et al. The sleep-wake cycle regulates brain interstitial fluid tau in mice and CSF tau in humans. Science 2019, 363, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Di Meco, A.; Joshi, Y.B.; Praticò, D. Sleep deprivation impairs memory, tau metabolism, and synaptic integrity of a mouse model of Alzheimer’s disease with plaques and tangles. Neurobiol. Aging 2014, 35, 1813–1820. [Google Scholar] [CrossRef]
- Rothman, S.M.; Herdener, N.; Frankola, K.A.; Mughal, M.R.; Mattson, M.P. Chronic mild sleep restriction accentuates contextual memory impairments, and accumulations of cortical Aβ and pTau in a mouse model of Alzheimer’s disease. Brain Res. 2013, 1529, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Peng, X.; Fan, R.; Dong, K.; Shi, X.; Zhang, S.; Yu, X.; Yang, Y. Disruption of Circadian Clocks Promotes Progression of Alzheimer’s Disease in Diabetic Mice. Mol. Neurobiol. 2021, 58, 4404–4412. [Google Scholar] [CrossRef]
- Tönnies, E.; Trushina, E. Oxidative Stress, Synaptic Dysfunction, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef] [Green Version]
- Uddin, M.S.; Sumsuzzman, D.M.; Jeandet, P.; Behl, T.; Rauf, A.; Amran, M.S.; Ashraf, G.M. Deciphering the Interacting Mechanisms of Circadian Disruption and Alzheimer’s Disease. Neurochem. Res. 2021, 46, 1603–1617. [Google Scholar] [CrossRef]
- Walker, W.H., 2nd; Walton, J.C.; DeVries, A.C.; Nelson, R.J. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, S.; Fuso, A.; Ippoliti, F.; Businaro, R. Stress-induced cytokines and neuronal dysfunction in Alzheimer’s disease. J. Alzheimer’s Dis. 2012, 28, 11–24. [Google Scholar] [CrossRef]
- Ennis, G.E.; An, Y.; Resnick, S.M.; Ferrucci, L.; O’Brien, R.J.; Moffat, S.D. Long-term cortisol measures predict Alzheimer disease risk. Neurology 2017, 88, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Lananna, B.V.; Musiek, E.S. The wrinkling of time: Aging, inflammation, oxidative stress, and the circadian clock in neurodegeneration. Neurobiol. Dis. 2020, 139, 104832. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Warabi, E.; Mann, G.E. Circadian control of BDNF-mediated Nrf2 activation in astrocytes protects dopaminergic neurons from ferroptosis. Free Radic. Biol. Med. 2019, 133, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Wright, K.P., Jr.; McHill, A.W.; Birks, B.R.; Griffin, B.R.; Rusterholz, T.; Chinoy, E.D. Entrainment of the human circadian clock to the natural light-dark cycle. Curr. Biol. 2013, 23, 1554–1558. [Google Scholar] [CrossRef] [Green Version]
- Golden, R.N.; Gaynes, B.N.; Ekstrom, R.D.; Hamer, R.M.; Jacobsen, F.M.; Suppes, T.; Wisner, K.L.; Nemeroff, C.B. The efficacy of light therapy in the treatment of mood disorders: A review and meta-analysis of the evidence. Am. J. Psychiatry 2005, 162, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Comtet, H.; Geoffroy, P.A.; Kobayashi Frisk, M.; Hubbard, J.; Robin-Choteau, L.; Calvel, L.; Hugueny, L.; Viola, A.U.; Bourgin, P. Light therapy with boxes or glasses to counteract effects of acute sleep deprivation. Sci. Rep. 2019, 9, 18073. [Google Scholar] [CrossRef] [Green Version]
- Bilu, C.; Einat, H.; Zimmet, P.; Vishnevskia-Dai, V.; Kronfeld-Schor, N. Beneficial effects of daytime high-intensity light exposure on daily rhythms, metabolic state and affect. Sci. Rep. 2020, 10, 19782. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, A.; van Raalte, D.H.; Nguyen, H.T.; Rutters, F.; van de Ven, P.M.; Elders, P.J.M.; Moll, A.C.; Van Someren, E.J.W.; Snoek, F.J.; Beekman, A.T.F.; et al. Effects of Light Therapy on Mood and Insulin Sensitivity in Patients With Type 2 Diabetes and Depression: Results From a Randomized Placebo-Controlled Trial. Diabetes Care 2019, 42, 529–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roccaro, I.; Smirni, D. Fiat Lux: The Light Became Therapy. An Overview on the Bright Light Therapy in Alzheimer’s Disease Sleep Disorders. J. Alzheimer’s Dis. 2020, 77, 113–125. [Google Scholar] [CrossRef]
- Asher, G.; Sassone-Corsi, P. Time for food: The intimate interplay between nutrition, metabolism, and the circadian clock. Cell 2015, 161, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Manoogian, E.N.C.; Panda, S. Circadian rhythms, time-restricted feeding, and healthy aging. Ageing Res. Rev. 2017, 39, 59–67. [Google Scholar] [CrossRef]
- Hawley, J.A.; Sassone-Corsi, P.; Zierath, J.R. Chrono-nutrition for the prevention and treatment of obesity and type 2 diabetes: From mice to men. Diabetologia 2020, 63, 2253–2259. [Google Scholar] [CrossRef]
- Gabel, K.; Kroeger, C.M.; Trepanowski, J.F.; Hoddy, K.K.; Cienfuegos, S.; Kalam, F.; Varady, K.A. Differential Effects of Alternate-Day Fasting Versus Daily Calorie Restriction on Insulin Resistance. Obesity 2019, 27, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Parr, E.B.; Devlin, B.L.; Radford, B.E.; Hawley, J.A. A Delayed Morning and Earlier Evening Time-Restricted Feeding Protocol for Improving Glycemic Control and Dietary Adherence in Men with Overweight/Obesity: A Randomized Controlled Trial. Nutrients 2020, 12, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makwana, K.; Gosai, N.; Poe, A.; Kondratov, R.V. Calorie restriction reprograms diurnal rhythms in protein translation to regulate metabolism. FASEB J. 2019, 33, 4473–4489. [Google Scholar] [CrossRef]
