
Per1/Per2 Disruption Reduces Testosterone Synthesis and Impairs Fertility in Elderly Male Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
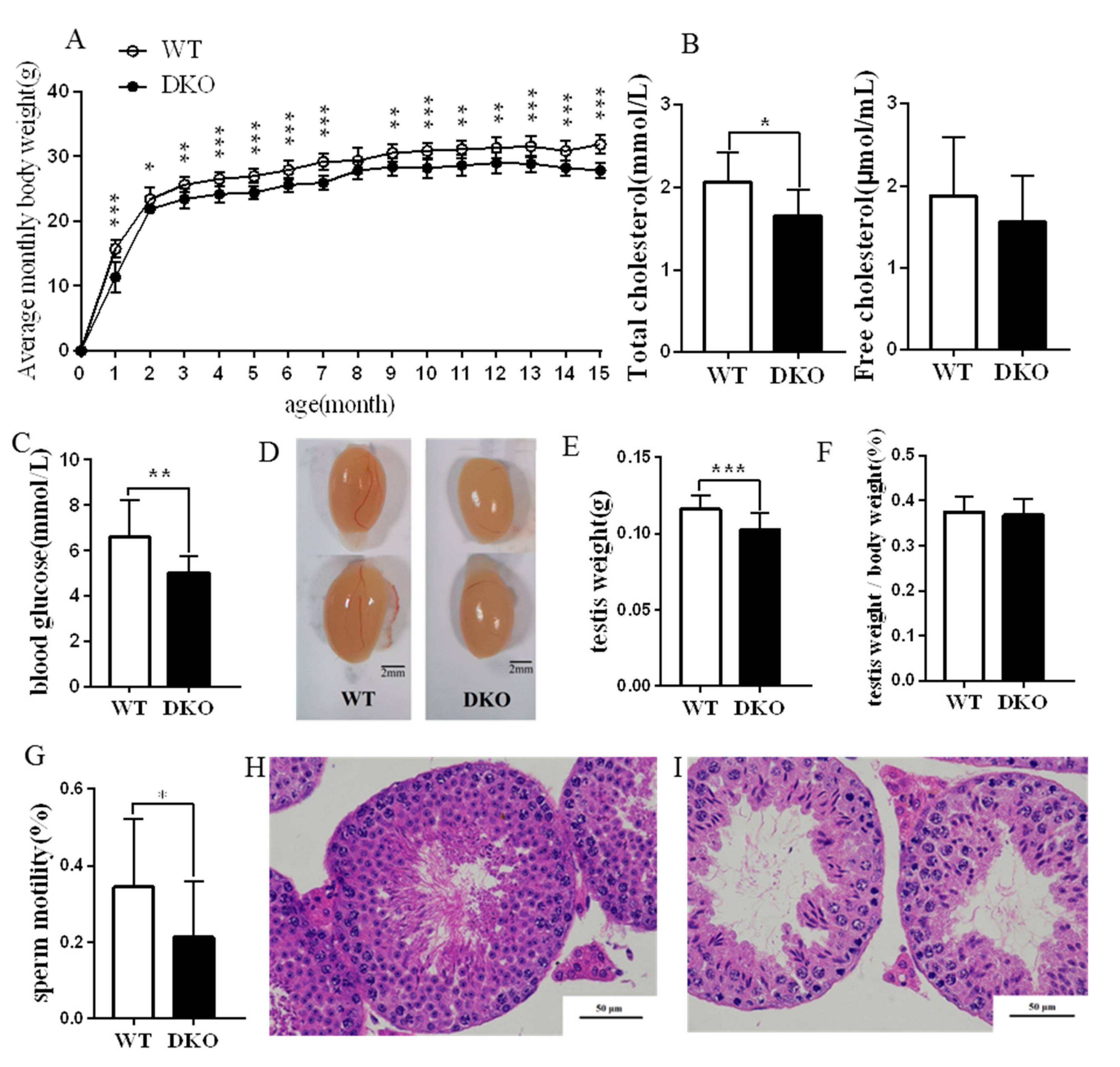
2.1. Physiological Effects of Per1/Per2 Double Knockout
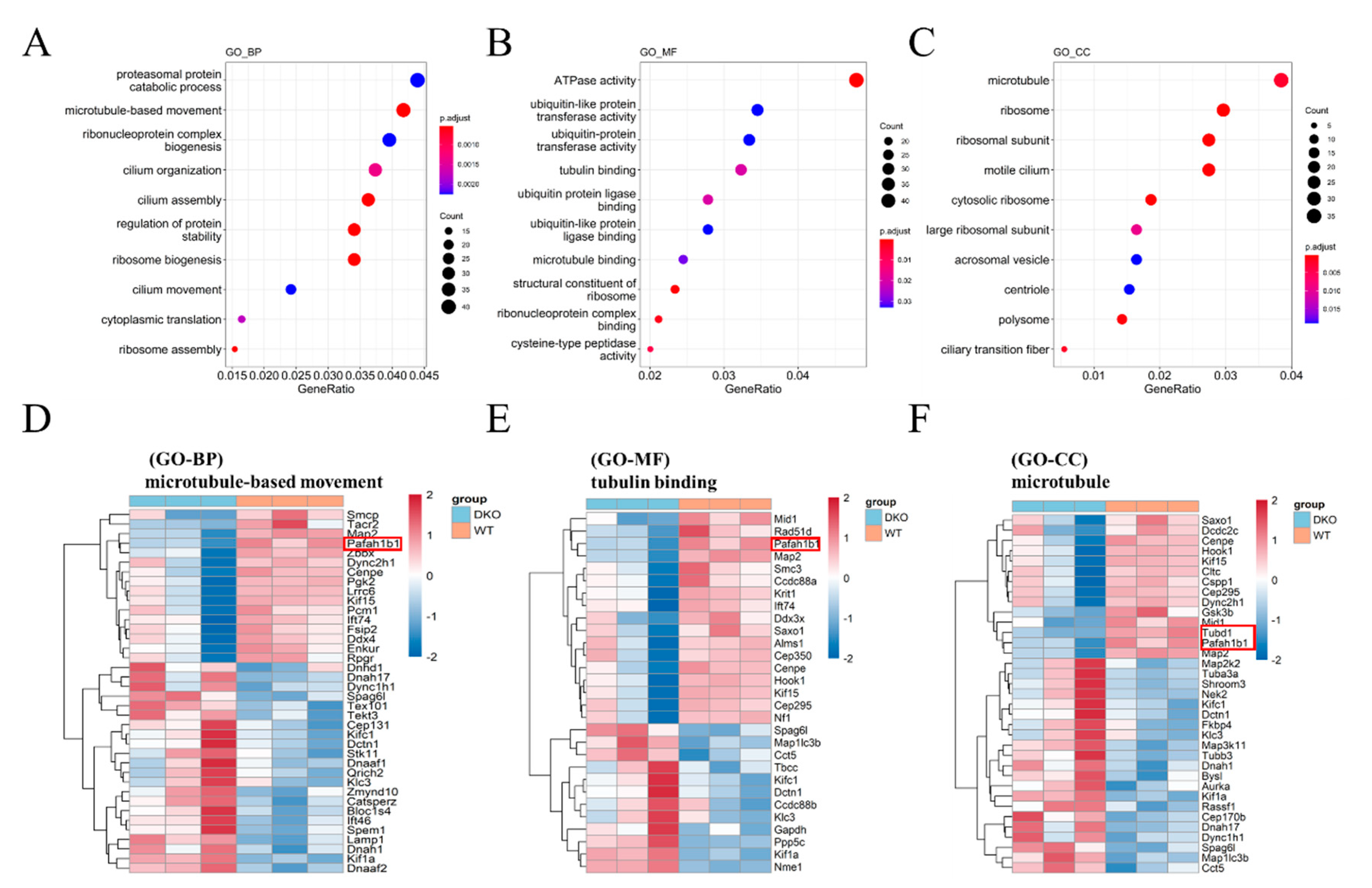
2.2. Functional Enrichment Analysis of Differentially Expressed Genes between WT and DKO Mice
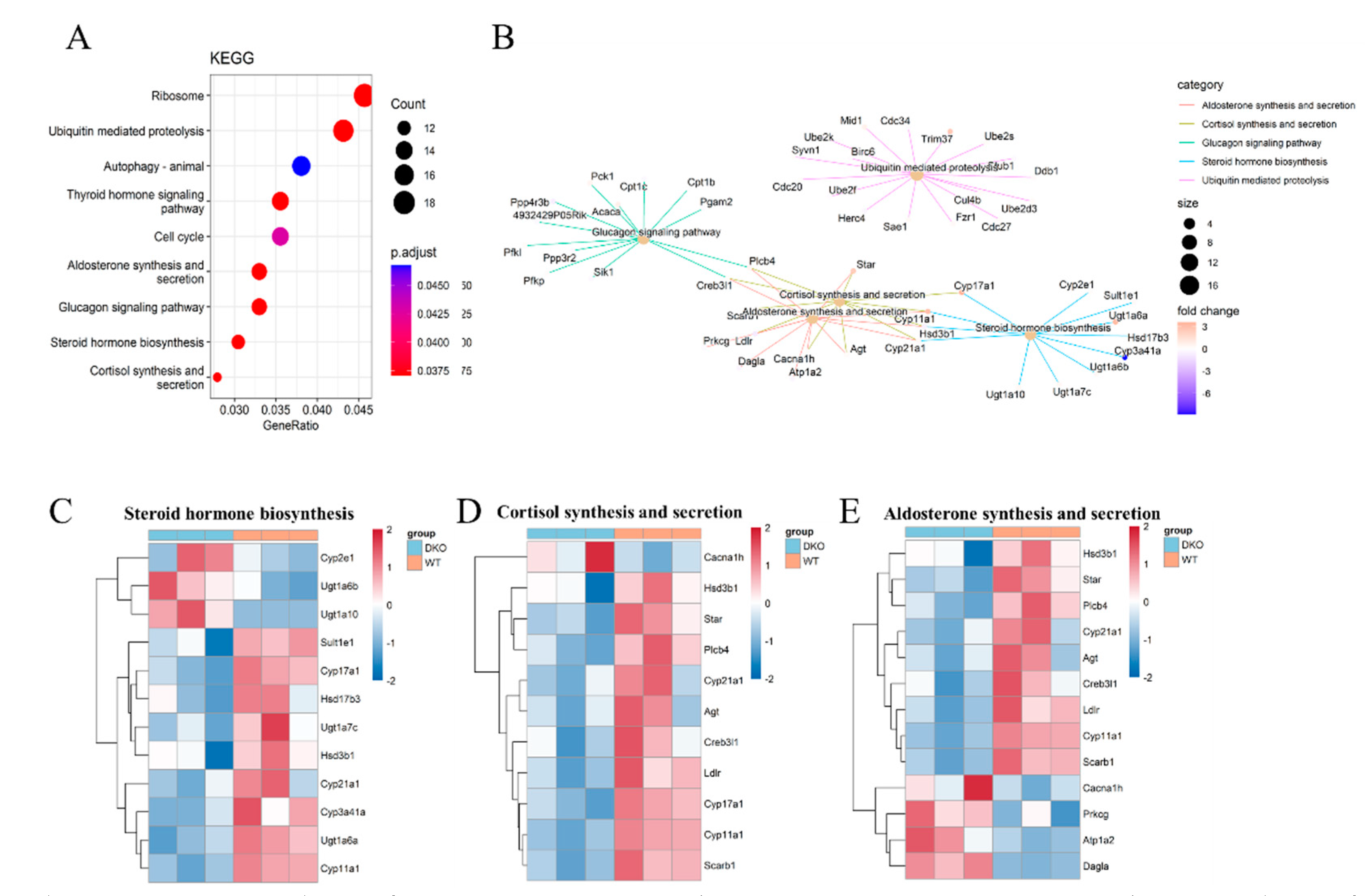
2.3. Transcriptome Sequencing Revealed Differences in Steroid Hormone Synthesis in Testicular Tissue
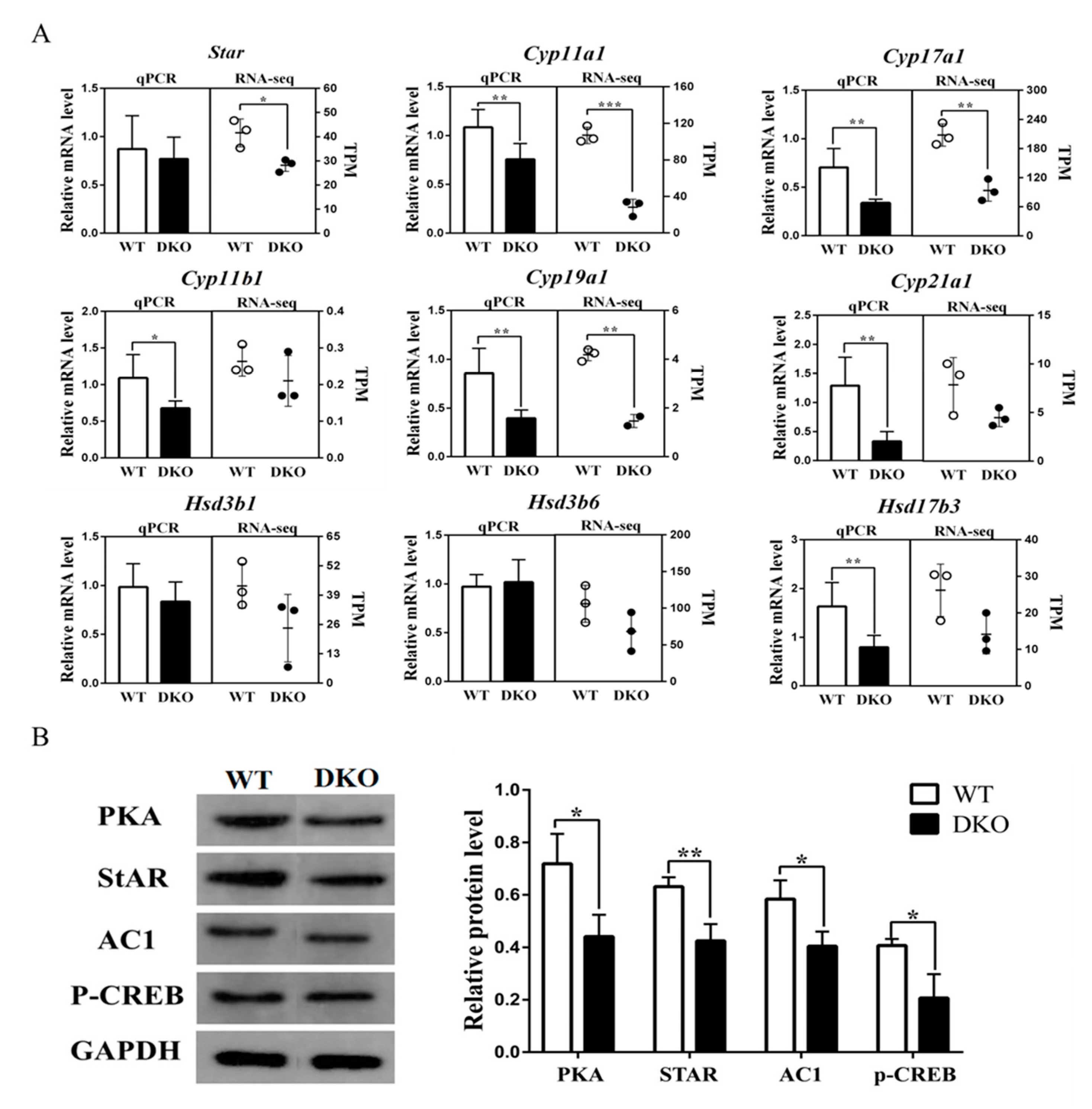
2.4. Per1/Per2 Double Knockout Affected the Expression of Genes Related to Steroid Biosynthesis Signaling Pathway
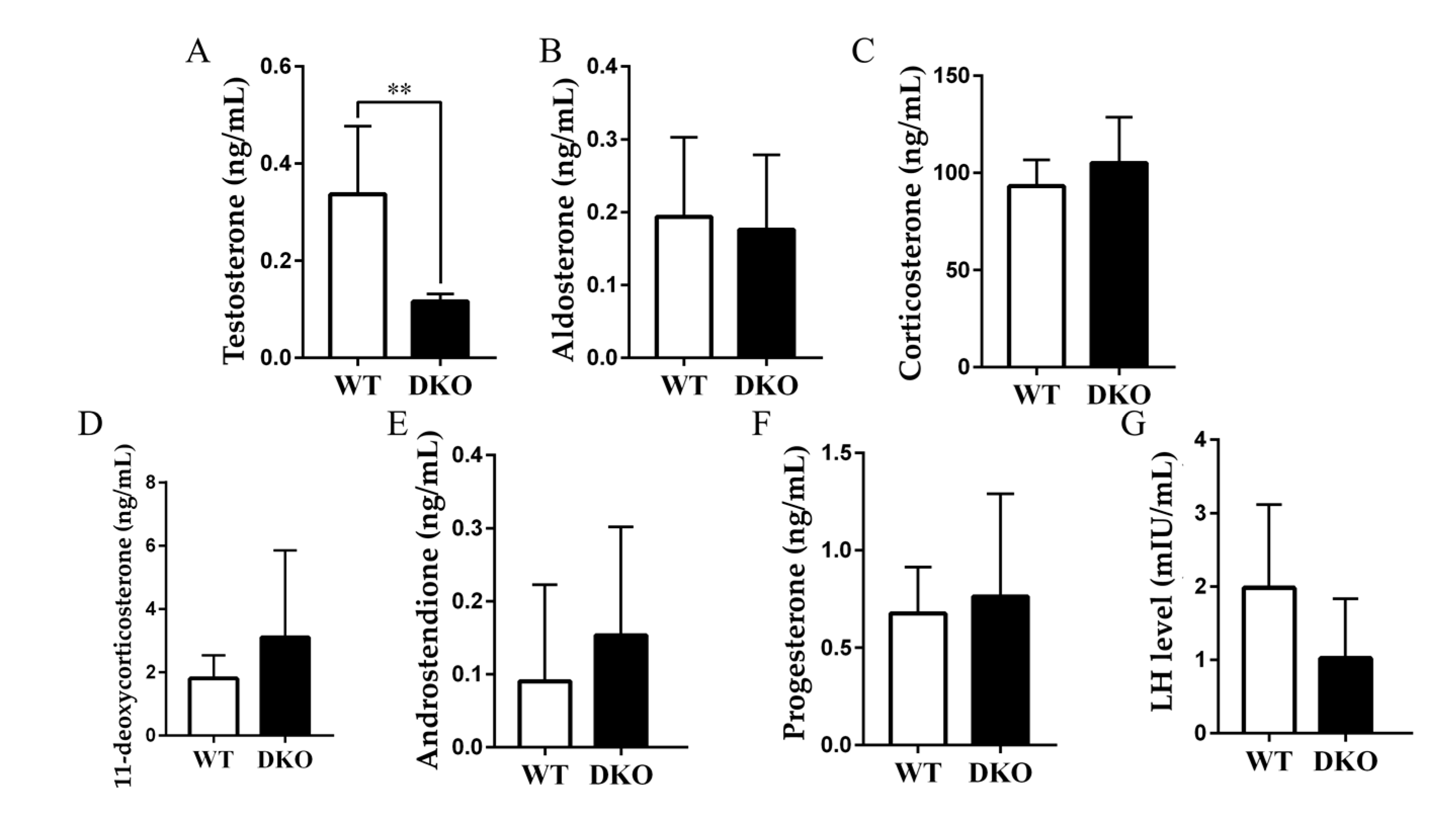
2.5. Plasma Testosterone Decreased in Per1/Per2 Double Knockout Male Mice
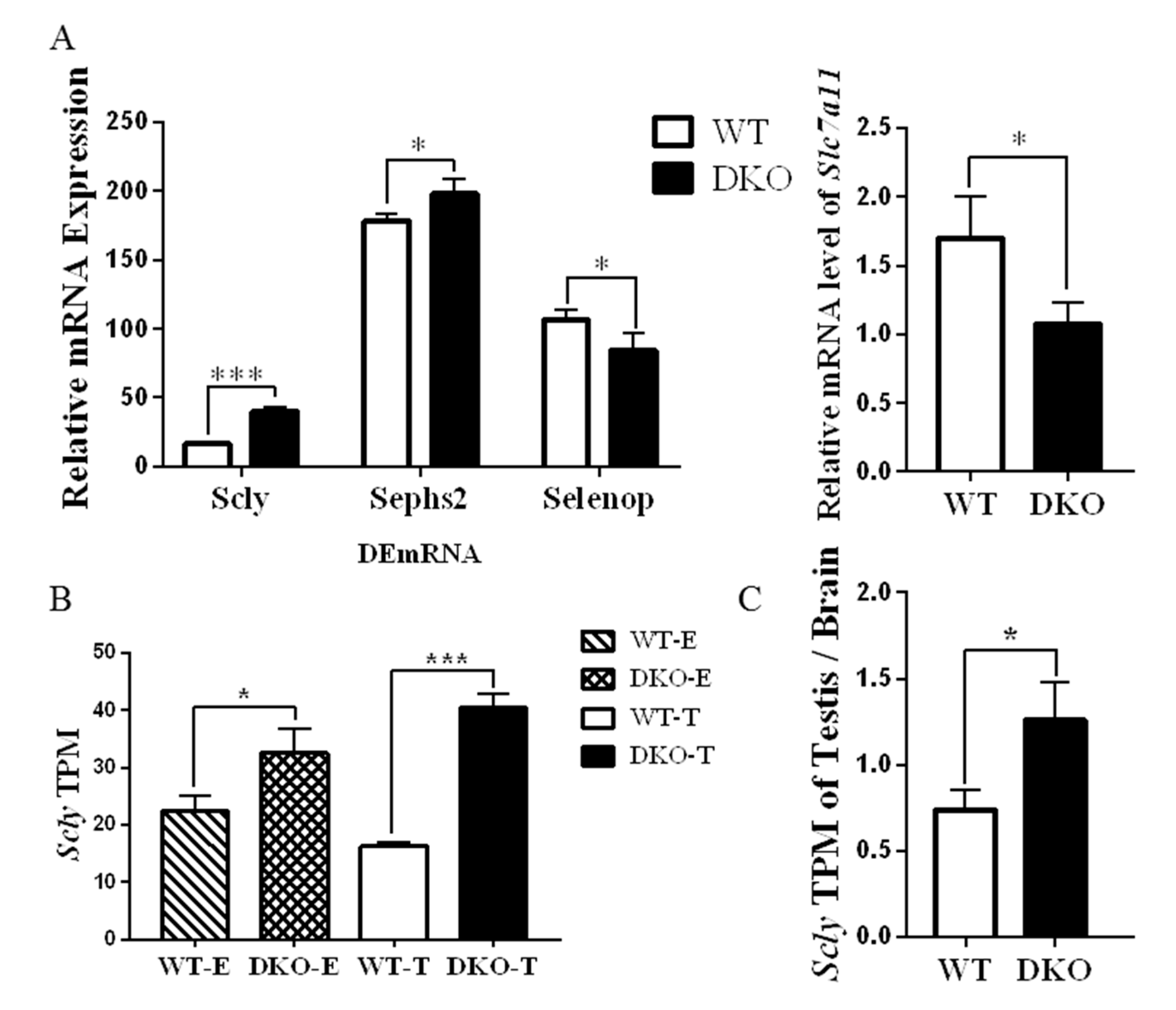
2.6. Genes Related to Production and Metabolism of Selenoprotein in Per1/Per2 Double Knockout Male Mice
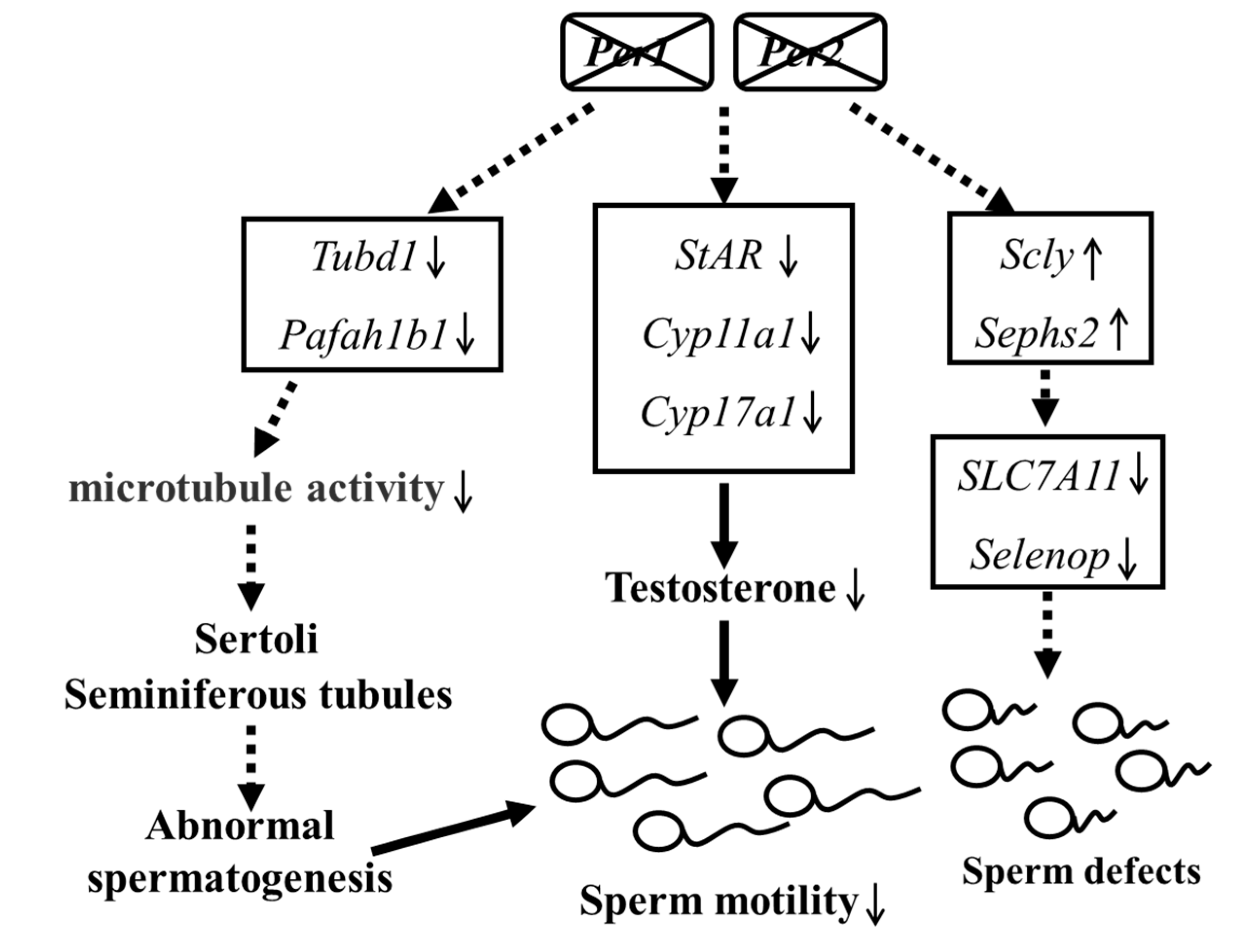
3. Discussion
4. Materials and Methods
4.1. Animal Housing
4.2. Body Weight and Feeding
4.3. Relative Sperm Counts and Motility
4.4. Histological Studies
4.5. LC/MS
4.6. RNA Isolation, Library Preparation, and Sequencing
4.7. RNAseq Data Analysis
4.8. Real-Time PCR Analysis
4.9. Western Blot Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasegawa, S.; Fukushima, H.; Hosoda, H.; Serita, T.; Ishikawa, R.; Rokukawa, T.; Kawahara-Miki, R.; Zhang, Y.; Ohta, M.; Okada, S.; et al. Hippocampal clock regulates memory retrieval via Dopamine and PKA-induced GluA1 phosphorylation. Nat. Commun. 