
Recent Advances in Molecular Research on Hydrogen Sulfide (H2S) Role in Diabetes Mellitus (DM)—A Systematic Review

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
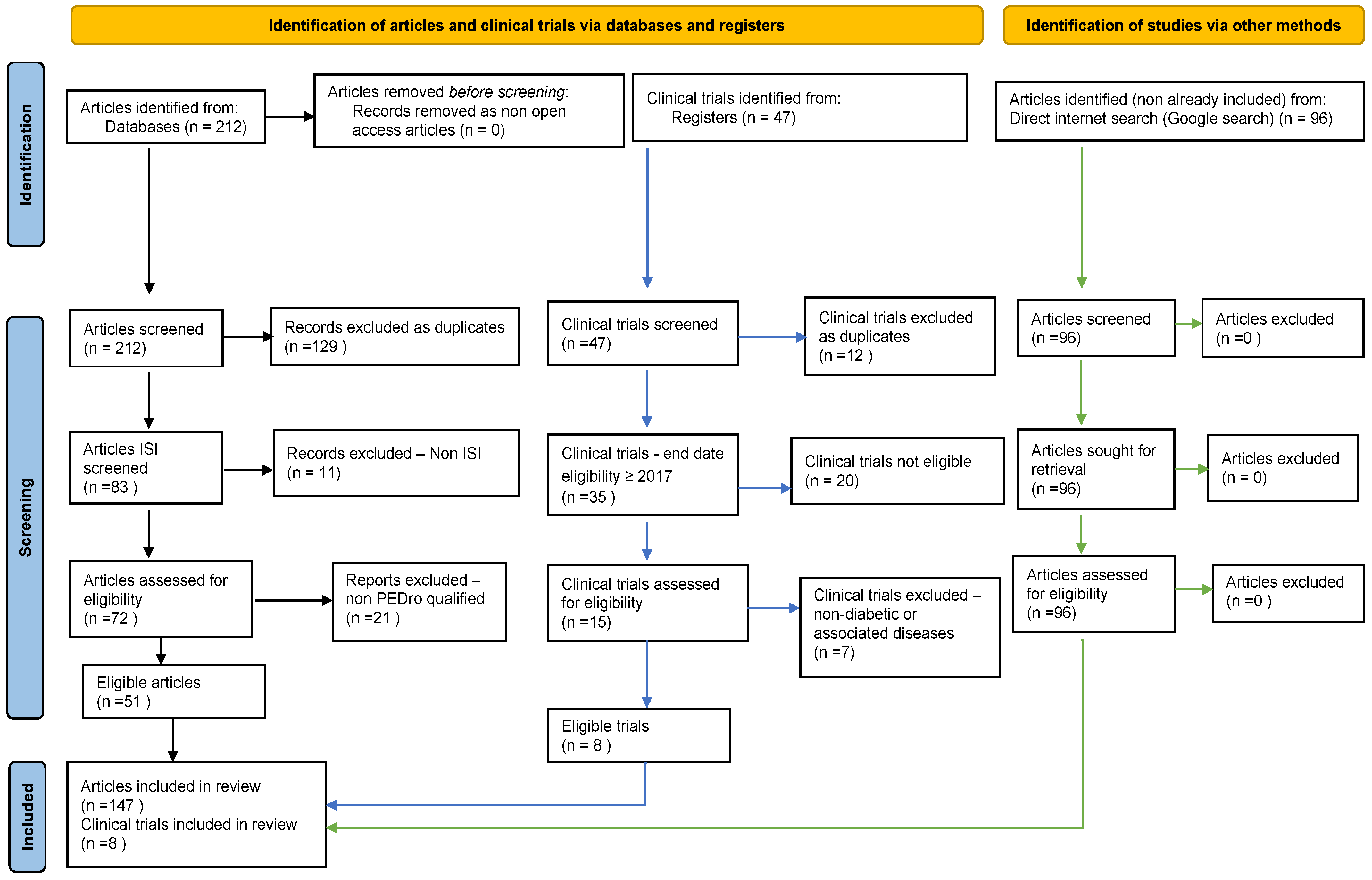
3.1. Search and Filtering Results
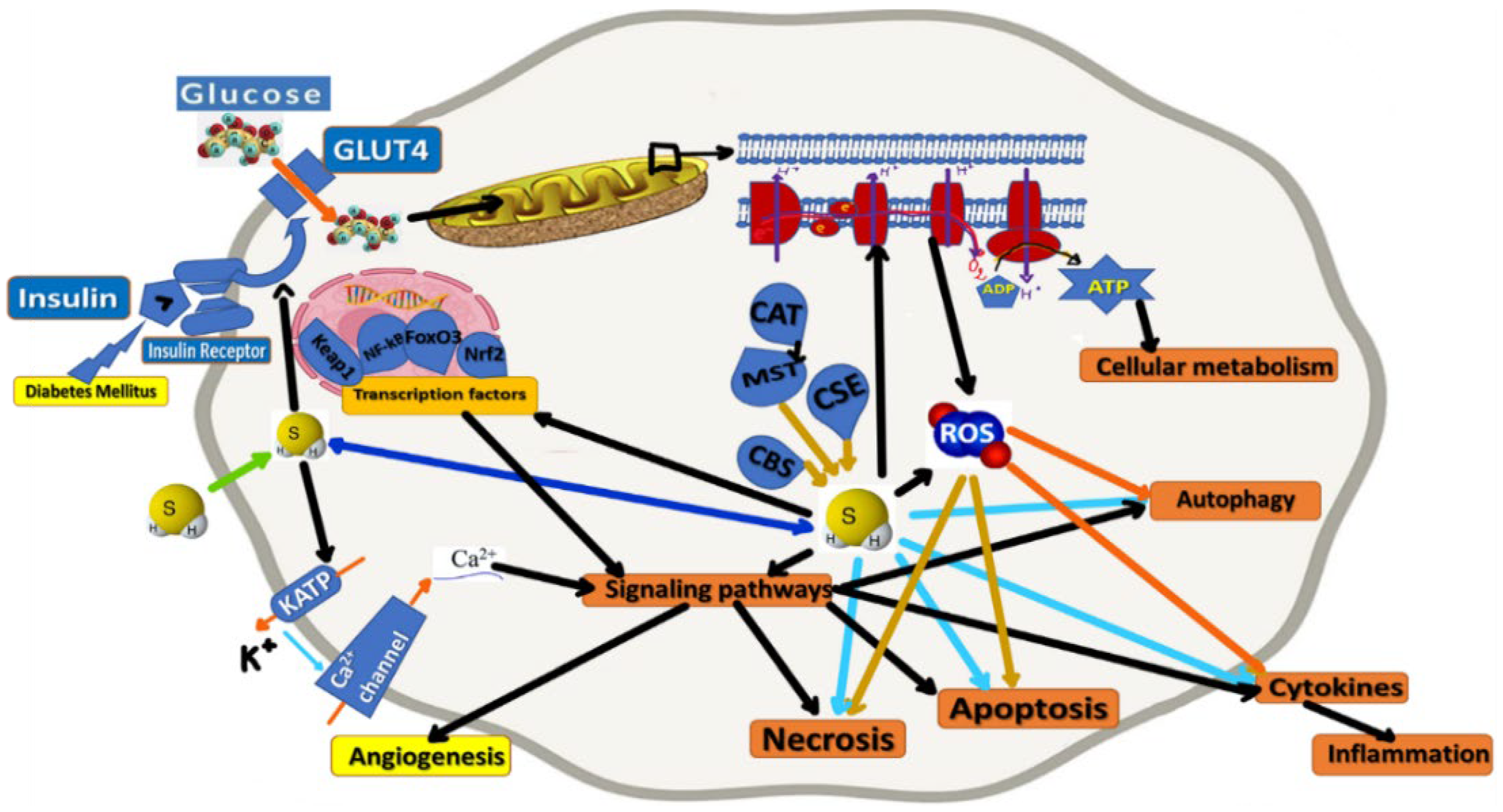
3.2. Physiological Properties of H2S
3.3. H2S in Pharmacology and Pathophysiology
3.4. H2S and Insulin Secretion and Sensitivity
3.5. H2S and Neurological Dysfunctions as Diabetes Associated Diseases
3.6. H2S and Cardio-Vascular Dysfunctions as Diabetes Associated Diseases
3.7. H2S and Renal Dysfunctions as Diabetes Associated Diseases
3.8. H2S Exogenous Sources as Possible Therapeutic Interventions in Diabetes or Related Diseases
3.9. Synthetic Slow-Releasing H2S Donors
3.10. H2S-Stimulating Agents
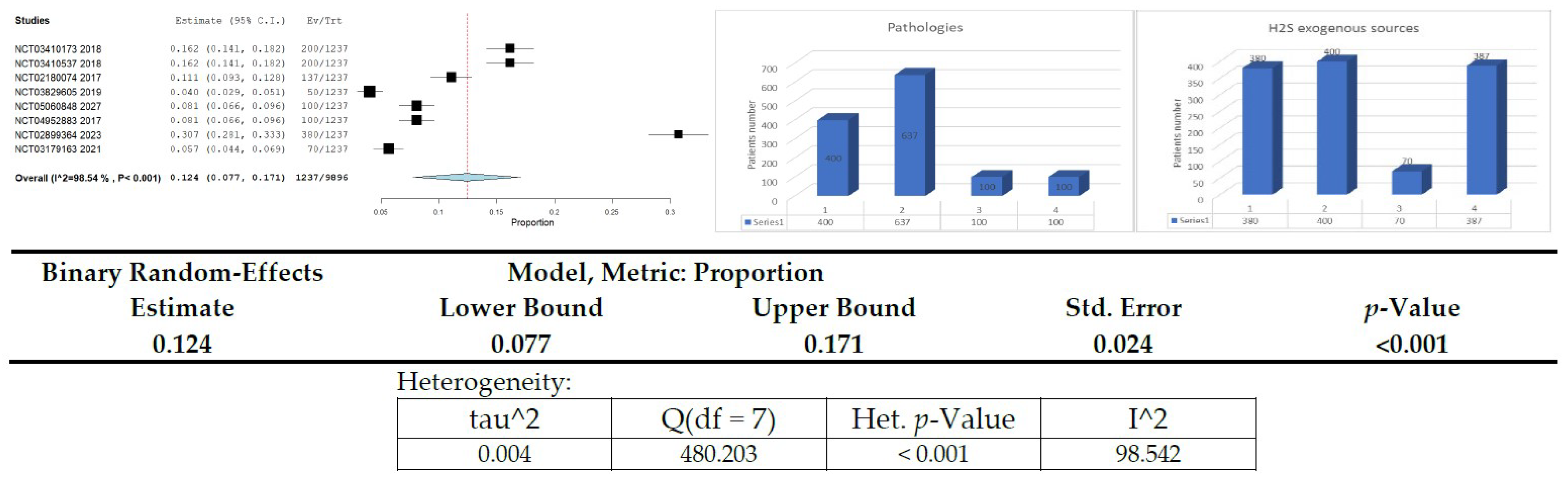
3.11. Clinical Studies on H2S Donors/Exogenous Sources in Diabetes or Related Diseases—Meta-Analysis
- ✓
- 17,498 Studies found for: Diabetes Mellitus
- ✓
- 2 Studies found for: H2S/Hydrogen sulfide|Diabetes Mellitus
- ✓
- 18 Studies found for: hydrogen sulfide
- ✓
- 6430 trials found: Diabetes Mellitus
- ✓
- 0 Studies found for: H2S/Hydrogen sulfide|Diabetes Mellitus
- ✓
