
The Potential Role of SP-G as Surface Tension Regulator in Tear Film: From Molecular Simulations to Experimental Observations

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
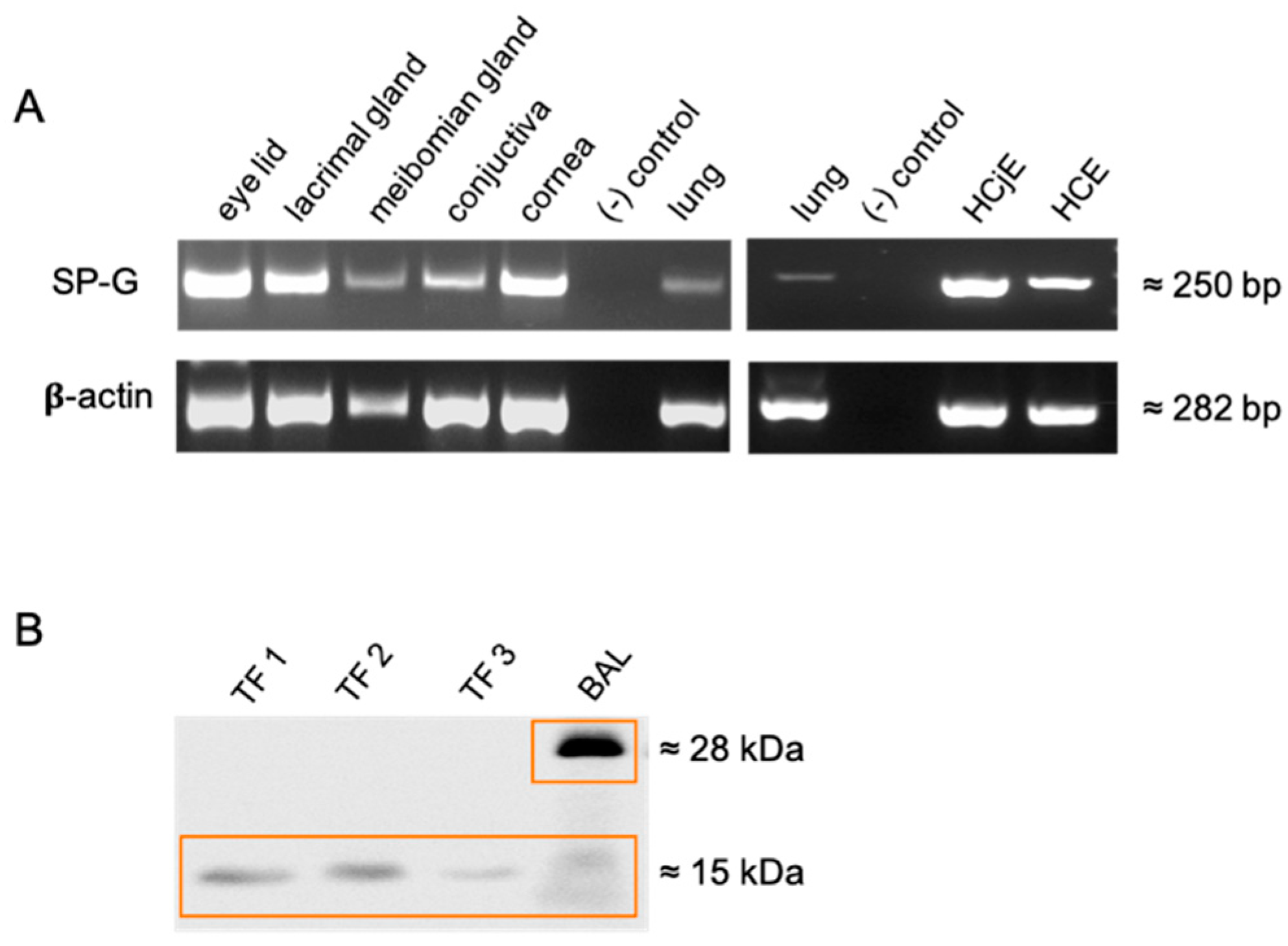
2.1. Detection of SP-G in Tissue of the Ocular Surface, Lacrimal Apparatus and in Tears
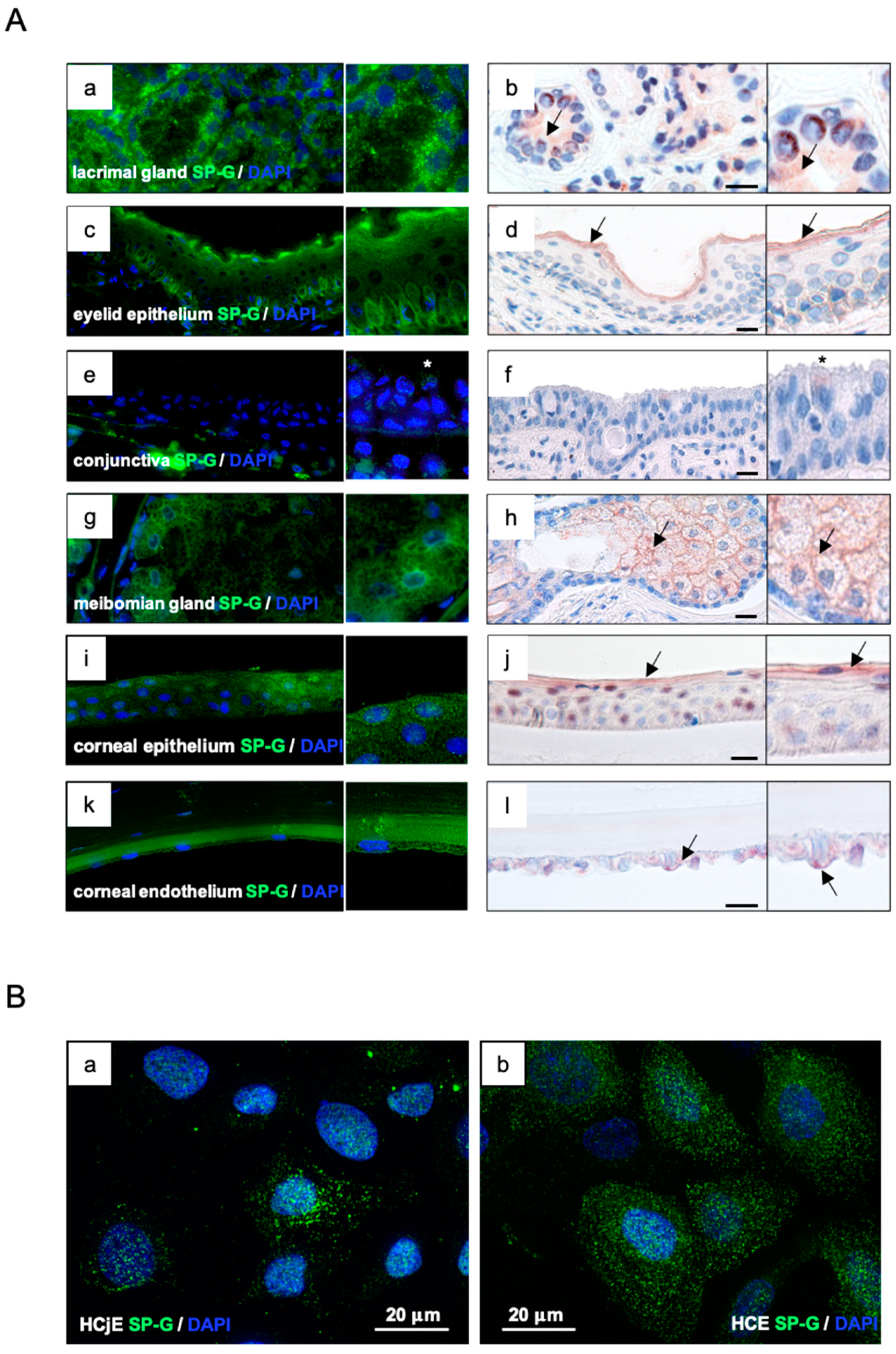
2.2. Localization of SP-G within the Tissue of the Ocular Surface and in Cultured HCE and HCjE Cells
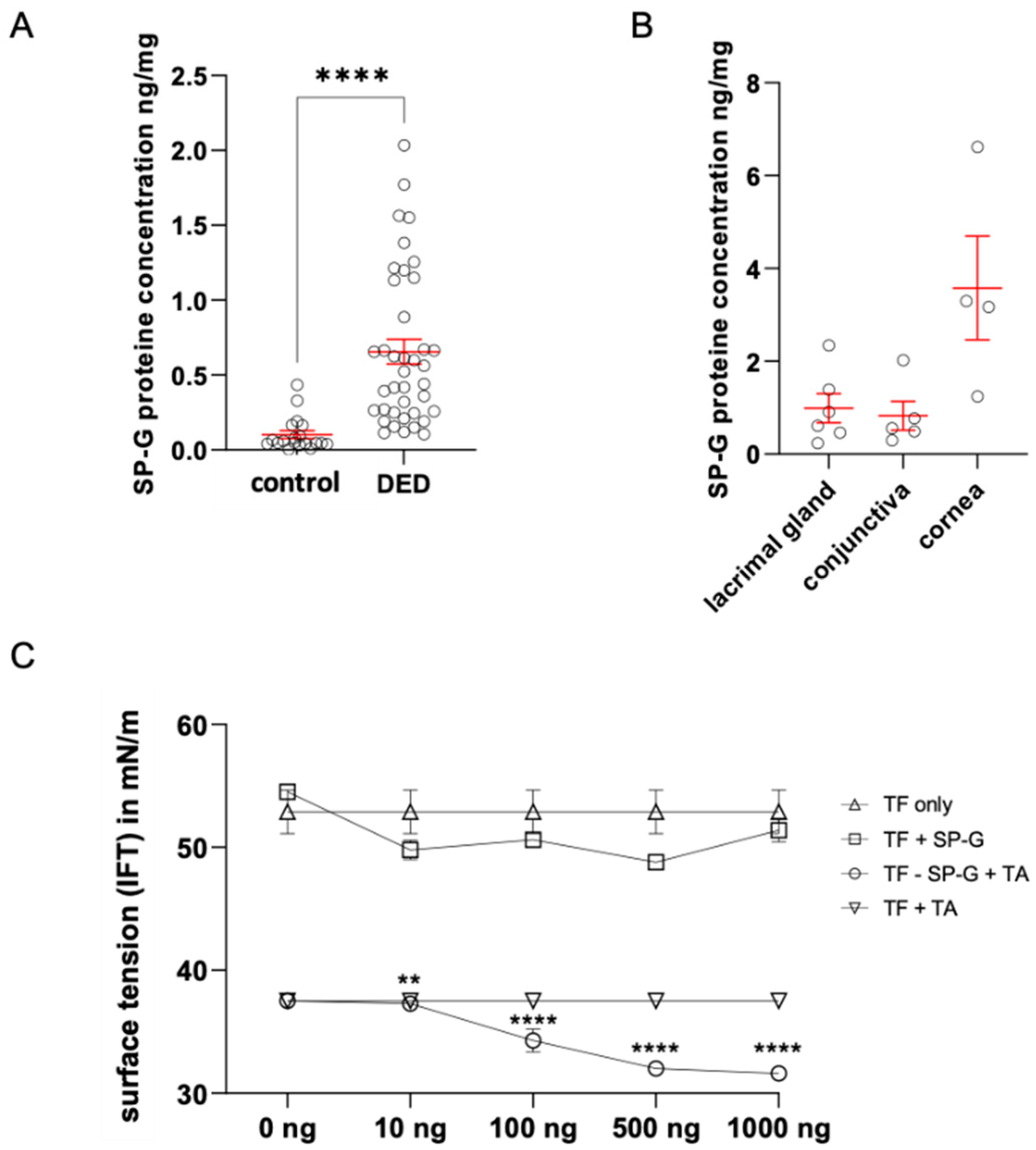
2.3. Quantification of SP-G in Tears by ELISA and Determination of Tear Surface Tension as a Function of SP-G Protein Concentration
2.4. Putative Surface Activity of SP-G in Tears
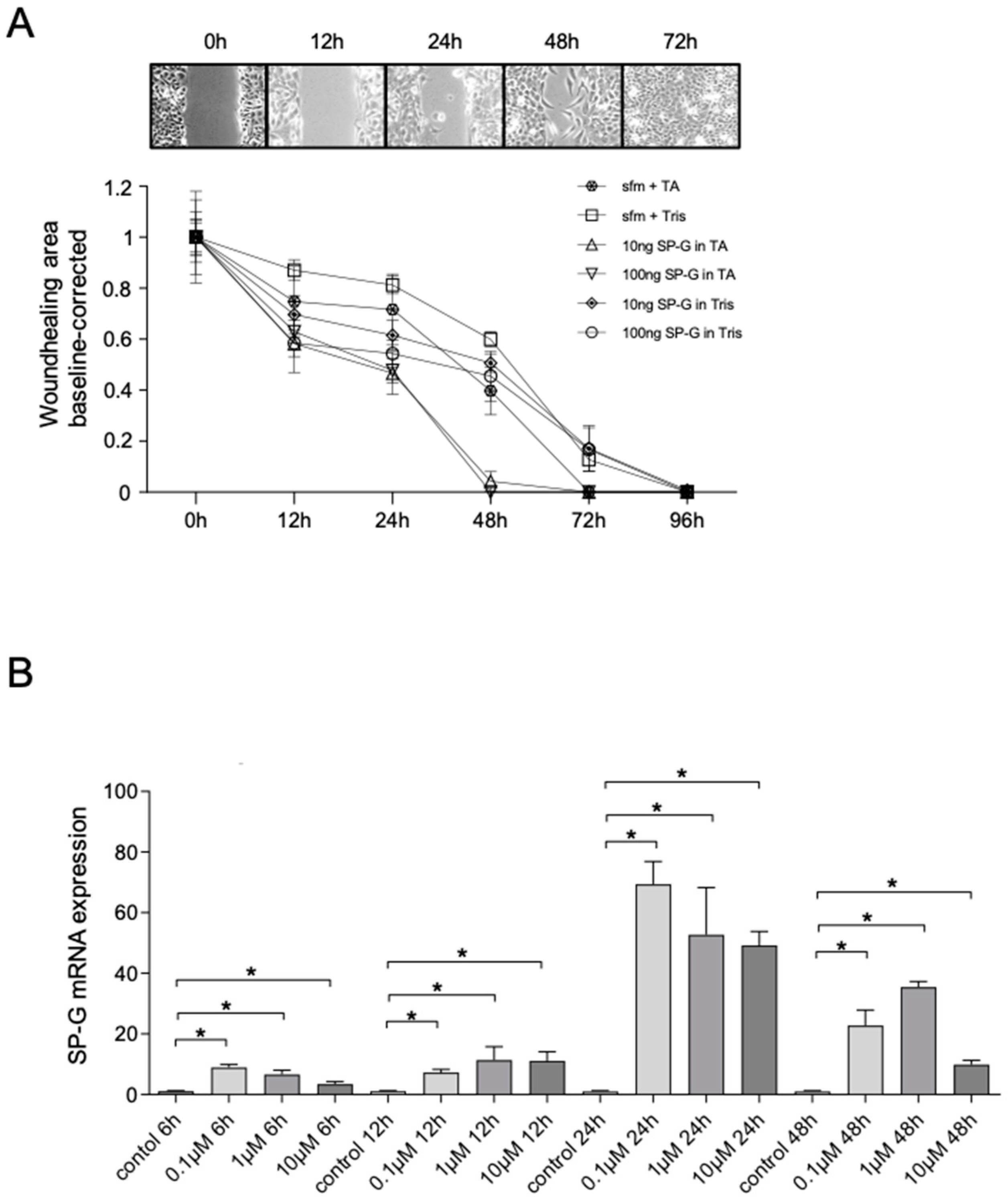
2.5. In Vitro Wound Healing Scratch Assay
2.6. Effects of Cortisol on SP-G mRNA Levels
2.7. In Silico Molecular-Level Insight into Interactions of SP-G with Lipids of the Tear Film Lipid Layer (TFLL)
3. Discussion
4. Materials and Methods
4.1. Generation of Samples
4.2. Cell Culture
4.3. mRNA Extraction and cDNA Synthesis
4.4. Polymerase Chain Reaction (PCR)
4.5. Quantitative Real-Time RT-PCR
4.6. Western Blot
4.7. Immunohistochemistry
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. In Vitro Wound Healing Scratch Assay
4.10. Interfacial Tension
4.11. Molecular Dynamics Simulations
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Craig, J.P.; Nelson, J.D.; Azar, D.T.; Belmonte, C.; Bron, A.J.; Chauhan, S.K.; de Paiva, C.S.; Gomes, J.A.P.; Hammitt, K.M.; Jones, L.; et al. TFOS DEWS II Report Executive Summary. Ocul. Surf. 2017, 15, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Uchino, M.; Schaumberg, D.A. Dry Eye Disease: Impact on Quality of Life and Vision. Curr. Ophthalmol. Rep. 2013, 1, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, H.; Sato, T.; Akimoto, N.; Ogura, T.; Ito, A. Identification and characterization of ABCB1-mediated and non-apoptotic sebum secretion in differentiated hamster sebocytes. Biochim. Et Biophys. Acta 2011, 1811, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef]
- Paulsen, F.; Garreis, F. What drives Meibomian gland disease? Arch. Soc. Esp. Oftalmol. 2014, 89, 175–176. [Google Scholar] [CrossRef]
- Nelson, J.D.; Shimazaki, J.; Benitez-del-Castillo, J.M.; Craig, J.P.; McCulley, J.P.; Den, S.; Foulks, G.N. The international workshop on meibomian gland dysfunction: Report of the definition and classification subcommittee. Investig. Ophthalmol. Vis. Sci 2011, 52, 1930–1937. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, J.; Garreis, F.; Paulsen, F. Pathophysiology of Meibomian Glands—An Overview. Ocul. Immunol. Inflamm. 2021, 29, 803–810. [Google Scholar] [CrossRef]
- Green-Church, K.B.; Butovich, I.; Willcox, M.; Borchman, D.; Paulsen, F.; Barabino, S.; Glasgow, B.J. The international workshop on meibomian gland dysfunction: Report of the subcommittee on tear film lipids and lipid-protein interactions in health and disease. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1979–1993. [Google Scholar] [CrossRef] [Green Version]
- Holly, F.J.; Lemp, M.A. Tear physiology and dry eyes. Surv. Ophthalmol. 1977, 22, 69–87. [Google Scholar] [CrossRef]
- Wright, J.R. Immunoregulatory functions of surfactant proteins. Nat. Rev. Immunol. 2005, 5, 58–68. [Google Scholar] [CrossRef]