- Hu, D.; Xie, Z.; Ye, Y.; Bahijri, S.; Chen, M. The beneficial effects of intermittent fasting: An update on mechanism, and the role of circadian rhythm and gut microbiota. Hepatobiliary Surg. Nutr. 2020, 9, 597–602. [Google Scholar] [CrossRef]
- Yu, Q.; Zou, L.; Kong, Z.; Yang, L. Cognitive Impact of Calorie Restriction: A Narrative Review. J. Am. Med. Dir. Assoc. 2020, 21, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Cremonini, A.L.; Caffa, I.; Cea, M.; Nencioni, A.; Odetti, P.; Monacelli, F. Nutrients in the Prevention of Alzheimer’s Disease. Oxidative Med. Cell. Longev. 2019, 2019, 9874159. [Google Scholar] [CrossRef] [Green Version]
- Shin, B.K.; Kang, S.; Kim, D.S.; Park, S. Intermittent fasting protects against the deterioration of cognitive function, energy metabolism and dyslipidemia in Alzheimer’s disease-induced estrogen deficient rats. Exp. Biol. Med. 2018, 243, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Gudden, J.; Arias Vasquez, A.; Bloemendaal, M. The Effects of Intermittent Fasting on Brain and Cognitive Function. Nutrients 2021, 13, 3166. [Google Scholar] [CrossRef]
- Wang, H.B.; Loh, D.H.; Whittaker, D.S.; Cutler, T.; Howland, D.; Colwell, C.S. Time-Restricted Feeding Improves Circadian Dysfunction as well as Motor Symptoms in the Q175 Mouse Model of Huntington’s Disease. eNeuro 2018, 5, ENEURO.0431-17.2017. [Google Scholar] [CrossRef]
- Currenti, W.; Godos, J.; Castellano, S.; Caruso, G.; Ferri, R.; Caraci, F.; Grosso, G.; Galvano, F. Association between Time Restricted Feeding and Cognitive Status in Older Italian Adults. Nutrients 2021, 13, 191. [Google Scholar] [CrossRef] [PubMed]
- Tahara, Y.; Aoyama, S.; Shibata, S. The mammalian circadian clock and its entrainment by stress and exercise. J. Physiol. Sci. 2017, 67, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Marchant, E.G.; Mistlberger, R.E. Entrainment and phase shifting of circadian rhythms in mice by forced treadmill running. Physiol. Behav. 1996, 60, 657–663. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Hashimoto, S.; Masubuchi, S.; Natsubori, A.; Nishide, S.Y.; Honma, S.; Honma, K. Differential regulation of circadian melatonin rhythm and sleep-wake cycle by bright lights and nonphotic time cues in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R546–R557. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Hashimoto, S.; Tanahashi, Y.; Nishide, S.Y.; Honma, S.; Honma, K. Physical exercise accelerates reentrainment of human sleep-wake cycle but not of plasma melatonin rhythm to 8-h phase-advanced sleep schedule. Am. J. Physiology. Regul. Integr. Comp. Physiol. 2010, 298, R681–R691. [Google Scholar] [CrossRef]
- Boulé, N.G.; Haddad, E.; Kenny, G.P.; Wells, G.A.; Sigal, R.J. Effects of exercise on glycemic control and body mass in type 2 diabetes mellitus: A meta-analysis of controlled clinical trials. JAMA 2001, 286, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Colberg, S.R.; Sigal, R.J.; Yardley, J.E.; Riddell, M.C.; Dunstan, D.W.; Dempsey, P.C.; Horton, E.S.; Castorino, K.; Tate, D.F. Physical Activity/Exercise and Diabetes: A Position Statement of the American Diabetes Association. Diabetes Care 2016, 39, 2065–2079. [Google Scholar] [CrossRef] [Green Version]
- Erickson, M.L.; Zhang, H.; Mey, J.T.; Kirwan, J.P. Exercise Training Impacts Skeletal Muscle Clock Machinery in Prediabetes. Med. Sci. Sports Exerc. 2020, 52, 2078–2085. [Google Scholar] [CrossRef]
- De la Rosa, A.; Olaso-Gonzalez, G.; Arc-Chagnaud, C.; Millan, F.; Salvador-Pascual, A.; García-Lucerga, C.; Blasco-Lafarga, C.; Garcia-Dominguez, E.; Carretero, A.; Correas, A.G.; et al. Physical exercise in the prevention and treatment of Alzheimer’s disease. J. Sport Health Sci. 2020, 9, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Tortosa-Martínez, J.; Manchado, C.; Cortell-Tormo, J.M.; Chulvi-Medrano, I. Exercise, the diurnal cycle of cortisol and cognitive impairment in older adults. Neurobiol. Stress 2018, 9, 40–47. [Google Scholar] [CrossRef]
- Dijckmans, B.; Tortosa-Martínez, J.; Caus, N.; González-Caballero, G.; Martínez-Pelegrin, B.; Manchado-Lopez, C.; Cortell-Tormo, J.M.; Chulvi-Medrano, I.; Clow, A. Does the diurnal cycle of cortisol explain the relationship between physical performance and cognitive function in older adults? Eur. Rev. Aging Phys. Act. 2017, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Cuesta, M.; Aungier, J.; Morton, A.J. Behavioral therapy reverses circadian deficits in a transgenic mouse model of Huntington’s disease. Neurobiol. Dis. 2014, 63, 85–91. [Google Scholar] [CrossRef]
- Ahasan, R.; Lewko, J.; Campbell, D.; Salmoni, A. Adaptation to night shifts and synchronisation processes of night workers. J. Physiol. Anthropol. Appl. Hum. Sci. 2001, 20, 215–226. [Google Scholar] [CrossRef]