- 25 trials found for: hydrogen sulfide
- ✓
- 2041 trials found: Diabetes Mellitus
- ✓
- 0 Studies found for: H2S/Hydrogen sulfide|Diabetes Mellitus
- ✓
- 1 trial found for: hydrogen sulfide
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munteanu, C.; Păun, D.-L.; Șuță, A.-M.; Florescu, S.A.; Onose, G. Diabetes mellitus and COVID-19 in the post-acute phase patients—possible links with physical and rehabilitation medicine and balneotherapy. Balneo Res. J. 2020, 11, 350–367. [Google Scholar] [CrossRef]
- Melino, S.; Leo, S.; Papajani, V.T. Natural Hydrogen Sulfide Donors from Allium sp. as a Nutraceutical Approach in Type 2 Diabetes Prevention and Therapy. Nutrients 2019, 11, 1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.-J.; Wu, Z.-Y.; Nie, X.-W.; Wang, X.-Y.; Bian, J.-S. An Updated Insight Into Molecular Mechanism of Hydrogen Sulfide in Cardiomyopathy and Myocardial Ischemia/Reperfusion Injury under Diabetes. Front. Pharmacol. 2021, 12, 651884. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.; Rotariu, M.; Dogaru, G.; Ionescu, E.V.; Ciobanu, V.; Onose, G. Mud therapy and rehabilitation—Scientific relevance in the last six years (2015–2020) Systematic literature review and meta-analysis based on the PRISMA paradigm. Balneo PRM Res. J. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Sashi Papu, J.A.; Kundu, S.; Pushpakumar, S.; Amin, M.; Tyagi, S.C.; Sen, U. Hydrogen sulfide inhibits Ca2+-induced mitochondrial permeability transition pore opening in type-1 diabetes. Am. J. Physiol.—Endocrinol. Metab. 2019, 317, E269–E283. [Google Scholar] [CrossRef] [PubMed]
- George, A.K.; Singh, M.; Homme, R.P.; Majumder, A.; Sandhu, H.S.; Tyagi, S.C. A hypothesis for treating inflammation and oxidative stress with hydrogen sulfide during age-related macular degeneration. Int. J. Ophthalmol. 2018, 11, 881–887. [Google Scholar] [CrossRef]
- Citi, V.; Martelli, A.; Gorica, E.; Brogi, S.; Testai, L.; Calderone, V. Role of hydrogen sulfide in endothelial dysfunction: Pathophysiology and therapeutic approaches. J. Adv. Res. 2021, 27, 99–113. [Google Scholar] [CrossRef]
- Gheibi, S.; Samsonov, A.P.; Gheibi, S.; Vazquez, A.B.; Kashfi, K. Regulation of carbohydrate metabolism by nitric oxide and hydrogen sulfide: Implications in diabetes. Biochem. Pharmacol. 2020, 176, 113819. [Google Scholar] [CrossRef]