2019, 10, 5766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Song, L.; Liu, M.; Ge, R.; Zhou, Q.; Liu, W.; Li, R.; Qie, J.; Zhen, B.; Wang, Y.; et al. A proteomics landscape of circadian clock in mouse liver. Nat. Commun. 2018, 9, 1553. [Google Scholar] [CrossRef] [PubMed]
- Bass, J.; Takahashi, J. Circadian integration of metabolism and energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [Green Version]
- Stigliani, S.; Massarotti, C.; De Leo, C.; Maccarini, E.; Sozzi, F.; Cagnacci, A.; Anserini, P.; Scaruffi, P. Fifteen Year Regional Center Experience in Sperm Banking for Cancer Patients: Use and Reproductive Outcomes in Survivors. Cancers 2021, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Cheng, S.; Jiang, X.; He, X.; Wang, Y.; Jiang, Z.; Hou, W.; Li, S.; Liu, Y.; Wang, Z. The Noncircadian Function of the Circadian Clock Gene in the Regulation of Male Fertility. J. Biol. Rhythm. 2013, 28, 208–217. [Google Scholar]
- Alvarez, J.; Hansen, A.; Ord, T.; Bębas, P.; Chappell, P.E.; Giebultowicz, J.M.; Williams, C.; Moss, S.; Sehgal, A. The Circadian Clock Protein BMAL1 Is Necessary for Fertility and Proper Testosterone Production in Mice. J. Biol. Rhythm. 2008, 23, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Pilorz, V.; Steinlechner, S. Low reproductive success in Per1 and Per2 mutant mouse females due to accelerated ageing? Reproduction 2008, 135, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Liu, C.; Li, Y.; Jiang, H.; Yang, P.; Tang, J.; Xu, Y.; Wang, H.; He, Y. Loss-of-function mutations with circadian rhythm regulator Per1/Per2 lead to premature ovarian insufficiency†. Biol. Reprod. 2019, 100, 1066–1072. [Google Scholar] [CrossRef]