- Chinoy, E.D.; Harris, M.P.; Kim, M.J.; Wang, W.; Duffy, J.F. Scheduled evening sleep and enhanced lighting improve adaptation to night shift work in older adults. Occup. Environ. Med. 2016, 73, 869–876. [Google Scholar] [CrossRef]
- Vetter, C.; Fischer, D.; Matera, J.L.; Roenneberg, T. Aligning work and circadian time in shift workers improves sleep and reduces circadian disruption. Curr. Biol. 2015, 25, 907–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garde, A.H.; Begtrup, L.; Bjorvatn, B.; Bonde, J.P.; Hansen, J.; Hansen Å, M.; Härmä, M.; Jensen, M.A.; Kecklund, G.; Kolstad, H.A.; et al. How to schedule night shift work in order to reduce health and safety risks. Scand. J. Work. Environ. Health 2020, 46, 557–569. [Google Scholar] [CrossRef]
- Gooley, J.J.; Chamberlain, K.; Smith, K.A.; Khalsa, S.B.; Rajaratnam, S.M.; Van Reen, E.; Zeitzer, J.M.; Czeisler, C.A.; Lockley, S.W. Exposure to room light before bedtime suppresses melatonin onset and shortens melatonin duration in humans. J. Clin. Endocrinol. Metab. 2011, 96, E463–E472. [Google Scholar] [CrossRef] [Green Version]
- Stevens, R.G.; Zhu, Y. Electric light, particularly at night, disrupts human circadian rhythmicity: Is that a problem? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razavi, P.; Devore, E.E.; Bajaj, A.; Lockley, S.W.; Figueiro, M.G.; Ricchiuti, V.; Gauderman, W.J.; Hankinson, S.E.; Willett, W.C.; Schernhammer, E.S. Shift Work, Chronotype, and Melatonin Rhythm in Nurses. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1177–1186. [Google Scholar] [CrossRef]
- Forrestel, A.C.; Miedlich, S.U.; Yurcheshen, M.; Wittlin, S.D.; Sellix, M.T. Chronomedicine and type 2 diabetes: Shining some light on melatonin. Diabetologia 2017, 60, 808–822. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.J.; Aeschbach, D.; Scheer, F.A. Circadian system, sleep and endocrinology. Mol. Cell. Endocrinol. 2012, 349, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.F.; Uddin, M.S.; Uddin, G.M.S.; Sumsuzzman, D.M.; Islam, M.S.; Barreto, G.E.; Mathew, B.; Ashraf, G.M. Melatonin in Alzheimer’s Disease: A Latent Endogenous Regulator of Neurogenesis to Mitigate Alzheimer’s Neuropathology. Mol. Neurobiol. 2019, 56, 8255–8276. [Google Scholar] [CrossRef] [PubMed]
- Vasey, C.; McBride, J.; Penta, K. Circadian Rhythm Dysregulation and Restoration: The Role of Melatonin. Nutrients 2021, 13, 3480. [Google Scholar] [CrossRef]
- Garfinkel, D.; Zorin, M.; Wainstein, J.; Matas, Z.; Laudon, M.; Zisapel, N. Efficacy and safety of prolonged-release melatonin in insomnia patients with diabetes: A randomized, double-blind, crossover study. Diabetes Metab. Syndr. Obes. Targets Ther. 2011, 4, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Tsunoda, T.; Yamada, M.; Akiyama, T.; Minami, T.; Yoshii, T.; Kondo, Y.; Satoh, S.; Terauchi, Y. The Effects of Ramelteon on Glucose Metabolism and Sleep Quality in Type 2 Diabetic Patients With Insomnia: A Pilot Prospective Randomized Controlled Trial. J. Clin. Med. Res. 2016, 8, 878–887. [Google Scholar] [CrossRef] [Green Version]
- Wade, A.G.; Farmer, M.; Harari, G.; Fund, N.; Laudon, M.; Nir, T.; Frydman-Marom, A.; Zisapel, N. Add-on prolonged-release melatonin for cognitive function and sleep in mild to moderate Alzheimer’s disease: A 6-month, randomized, placebo-controlled, multicenter trial. Clin. Interv. Aging 2014, 9, 947–961. [Google Scholar] [CrossRef]
- Asayama, K.; Yamadera, H.; Ito, T.; Suzuki, H.; Kudo, Y.; Endo, S. Double blind study of melatonin effects on the sleep-wake rhythm, cognitive and non-cognitive functions in Alzheimer type dementia. J. Nippon. Med. Sch. 2003, 70, 334–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Yang, M.; Wang, Y.; Ren, J.; Lin, P.; Cui, C.; Song, J.; He, Q.; Hu, H.; Wang, K.; et al. Melatonin prevents diabetes-associated cognitive dysfunction from microglia-mediated neuroinflammation by activating autophagy via TLR4/Akt/mTOR pathway. FASEB J. 2021, 35, e21485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lu, J.; Zhu, H.; Zhou, X.; Wei, X.; Gu, M. Association of Serum Melatonin Level with Mild Cognitive Impairment in Type 2 Diabetic Patients: A Cross-Sectional Study. Int. J. Endocrinol. 2021, 2021, 5566019. [Google Scholar] [CrossRef]
- Tsujino, N.; Sakurai, T. Orexin/hypocretin: A neuropeptide at the interface of sleep, energy homeostasis, and reward system. Pharmacol. Rev. 2009, 61, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Saper, C.B.; Scammell, T.E.; Lu, J. Hypothalamic regulation of sleep and circadian rhythms. Nature 2005, 437, 1257–1263. [Google Scholar] [CrossRef]
- Greco, C.M.; Sassone-Corsi, P. Circadian blueprint of metabolic pathways in the brain. Nat. Rev. Neurosci. 2019, 20, 71–82. [Google Scholar] [CrossRef]
- Kon, K.; Tsuneki, H.; Ito, H.; Takemura, Y.; Sato, K.; Yamazaki, M.; Ishii, Y.; Sasahara, M.; Rudich, A.; Maeda, T.; et al. Chronotherapeutic effect of orexin antagonists on glucose metabolism in diabetic mice. J. Endocrinol. 2019, 243, 59–72. [Google Scholar] [CrossRef]