- Crouch, E.; Wright, J.R. Surfactant proteins a and d and pulmonary host defense. Annu. Rev. Physiol. 2001, 63, 521–554. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.H.; Possmayer, F. Role of bovine pulmonary surfactant-associated proteins in the surface-active property of phospholipid mixtures. Biochim. Biophys. Acta 1990, 1046, 233–241. [Google Scholar] [CrossRef]
- Rausch, F.; Schicht, M.; Paulsen, F.; Ngueya, I.; Bräuer, L.; Brandt, W. “SP-G”, a putative new surfactant protein-tissue localization and 3D structure. PLoS ONE 2012, 7, e47789. [Google Scholar] [CrossRef]
- Schicht, M.; Rausch, F.; Finotto, S.; Mathews, M.; Mattil, A.; Schubert, M.; Koch, B.; Traxdorf, M.; Bohr, C.; Worlitzsch, D.; et al. SFTA3, a novel protein of the lung: Three-dimensional structure, characterisation and immune activation. Eur. Respir. J. 2014, 44, 447–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bräuer, L.; Kindler, C.; Jäger, K.; Sel, S.; Nölle, B.; Pleyer, U.; Ochs, M.; Paulsen, F.P. Detection of Surfactant Proteins A and D in Human Tear Fluid and the Human Lacrimal System. Investig. Ophthalmol. Vis. Sci 2007, 48, 3945–3953. [Google Scholar] [CrossRef] [PubMed]
- Bräuer, L.; Johl, M.; Börgermann, J.; Pleyer, U.; Tsokos, M.; Paulsen, F.P. Detection and Localization of the Hydrophobic Surfactant Proteins B and C in Human Tear Fluid and the Human Lacrimal System. Curr. Eye Res. 2007, 32, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Schicht, M.; Posa, A.; Paulsen, F.; Bräuer, L. The ocular surfactant system and its relevance in the dry eye. Klin. Mon. Fur Augenheilkd. 2010, 227, 864–870. [Google Scholar] [CrossRef]
- Schicht, M.; Garreis, F.; Hartjen, N.; Beileke, S.; Jacobi, C.; Sahin, A.; Holland, D.; Schroder, H.; Hammer, C.M.; Paulsen, F.; et al. SFTA3—A novel surfactant protein of the ocular surface and its role in corneal wound healing and tear film surface tension. Sci. Rep. 2018, 8, 9791. [Google Scholar] [CrossRef]