- Clermont, Y. Kinetics of spermatogenesis in mammals: Seminiferous epithelium cycle and spermatogonial renewal. Physiol. Rev. 1972, 52, 198–236. [Google Scholar] [CrossRef]
- Hayden, R.; Flannigan, R.; Schlegel, P. The Role of Lifestyle in Male Infertility: Diet, Physical Activity, and Body Habitus. Curr. Urol. Rep. 2018, 19, 56. [Google Scholar] [CrossRef]
- Symons, A.; Arendt, J.; Pryde, S. Differential effects of melatonin on the stimulated release of LH from dispersed pituitary cells of the prepubertal female rat. J. Endocrinol. 1985, 107, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.; Chen, D.; Storer, E.; Sehgal, A. Non-cyclic and Developmental Stage-Specific Expression of Circadian Clock Proteins During Murine Spermatogenesis1. Biol. Reprod. 2003, 69, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittman, E.L.; Doherty, L.; Huang, L.; Paroskie, A. Period gene expression in mouse endocrine tissues. Am. J. Physiol. Integr. Comp. Physiol. 2003, 285, R561–R569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, D.; Cermakian, N.; Brancorsini, S.; Parvinen, M.; Sassone-Corsi, P. No Circadian Rhythms in Testis: Period1 Expression Is Clock Independent and Developmentally Regulated in the Mouse. Mol. Endocrinol. 2003, 17, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Utzinger, K.S.; Blickenstorfer, K.; Leeners, B. Diurnal and seasonal changes in semen quality of men in subfertile partnerships. Chronobiol. Int. 2018, 35, 1375–1384. [Google Scholar] [CrossRef]