- Tsuneki, H.; Kon, K.; Ito, H.; Yamazaki, M.; Takahara, S.; Toyooka, N.; Ishii, Y.; Sasahara, M.; Wada, T.; Yanagisawa, M.; et al. Timed Inhibition of Orexin System by Suvorexant Improved Sleep and Glucose Metabolism in Type 2 Diabetic db/db Mice. Endocrinology 2016, 157, 4146–4157. [Google Scholar] [CrossRef]
- Yoshikawa, F.; Shigiyama, F.; Ando, Y.; Miyagi, M.; Uchino, H.; Hirose, T.; Kumashiro, N. Chronotherapeutic efficacy of suvorexant on sleep quality and metabolic parameters in patients with type 2 diabetes and insomnia. Diabetes Res. Clin. Pract. 2020, 169, 108412. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.; Beuckmann, C.T.; Willie, J.T.; Hara, J.; Tsujino, N.; Mieda, M.; Tominaga, M.; Yagami, K.; Sugiyama, F.; Goto, K.; et al. Hypothalamic orexin neurons regulate arousal according to energy balance in mice. Neuron 2003, 38, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Ueta, Y.; Date, Y.; Nakazato, M.; Hara, Y.; Serino, R.; Nomura, M.; Shibuya, I.; Matsukura, S.; Yamashita, H. Down regulation of the prepro-orexin gene expression in genetically obese mice. Mol. Brain Res. 1999, 65, 14–22. [Google Scholar] [CrossRef]
- Tsuneki, H.; Tokai, E.; Nakamura, Y.; Takahashi, K.; Fujita, M.; Asaoka, T.; Kon, K.; Anzawa, Y.; Wada, T.; Takasaki, I.; et al. Hypothalamic orexin prevents hepatic insulin resistance via daily bidirectional regulation of autonomic nervous system in mice. Diabetes 2015, 64, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Zarifkar, M.; Noshad, S.; Shahriari, M.; Afarideh, M.; Khajeh, E.; Karimi, Z.; Ghajar, A.; Esteghamati, A. Inverse Association of Peripheral Orexin-A with Insulin Resistance in Type 2 Diabetes Mellitus: A Randomized Clinical Trial. Rev. Diabet. Stud. 2017, 14, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S., 2nd; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA. 1998, 95, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Jiang, W.; Zhang, E.E. Orexin signaling regulates both the hippocampal clock and the circadian oscillation of Alzheimer’s disease-risk genes. Sci. Rep. 2016, 6, 36035. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Meng, Y.; Chu, B.; Shen, Y.; Xue, X.; Song, C.; Liu, X.; Ding, M.; Cao, X.; Wang, P.; et al. Orexin-A exacerbates Alzheimer’s disease by inducing mitochondrial impairment. Neurosci. Lett. 2020, 718, 134741. [Google Scholar] [CrossRef] [PubMed]
- Beuckmann, C.T.; Suzuki, H.; Musiek, E.S.; Ueno, T.; Sato, T.; Bando, M.; Osada, Y.; Moline, M. Evaluation of SAMP8 Mice as a Model for Sleep-Wake and Rhythm Disturbances Associated with Alzheimer’s Disease: Impact of Treatment with the Dual Orexin (Hypocretin) Receptor Antagonist Lemborexant. J. Alzheimer’s Dis. 2021, 81, 1151–1167. [Google Scholar] [CrossRef]
- Herring, W.J.; Ceesay, P.; Snyder, E.; Bliwise, D.; Budd, K.; Hutzelmann, J.; Stevens, J.; Lines, C.; Michelson, D. Polysomnographic assessment of suvorexant in patients with probable Alzheimer’s disease dementia and insomnia: A randomized trial. Alzheimer’s Dement. 2020, 16, 541–551. [Google Scholar] [CrossRef]
- Tsuneki, H.; Wada, T.; Sasaoka, T. Chronopathophysiological implications of orexin in sleep disturbances and lifestyle-related disorders. Pharmacol. Ther. 2018, 186, 25–44. [Google Scholar] [CrossRef]
- Duez, H.; Staels, B. The nuclear receptors Rev-erbs and RORs integrate circadian rhythms and metabolism. Diabetes Vasc. Dis. Res. 2008, 5, 82–88. [Google Scholar] [CrossRef]
- He, B.; Nohara, K.; Park, N.; Park, Y.S.; Guillory, B.; Zhao, Z.; Garcia, J.M.; Koike, N.; Lee, C.C.; Takahashi, J.S.; et al. The Small Molecule Nobiletin Targets the Molecular Oscillator to Enhance Circadian Rhythms and Protect against Metabolic Syndrome. Cell Metab. 2016, 23, 610–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roby, D.A.; Ruiz, F.; Kermath, B.A.; Voorhees, J.R.; Niehoff, M.; Zhang, J.; Morley, J.E.; Musiek, E.S.; Farr, S.A.; Burris, T.P. Pharmacological activation of the nuclear receptor REV-ERB reverses cognitive deficits and reduces amyloid-β burden in a mouse model of Alzheimer’s disease. PloS ONE 2019, 14, e0215004. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.K.; Zhu, Y.; Sun, H.Y.; Xu, X.Y.; Zhang, S.; Hao, Z.B.; Wang, G.H.; Mu, C.C.; Ren, H.G. Pharmacological activation of REV-ERBα represses LPS-induced microglial activation through the NF-κB pathway. Acta Pharmacol. Sin. 2019, 40, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Nohara, K.; Wirianto, M.; Escobedo, G., Jr.; Lim, J.Y.; Morales, R.; Yoo, S.H.; Chen, Z. Effects of the Clock Modulator Nobiletin on Circadian Rhythms and Pathophysiology in Female Mice of an Alzheimer’s Disease Model. Biomolecules 2021, 11, 1004. [Google Scholar] [CrossRef]
- Lamos, E.M.; Levitt, D.L.; Munir, K.M. A review of dopamine agonist therapy in type 2 diabetes and effects on cardio-metabolic parameters. Prim. Care Diabetes 2016, 10, 60–65. [Google Scholar] [CrossRef]
- Holt, R.I.; Barnett, A.H.; Bailey, C.J. Bromocriptine: Old drug, new formulation and new indication. Diabetes Obes. Metab. 2010, 12, 1048–1057. [Google Scholar] [CrossRef]