- Mittal, R.A.; Hammel, M.; Schwarz, J.; Heschl, K.M.; Bretschneider, N.; Flemmer, A.W.; Herber-Jonat, S.; Konigshoff, M.; Eickelberg, O.; Holzinger, A. SFTA2-A Novel Secretory Peptide Highly Expressed in the Lung-Is Modulated by Lipopolysaccharide but Not Hyperoxia. PLoS ONE 2012, 7, e40011. [Google Scholar]
- Keicho, N.; Ohashi, J.; Tamiya, G.; Nakata, K.; Taguchi, Y.; Azuma, A.; Ohishi, N.; Emi, M.; Park, M.H.; Inoko, H.; et al. Fine localization of a major disease-susceptibility locus for diffuse panbronchiolitis. Am. J. Hum. Genet. 2000, 66, 501–507. [Google Scholar] [CrossRef] [Green Version]
- Rausch, F.; Schicht, M.; Brauer, L.; Paulsen, F.; Brandt, W. Protein modeling and molecular dynamics simulation of the two novel surfactant proteins SP-G and SP-H. J. Mol. Modeling 2014, 20, 2513. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.P.; Purslow, C.; Murphy, P.J.; Wolffsohn, J.S. Effect of a liposomal spray on the pre-ocular tear film. Cont. Lens Anterior Eye 2010, 33, 83–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, D.B.; O’Callaghan, J.P. Neuroendocrine aspects of the response to stress. Metabolism 2002, 51 (Suppl. S1), 5–10. [Google Scholar] [CrossRef]
- Zhang, Z.; Henzel, W.J. Signal peptide prediction based on analysis of experimentally verified cleavage sites. Protein Sci. 2004, 13, 2819–2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schicht, M.; Rausch, F.; Beron, M.; Jacobi, C.; Garreis, F.; Hartjen, N.; Beileke, S.; Kruse, F.; Bräuer, L.; Paulsen, F. Palate Lung Nasal Clone (PLUNC), a Novel Protein of the Tear Film: Three-Dimensional Structure, Immune Activation, and Involvement in Dry Eye Disease (DED). Investig. Ophthalmol. Vis. Sci 