- Baburski, A.; Andric, S.; Kostic, T. Luteinizing hormone signaling is involved in synchronization of Leydig cell’s clock and is crucial for rhythm robustness of testosterone production. Biol. Reprod. 2019, 100, 1406–1415. [Google Scholar] [CrossRef]
- Tenover, J.S.; Matsumoto, A.M.; Clifton, D.K.; Bremner, W.J. Age-Related Alterations in the Circadian Rhythms of Pulsatile Luteinizing Hormone and Testosterone Secretion in Healthy Men. J. Gerontol. 1988, 43, M163–M169. [Google Scholar] [CrossRef]
- Schill, W. Fertility and sexual life of men after their forties and in older age. Asian J. Androl. 2001, 3, 1–7. [Google Scholar]
- Balay, S.D.; Hochstoeger, T.; Vilceanu, A.; Malkemper, E.P.; Snider, W.; Dürnberger, G.; Mechtler, K.; Schuechner, S.; Ogris, E.; Nordmann, G.C.; et al. The expression, localisation and interactome of pigeon CRY2. Sci. Rep. 2021, 11, 1–13. [Google Scholar]
- Li, Y.; Khanal, P.; Norheim, F.; Hjorth, M.; Bjellaas, T.; Drevon, C.A.; Vaage, J.; Kimmel, A.R.; Dalen, K.T. Plin2 deletion increases cholesteryl ester lipid droplet content and disturbs cholesterol balance in adrenal cortex. J. Lipid Res. 2021, 62, 100048. [Google Scholar] [CrossRef]
- Miller, W. Molecular biology of steroid hormone synthesis. Endocr. Rev. 1988, 9, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.; Short, K.; Hooper, S. The science of steroids. Semin. Fetal Neonatal Med. 2019, 24, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, V.; Stocco, D.; Clark, B. Current knowledge on the acute regulation of steroidogenesis. Biol. Reprod. 