- Cincotta, A.H.; Meier, A.H.; Cincotta, M., Jr. Bromocriptine improves glycaemic control and serum lipid profile in obese Type 2 diabetic subjects: A new approach in the treatment of diabetes. Expert Opin. Investig. Drugs 1999, 8, 1683–1707. [Google Scholar] [CrossRef]
- Meguro, K.; Meguro, M.; Tanaka, Y.; Akanuma, K.; Yamaguchi, K.; Itoh, M. Risperidone is effective for wandering and disturbed sleep/wake patterns in Alzheimer’s disease. J. Geriatr. Psychiatry Neurol. 2004, 17, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.Y.; Xia, Q.P.; Hu, Y.H.; Wang, C.; He, L. Dopamine D1 receptor agonist A-68930 ameliorates Aβ(1-42)-induced cognitive impairment and neuroinflammation in mice. Int. Immunopharmacol. 2020, 88, 106963. [Google Scholar] [CrossRef] [PubMed]
- Grigolon, R.B.; Brietzke, E.; Mansur, R.B.; Idzikowski, M.A.; Gerchman, F.; De Felice, F.G.; McIntyre, R.S. Association between diabetes and mood disorders and the potential use of anti-hyperglycemic agents as antidepressants. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2019, 95, 109720. [Google Scholar] [CrossRef]
- Sharma, A.; Mohammad, A.; Saini, A.K.; Goyal, R. Neuroprotective Effects of Fluoxetine on Molecular Markers of Circadian Rhythm, Cognitive Deficits, Oxidative Damage, and Biomarkers of Alzheimer’s Disease-Like Pathology Induced under Chronic Constant Light Regime in Wistar Rats. ACS Chem. Neurosci. 2021, 12, 2233–2246. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, K.; Tchekalarova, J.; Atanasova, D.; Kortenska, L.; Atanasova, M. Antidepressant agomelatine attenuates behavioral deficits and concomitant pathology observed in streptozotocin-induced model of Alzheimer’s disease in male rats. Horm. Behav. 2019, 107, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Y.; Bao, H.; Zhang, M.; Gao, F.; Kang, D.; Shen, Y. A New Neurovascular Panel Discriminates Between Patients with Type 2 Diabetes Mellitus with Cognitive Impairment and Cognitive Impairment Alone. J. Alzheimer’s Dis. 2020, 75, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Cicognola, C.; Chiasserini, D.; Eusebi, P.; Andreasson, U.; Vanderstichele, H.; Zetterberg, H.; Parnetti, L.; Blennow, K. No diurnal variation of classical and candidate biomarkers of Alzheimer’s disease in CSF. Mol. Neurodegener. 2016, 11, 65. [Google Scholar] [CrossRef] [Green Version]
- Lananna, B.V.; McKee, C.A.; King, M.W.; Del-Aguila, J.L.; Dimitry, J.M.; Farias, F.H.G.; Nadarajah, C.J.; Xiong, D.D.; Guo, C.; Cammack, A.J.; et al. Chi3l1/YKL-40 is controlled by the astrocyte circadian clock and regulates neuroinflammation and Alzheimer’s disease pathogenesis. Sci. Transl. Med. 2020, 12, eaax3519. [Google Scholar] [CrossRef]
- Salas, I.H.; De Strooper, B. Diabetes and Alzheimer’s Disease: A Link not as Simple as it Seems. Neurochem. Res. 2019, 44, 1271–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, S.M.; Malkani, R.G.; Zee, P.C. Circadian disruption and human health: A bidirectional relationship. Eur. J. Neurosci. 2020, 51, 567–583. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| First Author, Year | Animals | Age | Type of Circadian Markers | Results |
|---|---|---|---|---|
| Hou, 2019 | Db/db, Db/+ mice | 16–24 w | Daily rhythms of BP, baroreflex sensitivity and mPer2Luc oscillations | Db/db mice had disrupted daily rhythms of BP, baroreflex sensitivity, and advanced phase shift of mPer2 daily oscillation in the liver, kidney, and submandibular gland. |
| Grosbellet, 2016 | Db/db, Db/+ mice | 10 w | Body temperature rhythm, general activity rhythm | Db/db mice had lower amplitude in body temperature rhythm and general activity rhythm under normal LD cycle and had a longer endogenous period for both activity and temperature rhythms compared with db/+ mice under constant darkness. |
| Su, 2012 | Db/db, Db/+ mice | 9–10 w | Daily rhythms of mRNA levels of multiple clock genes (Clock, Bmal1, Per1/2, Cry1/2, Rev-Erba) and target genes (Dbp and Pparγ) in the aorta, mesenteric arteries, heart, kidney, and SCN | Db/db mice had suppressed 24-h mRNA rhythms of the following clock and target genes: Per1/2, Cry1/2, their target genes, Dbp and Pparγ, in the aorta and mesenteric arteries; Dbp in the heart; Per1, Rev-Erba, and Dbp in the kidney; Per1 in the SCN. |
| Caton, 2011 | Db/db, Db/+ mice | 8 w | Expression levels of Clock, Bmal1, Per2, Cry1 in mRNA and protein in WAT | Db/db mice had lower expression of Clock mRNA (42%), CLOCK protein (72%), BMAL1 protein (34%; but not mRNA), Per2 mRNA (17%) in WAT, while no difference was found in Cry1 in WAT, compared with db/+ mice. |
| Senador, 2009 | Db/db, Db/+ mice | 7–8 w | Circadian rhythms of systolic arterial pressure variance and its low-frequency component | Circadian rhythms of systolic arterial pressure variance and its low-frequency component are absent in db/db mice |
| Su, 2008 | Db/db, Db/+ mice | 15–17 w | Oscillations of clock genes DBP and Bmal1 in vasculature; circadian rhythms of BP, heart rate, and locomotor circadian rhythm | Db/db mice had a disrupted BP, heart rate, and locomotor circadian rhythm associated with dampened oscillations of clock genes Dbp and Bmal1 mRNA in vasculature. |