2015, 56, 7312–7323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartshorn, K.L.; Crouch, E.; White, M.R.; Colamussi, M.L.; Kakkanatt, A.; Tauber, B.; Shepherd, V.; Sastry, K.N. Pulmonary surfactant proteins A and D enhance neutrophil uptake of bacteria. Am. J. Physiol 1998, 274 Pt 1, L958–L969. [Google Scholar] [CrossRef]
- Diler, E.; Schicht, M.; Rabung, A.; Tschernig, T.; Meier, C.; Rausch, F.; Garreis, F.; Bräuer, L.; Paulsen, F. The novel surfactant protein SP-H enhances the phagocytosis efficiency of macrophage-like cell lines U937 and MH-S. Bmc Res. Notes 2014, 7, 851. [Google Scholar] [CrossRef] [Green Version]
- Lemp, M.A.; Foulks, G.N. The definition and classification of dry eye disease. Ocul. Surf. 2007, 5, 75–92. [Google Scholar]
- Albertsmeyer, A.-C.; Kakkassery, V.; Spurr-Michaud, S.; Beeks, O.; Gipson, I.K. Effect of pro-inflammatory mediators on membrane-associated mucins expressed by human ocular surface epithelial cells. Exp. Eye Res. 2010, 90, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Ballard, P.L. Hormonal influences during fetal lung development. Ciba Found. Symp 1980, 78, 251–274. [Google Scholar]
- Liley, H.G.; White, R.T.; Benson, B.J.; Ballard, P.L. Glucocorticoids both stimulate and inhibit production of pulmonary surfactant protein A in fetal human lung. Proc. Natl. Acad. Sci. USA 1988, 85, 9096–9100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dausch, D.; Lee, S.; Dausch, S.; Kim, J.C.; Schwert, G.; Michelson, W. Comparative study of treatment of the dry eye syndrome due to disturbances of the tear film lipid layer with lipid-containing tear substitutes. Klin. Mon. Fur Augenheilkd. 2006, 223, 974–983. [Google Scholar] [CrossRef] [PubMed]