2018, 99, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Preston, B.T.; Stevenson, I.R.; Lincoln, G.A.; Monfort, S.L.; Pilkington, J.G.; Wilson, K. Testes size, testosterone production and reproductive behaviour in a natural mammalian mating system. J. Anim. Ecol. 2011, 81, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Carlisle, A.E.; Lee, N.; Matthew-Onabanjo, A.N.; Spears, M.E.; Park, S.J.; Youkana, D.; Doshi, M.B.; Peppers, A.; Li, R.; Joseph, A.B.; et al. Selenium detoxification is required for cancer-cell survival. Nat. Metab. 2020, 2, 603–611. [Google Scholar] [CrossRef]
- Griswold, M. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 1998, 9, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaum, N.; Lehallier, B.; Hahn, O.; Pálovics, R.; Hosseinzadeh, S.; Lee, S.E.; Sit, R.; Lee, D.P.; Losada, P.M.; Zardeneta, M.E.; et al. Ageing hallmarks exhibit organ-specific temporal signatures. Nature 2020, 583, 596–602. [Google Scholar] [CrossRef]
- Griswold, M. 50 years of spermatogenesis: Sertoli cells and their interactions with germ cells. Biol. Reprod. 2018, 99, 87–100. [Google Scholar] [CrossRef]
- Mori, Y.; Takashima, S.; Kanatsu-Shinohara, M.; Yi, Z.; Shinohara, T. Cdc42 is required for male germline niche development in mice. Cell Rep. 2021, 36, 109550. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Kohn, T.P.; Santiago, J.E.; Sigalos, J.T.; Kirby, E.W.; Hockenberry, M.S.; Pickett, S.M.; Pastuszak, A.W.; Lipshultz, L.I. Increased Risk of Hypogonadal Symptoms in Shift Workers With Shift Work Sleep Disorder. Urology 2020, 138, 52–59. [Google Scholar] [CrossRef]