| Laposky, 2008 | Db/db, wt/wt mice | 14–18 w | Diurnal rhythmicity of sleep-wake patterns | Db/db mice had increased total sleep time, sleep fragmentation and attenuated the daily rhythm of the sleep-wake cycle. |
| Kudo, 2004 | Db/db, Db/+ mice | 13–14 w | Daily oscillations of Per2 and Bmal1 mRNA expression in liver | The expression of Per2 mRNA levels was severely diminished and the phase of Bmal1 mRNA oscillation was advanced in the db/db mouse liver. |
| Woodie, 2020 | C5Bl/6N | 7 w; HFD 16 w | Daily rhythms of mRNA expressions of clock genes (Clock, Bmal1, Per2, Cry1) in the liver, hypothalamus and hippocampus | HFD caused disruptive mRNA expression rhythms of Bmal1 in the liver, Cry1 in the hypothalamus and all core clock genes (Clock, Bmal1, Per2, Cry1) in the hippocampus along with abnormal rhythms of AD-associated genes. |
| Katrina, 2015 | C57BL/6J | 8 w; HFD 5 w | Eating behavior and locomotor activity rhythms; PER2:LUC bioluminescence rhythms in liver | HFD disrupted eating behavior and locomotor activity rhythms; The phase of Per2 was advanced by 4 h in the liver. |
| Pendergast, 2013 | C57BL/6J | 7 w; HFD 1 w | PER2 expression in the gonadal white adipose tissue (surrounding the gonads), liver, lung, spleen, aorta, pituitary, SCN and arcuate complex | the phase of the PER2 rhythm was markedly advanced (by 5 h) in the liver of HFD mice, whereas rhythms in other tissues were not affected. |
| Hatori, 2012 | C57BL/6J | 12 w; HFD 6 w | Diurnal rhythms in food intake and RER; the oscillations of circadian clock genes (Per1, Per2, Cry1, Bmal1, Clock, Rorα Rev-erbα, Dbp) mRNA levels | HFD damped diurnal rhythms in food intake and RER in mice; HFD also dampened the oscillations of circadian clock components (Per1, Per2, Cry1, Bmal1, Clock, Rorα Rev-erbα and Dbp) in the liver. |
| Caton, 2011 | C56Bl/6 | 8 w; HFD 16 w | Expression levels of clock, bmal1, per2, CRY1 in mRNA and protein in WAT | Clock mRNA (60%) and protein levels (42%) were decreased in WAT of HFD mice compared to control. |
| Hsieh, 2010 | C57BL/6 | HFD 11 m | The mRNA expressions of circadian-clock genes and clock-controlled genes, including Per1-3, Cry1-2, Bmal1, Dbp, E4BP4, CK1varepsilon, PEPCK, PDK4 and NHE3 in the liver and kidneys | HFD disrupted the circadian rhythms of Per1-3, Cry1-2, Bmal1, Dbp, E4BP4, CK1varepsilon, PEPCK, PDK4 and NHE3 in the liver and kidneys. |
| Kohsaka, 2007 | C57BL/6J | 6 w; HFD 6 w | Free-running period, feeding behavior rhythms, Clock, Bmal1 and Per2 m RNA expression in the fat and liver. | HFD lengthened the free-running period in mice and attenuated the diurnal pattern of feeding behavior. The amplitudes of Clock, Bmal1 and Per2 mRNA expression were decreased in both the fat and liver of the HFD mice. |
| Yang, 2013 | C57BL/6 | Postnatalday 2 with a single injection of STZ | Circadian clock genes mRNA levels in the livers of mice at the age of 16 weeks | Only Bmal1, Cry1 and Per2 mRNA expressions were elevated for the group injected with STZ on the postnatal day 2. |
| Bostwick, 2010 | C57BL/6J | 10–12 w; STZ injection once a day for 4 days | The mRNA expressions of Per genes in the stomach body, proximal and distal colon, liver, kidney and lung | Per2 and Per3 mRNA expression levels of STZ-injected mice were generally phase-delayed within the gastrointestinal tract but not within the kidney or lung in acute (1 week) and chronic (12 weeks) STZ-induced diabetes compared with control mice, although the rhythmicity in expression of Per2 and Per3 persisted in all organs. |
| Stebelová, 2007 | Wistar rats | One single injection of STZ | Daily rhythm of melatonin concentrations in the pineal gland, plasma, pancreas, kidney, spleen and duodenum | The diabetic group resulted in lower melatonin levels in the pineal gland, pancreas, kidney and duodenum as compared to control, while no difference was found in the spleen on day 17 after STZ treatment. |
| Herichová, 2005 | Wistar rats | 8 w; STZ injection once | The mRNA expression of Per2 and Dbp in the heart and liver. | The rhythm in per2 and dbp expression was slightly advanced in hearts, although the rhythms existed; the daily rhythm of Per2 was lost and Dbp showed a similar advanced shift in the liver of mice 4 weeks after the STZ injection. |
| Kuriyama, 2004 | ddY mice | 15 w with a single injection of STZ | Rhythmic expression of Per2 mRNA and protein in the SCN, cerebral cortex and liver | mRNA and protein expression levels of Per2 were dampened in the liver but not SCN and cerebral cortex of mice on the fifth day after injection with STZ. |
| Young, 2002 | Wistar rats | One single injection of STZ | Daily rhythms of mRNA expressions of clock genes (Bmal1, Clock, Cry, Per), as well as three clock output genes (Dbp, Hlf and Tef) in the hearts | A phase shift (approximately 3 h early) was observed for the mRNA expression levels of Bmal1, Per, Cry and the three output genes (Dbp, Hlf and Tef) in the hearts of mice in 4 weeks after the initial STZ injection. |
| First Author, Year | Participants | Type of Circadian Markers | Results |