- Nosch, D.S.; Joos, R.E.; Job, M. Prospective randomized study to evaluate the efficacy and tolerability of Ectoin(R) containing Eye Spray (EES09) and comparison to the liposomal Eye Spray Tears Again(R) (TA) in the treatment of dry eye disease. Cont. Lens Anterior Eye 2021, 44, 101318. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving physical realism, stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins Struct. Funct. Bioinform. 2009, 77 (Suppl. S9), 114–122. [Google Scholar] [CrossRef] [Green Version]
- Cwiklik, L. Tear film lipid layer: A molecular level view. Biochim. Et Biophys. Acta 2016, 1858, 2421–2430. [Google Scholar] [CrossRef]
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci 1995, 36, 614–621. [Google Scholar]
- Diebold, Y.; Calonge, M.; Enriquez de Salamanca, A.; Callejo, S.; Corrales, R.M.; Saez, V.; Siemasko, K.F.; Stern, M.E. Characterization of a spontaneously immortalized cell line (IOBA-NHC) from normal human conjunctiva. Investig. Ophthalmol. Vis. Sci 2003, 44, 4263–4274. [Google Scholar] [CrossRef] [Green Version]
- Schicht, M.; Knipping, S.; Hirt, R.; Beileke, S.; Sel, S.; Paulsen, F.; Bräuer, L. Detection of surfactant proteins A, B, C, and D in human nasal mucosa and their regulation in chronic rhinosinusitis with polyps. Am. J. Rhinol. Allergy 2013, 27, 24–29. [Google Scholar] [CrossRef]
- Wizert, A.; Iskander, D.R.; Cwiklik, L. Organization of lipids in the tear film: A molecular-level view. PLoS ONE 2014, 9, e92461. [Google Scholar] [CrossRef] [Green Version]
- Wizert, A.; Iskander, D.R.; Cwiklik, L. Interaction of lysozyme with a tear film lipid layer model: A molecular dynamics simulation study. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2289–2296. [Google Scholar] [CrossRef]