- Bailey, M.; Silver, R. Sex differences in circadian timing systems: Implications for disease. Front. Neuroendocrinol. 2014, 35, 111–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bain, J.; Duthie, M.; Keene, J. Relationship of seminal plasma testosterone and dihydrotestosterone to sperm count and motility in man. Arch. Androl. 1979, 2, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.; Walker, W. The regulation of spermatogenesis by androgens. Semin. Cell Dev. Biol. 2014, 30, 2–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, W. Non-classical actions of testosterone and spermatogenesis. Philos. Trans. R Soc. B Biol. Sci. 2010, 365, 1557–1569. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.; Bose, H. Early steps in steroidogenesis: Intracellular cholesterol trafficking. J. Lipid Res. 2011, 52, 2111–2135. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Collins, D.; Schleicher, R. Sources of cholesterol for testosterone biosynthesis in murine Leydig cells. Endocrinology 1990, 127, 2047–2055. [Google Scholar]
- Miller, W. Steroidogenic acute regulatory protein (StAR), a novel mitochondrial cholesterol transporter. Biochim. Et Biophys. Acta 2007, 1771, 663–676. [Google Scholar] [CrossRef]
- Saito, Y. Selenium Transport Mechanism via Selenoprotein P-Its Physiological Role and Related Diseases. Front. Nutr. 2021, 8, 685517. [Google Scholar] [CrossRef]