|---|---|---|---|
| Ushijima, 2020 | 13 non-T2DM and 12 T2DM with gastric cancer | Clock associated gene Dbp and PPAR-γ mRNA expression in omental adipose tissue | DBP and PPAR-γ mRNA expression are reduced in omental adipose tissue in T2DM patients. |
| Petrenko, 2020 | 12 T2DM patients and 27 nondiabetic patients | Clock genes (Clock, Bmal1, Per, Cry, Rev-erbα, Dpp) mRNA expression levels in human islets from T2DM and nondiabetic donors | mRNA expression levels of Per1-3, Cry2, Rev-erbα, Clock and Dbp were significantly diminished in T2D compared to nondiabetic islet cells combined with, while BMAL1 and CRY1 did not change. |
| Ando, 2020 | Study 1: 8 T2DM patients and 6 comparatively young non-diabetic volunteers Study 2: 12 male T2DM patients and 14 age-matched men | Clock genes (Clock, Bmal1, Per1, Per2, Per3 and Cry1) mRNA expression levels at 9 a.m., 3 p.m., 9 p.m., and 3 a.m. (study 1) and at 9 a.m. (study 2) in peripheral leucocytes | In study 1, mRNA expression levels of Bmal1, Per1, Per2 and Per3 were significantly lower in T2DM patients than in non-diabetic individuals at one or more time points. In study 2, lower transcript levels of Bmal1, Per1 and Per3 were found in leucocytes obtained from T2DM patients than in control individuals, and the transcript expression was inversely correlated with HbA(1c) levels. |
| Yu, 2019 | 36 T2DM patients and 14 non-diabetic volunteers | Transcript levels of circadian clock genes (Clock, Bmal1, Per1, Cry1 and Cry2) in peripheral blood leucocytes | The T2DM patients had lower CLOCK, BMAL1, PER1, CRY1 and CRY2 mRNA levels than nondiabetic participants in peripheral blood leucocytes. Blood inflammatory markers (IL-6, TNF-α) HbA1c levels were negatively correlated with Bmal1, Per1 and Cry1 mRNA levels. |
| Hygum, 2019 | 5 T1DM, 5 T2DM patients and 5 controls (age > 50 years) | 24-h variation of bone formation | The rhythms of bone formation marker procollagen type 1 N-terminal propeptide were lower in T2DM compared with controls. |
| Sinturel, 2019 | 9 obese and 8 non-obese individuals with T2DM and 11 non-diabetic controls | Rhythms of clock gene Bmal1 in dermal fibroblasts established from skin biopsies | The oscillation period of the Bmal1-luc reporter was significantly shorter in the type 2 diabetes group (particularly the obese subgroup) than controls. HbA1c values were found to be significantly inverse (ρ = −0.592; p < 0.05) with the circadian period length within cells from the T2DM group |
| Dumpala, 2019 | 23 patients with T2DM and 24 age-matched healthy controls | DLMO; sleep questionnaires; light exposure measured by actigraphy | T2DM had earlier DLMO (1 h), higher subjective sleep score than controls although no significant difference was found in light exposure pattern. |
| Perciaccante, 2016 | 90 Caucasian IR subjects (divided into four groups: IR with normal OGTT results, IR with IFT, IR with IGT and T2DM) and 25 control subjects | Autonomic nervous activity measured by 24-h ECG recording and heart rate variability | The IR groups all showed impaired autonomic activity reflected by sympathovagal balance (expressed by the LF/HF ratio) and reduced standard deviation of all sinus rhythm RR values compared to the controls. |
| Stamenkovic, 2012 | 5 T2DM patients and 55 nondiabetic controls | The core clock genes (Clock, Bmal1, Per1 to 3, Cry1 and Cry2) in islets from donors | The mRNA levels of Per2, Per3 and Cry2 were significantly lower in islets from donors with T2DM than the non-diabetic controls. mRNA levels of Per2, Per3, and Cry2 correlated positively with insulin content, and the expression of Per3 and Cry2 correlated negatively with glycated hemoglobin levels. |
| Lederbogen, 2011 | 63 ambulatory individuals with T2DM and 916 non-diabetic control subjects | Saliva cortisol concentrations on waking, a salivette ½ h, 8 h and 14 h after waking | Diabetic subjects had a flattened circadian cortisol profile, with lower levels in the morning and higher levels in the afternoon and evening. |
| First Author, Year | Study Design | Participant | Treatment | Type of Circadian Markers | Results |
|---|---|---|---|---|---|
| Brouwer, 2015 | RCT | 83 adult T2DM patients with major depressive episodes (mean age = 60.1/62.9 years) | Light therapy (10,000 lux) for 30 min every morning for 4 weeks at home | Sleep (duration, efficiency, time) | Light therapy did not result in significant changes in sleep duration, sleep efficiency, or mid-sleep time. Light therapy did not affect depressive symptoms in participants with higher insulin sensitivity, but it did produce an anti-depression effect in participants with lower insulin sensitivity. |
| Yamadera, 2000 | RCT | 27 adults with AD (mean age = 79.9 years) | Light therapy (3000 Lux; 9–11 a.m.) for 4 weeks | Sleep/nap time, awakenings in the night | The therapy improved circadian rhythm disturbances and MMSE scores, especially in the early stages of AD, although the CDR scores were not improved. |