- Marrink, S.J.; Tieleman, D.P. Perspective on the Martini model. Chem Soc. Rev. 2013, 42, 6801–6822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Points | Pairings | Statistical Significance |
|---|---|---|
| 12 h | sfm + TA vs. 100 ng SP-G + TA | *** |
| 12 h | sfm + Tris vs. 100 ng SP-G + Tris | *** |
| 24 h | sfm + TA vs. 10 ng SP-G + TA | *** |
| 24 h | sfm + TA vs. 100 ng SP-G + TA | *** |
| 24 h | sfm + Tris vs. 100 ng SP-G + Tris | ** |
| 48 h | sfm + TA vs. 10 ng SP-G + TA | ** |
| 48 h | sfm + TA vs. 100 ng SP-G + TA | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schicht, M.; Riedlová, K.; Kukulka, M.; Li, W.; Scheer, A.; Garreis, F.; Jacobi, C.; Paulsen, F.; Cwiklik, L.; Bräuer, L. The Potential Role of SP-G as Surface Tension Regulator in Tear Film: From Molecular Simulations to Experimental Observations. Int. J. Mol. Sci. 2022, 23, 5783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105783
Schicht M, Riedlová K, Kukulka M, Li W, Scheer A, Garreis F, Jacobi C, Paulsen F, Cwiklik L, Bräuer L. The Potential Role of SP-G as Surface Tension Regulator in Tear Film: From Molecular Simulations to Experimental Observations. International Journal of Molecular Sciences. 2022; 23(10):5783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105783
Chicago/Turabian StyleSchicht, Martin, Kamila Riedlová, Mercedes Kukulka, Wenyue Li, Aurelius Scheer, Fabian Garreis, Christina Jacobi, Friedrich Paulsen, Lukasz Cwiklik, and Lars Bräuer. 2022. "The Potential Role of SP-G as Surface Tension Regulator in Tear Film: From Molecular Simulations to Experimental Observations" International Journal of Molecular Sciences 23, no. 10: 5783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105783