- Behne, D.; Höfer, T.; Von Berswordt-Wallrabe, R.; Elger, W. Selenium in the Testis of the Rat: Studies on Its Regulation and Its Importance for the Organism. J. Nutr. 1982, 112, 1682–1687. [Google Scholar] [CrossRef]
- Shetty, S.; Marsicano, J.; Copeland, P. Uptake and Utilization of Selenium from Selenoprotein P. Biol. Trace Elem. Res. 2018, 181, 54–61. [Google Scholar] [CrossRef]
- Olson, G.E.; Winfrey, V.P.; Nagdas, S.K.; Hill, K.E.; Burk, R.F. Selenoprotein P Is Required for Mouse Sperm Development. Biol. Reprod. 2005, 73, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Salaberry, N.L.; Hamm, H.; Felder-Schmittbuhl, M.-P.; Mendoza, J. A suprachiasmatic-independent circadian clock(s) in the habenula is affected by Per gene mutations and housing light conditions in mice. Anat. Embryol. 2018, 224, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Parasram, K.; Bernardon, N.; Hammoud, M.; Chang, H.; He, L.; Perrimon, N.; Karpowicz, P. Intestinal Stem Cells Exhibit Conditional Circadian Clock Function. Stem Cell Rep. 2018, 11, 1287–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and Metabolic Syndrome in Circadian Clock Mutant Mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Shen, W.; Chen, C.; Wang, Q.; Lu, Q.; Shao, W.; Jiang, Z.; Hu, H. Circadian Rhythm Disruption Influenced Hepatic Lipid Metabolism, Gut Microbiota and Promoted Cholesterol Gallstone Formation in Mice. Front. Endocrinol. 2021, 12, 723918. [Google Scholar] [CrossRef]
- O’Donnell, L.; O’Bryan, M. Microtubules and spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 45–54. [Google Scholar] [CrossRef]