| Gabel, 2019 | RCT | 43 insulin-resistant subjects (mean age = 44 years) | IF (25% of energy needs on “fast days”; 125% of needs on alternating “feast days”) or CR (75% of energy needs every day) for 12-month | None | IF and CR caused similar decreases in body weight compared with the control group. IF contributed to greater reductions in fasting insulin and insulin resistance than CR. |
| Parr, 2020 | RCT | 11 sedentary males (mean age = 38 years; mean BMI = 32.2 kg/m2) | Two isoenergetic diet protocols for 5 days, consuming meals at 10 a.m., 1 p.m. and 5 p.m. (TRF) or 7 a.m., 2 p.m., and 9 p.m. (EXF). | None | Total 24-h area under the curve of glucose tended to be lower for TRF versus EXF (−5.5 ± 9.0 mmol/L/h, P = 0.09). Area under the curve of nocturnal glucose was lower in TRF (−4.2 ± 5.8 mmol/L/h, P = 0.04). |
| Currenti, 2021 | observational study | 883 adults (age ≥ 50 years). Participants with an eating time window of less than 10 h over the last 6 months were identified as the TRF group. | None | None | Individuals adhering to TRF were less likely to have a cognitive impairment, compared to those with no eating time restrictions (OR = 0.28; 95% CI: 0.07–0.90). |
| Erickson, 2020 | RCT | 24 adults with obesity and prediabetes (mean age = 66 mean BMI = 34 kg/m2 mean fasting plasma glucose = 105 mg/dL) | Exercise intervention for 12 weeks (5 days per week at ~85% of heart rate max on a treadmill for 60 min) | Expression of circadian clock genes (BMAL1, CLOCK, CRY1/2, and PER 1/2) in skeletal muscle | BMI, peripheral insulin sensitivity and exercise capacity all improved (P < 0.005) with exercise training. Skeletal muscle BMAL1 gene (fold change, 1.62 ± 1.01; P = 0.027) and PER2 protein expression (fold change, 1.35 ± 0.05; P = 0.02) increased, whereas CLOCK, CRY1/2, and PER1 were unchanged. The fold change in BMAL1 correlated with insulin sensitivity (r = 0.43, P = 0.044), BMI (r = −0.44, P = 0.042), and body weight changes (r = −0.44, P = 0.039). |
| Dijckmans, 2017 | observational study | cognitive impairment group (n = 30) and normal group(n = 30) (mean age = 70.6) | None | Cortisol circadian rhythms | Better cognitive function was associated with better physical performance. A greater variance in cortisol levels across the day from morning to evening was associated with better cognitive function and physical performance. |
| Garfinkel, 2011 | RCT | 36 independently living T2DM patients with insomnia (mean age = 63) | Period 1: treatment with prolonged-release melatonin (2 mg) or placebo for 3 weeks Period 2: treatment for another 3 weeks after a one-week washout periodPeriod 3: treatment for an extension period of 5 months | Sleep efficiency, wake time after sleep onset, and number of awakenings | 3 weeks of prolonged-release melatonin treatment improved sleep quality. Following 5 months of prolonged-release melatonin treatment, HbA1c was significantly lower than at baseline (9.13% ± 1.55% versus 8.47% ± 1.67%, respectively, P = 0.005). |
| Asayama, 2003 | RCT | 20 AD patients divided into placebo group (n = 9) and melatonin group (n = 11) (mean age = 79.2 years) | Melatonin (3 mg) were given at 8.30 p.m. each day for 4 weeks | sleep time and activity | Melatonin administration had the effect of improving sleep time, night activity and ADAS scores |
| Zarifkar, 2017 | RCT | 59 newly diagnosed T2DM patients (30 in the metformin group and 29 in the pioglitazone group | Treatment with either metformin (1000 mg daily) or pioglitazone (30 mg daily) for 3 months | orexin | Three-month anti-hyperglycemic treatment with proportionate doses of metformin or pioglitazone both improve insulin resistance and glycemic control. A negative association between peripheral orexin concentrations and insulin resistance was observed in T2DM patients. |
| Herring, 2020 | RCT | 285 participants with AD and insomnia (suvorexant, N = 142; placebo, N = 141, mean age = 69) | Treatment with 10 mg suvorexant (an orexin antagonist) (could be increased to 20 mg based on clinical response) or a placebo for 4 weeks | total sleep time | Suvorexant improved total sleep time in patients with probable AD dementia and insomnia. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Fan, R.; Xie, L.; Shi, X.; Dong, K.; Zhang, S.; Tao, J.; Xu, W.; Ma, D.; Chen, J.; et al. A Growing Link between Circadian Rhythms, Type 2 Diabetes Mellitus and Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 504. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010504
Peng X, Fan R, Xie L, Shi X, Dong K, Zhang S, Tao J, Xu W, Ma D, Chen J, et al. A Growing Link between Circadian Rhythms, Type 2 Diabetes Mellitus and Alzheimer’s Disease. International Journal of Molecular Sciences. 2022; 23(1):504. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010504
Chicago/Turabian StylePeng, Xuemin, Rongping Fan, Lei Xie, Xiaoli Shi, Kun Dong, Shujun Zhang, Jing Tao, Weijie Xu, Delin Ma, Juan Chen, and et al. 2022. "A Growing Link between Circadian Rhythms, Type 2 Diabetes Mellitus and Alzheimer’s Disease" International Journal of Molecular Sciences 23, no. 1: 504. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010504