- Zheng, B.; Albrecht, U.; Kaasik, K.; Sage, M.; Lu, W.; Vaishnav, S.; Li, Q.; Sun, Z.S.; Eichele, G.; Bradley, A.; et al. Nonredundant Roles of the mPer1 and mPer2 Genes in the Mammalian Circadian Clock. Cell 2001, 105, 683–694. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Larkin, D.W.; Albrecht, U.; Sun, Z.; Sage, M.; Eichele, G.; Lee, C.C.; Bradley, A. The mPer2 gene encodes a functional component of the mammalian circadian clock. Nature 1999, 400, 169–173. [Google Scholar] [CrossRef]
- Peng, J.; Shen, L.; Chen, J.; Cao, X.; Zhou, Y.; Weng, H.; Long, C.; Zhang, D.; Tu, S.; Zhang, Y.; et al. New discovery of cryptorchidism: Decreased retinoic acid in testicle. Saudi Pharm. J. 2016, 24, 279–285. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Wang, H.; Wang, H.; Li, N.; He, R.; Liu, Z. Per1/Per2 Disruption Reduces Testosterone Synthesis and Impairs Fertility in Elderly Male Mice. Int. J. Mol. Sci. 2022, 23, 7399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137399
Liu Q, Wang H, Wang H, Li N, He R, Liu Z. Per1/Per2 Disruption Reduces Testosterone Synthesis and Impairs Fertility in Elderly Male Mice. International Journal of Molecular Sciences. 2022; 23(13):7399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137399
Chicago/Turabian StyleLiu, Qinrui, Hu Wang, Hualin Wang, Na Li, Ruyi He, and Zhiguo Liu. 2022. "Per1/Per2 Disruption Reduces Testosterone Synthesis and Impairs Fertility in Elderly Male Mice" International Journal of Molecular Sciences 23, no. 13: 7399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137399