- Stehouwer, C.D.; Lambert, J.; Donker, A.; Van Hinsbergh, V.W. Endothelial dysfunction and pathogenesis of diabetic angiopathy. Cardiovasc. Res. 1997, 34, 55–68. [Google Scholar] [CrossRef]
- Zou, W.; Yuan, J.; Tang, Z.-J.; Wei, H.-J.; Zhu, W.-W.; Zhang, P.; Gu, H.-F.; Wang, C.-Y.; Tang, X.-Q. Hydrogen sulfide ameliorates cognitive dysfunction in streptozotocin-induced diabetic rats: Involving suppression in hippocampal endoplasmic reticulum stress. Oncotarget 2017, 8, 64203–64216. [Google Scholar] [CrossRef] [Green Version]
- Rey, F.; Ottolenghi, S.; Giallongo, T.; Balsari, A.; Martinelli, C.; Rey, R.; Allevi, R.; Giulio, A.M.; Zuccotti, G.V.; Mazzucchelli, S.; et al. Mitochondrial metabolism as target of the neuroprotective role of erythropoietin in Parkinson’s disease. Antioxidants 2021, 10, 121. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Brancaleone, V.; Flori, L.; Montanaro, R.; Calderone, V. Modulation of EndMT by Hydrogen Sulfide in the Prevention of Cardiovascular Fibrosis. Antioxidants 2021, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Ciccone, V.; Genah, S.; Morbidelli, L. Endothelium as a Source and Target of H2S to Improve Its Trophism and Function. Antioxidants 2021, 10, 486. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.C.; Sohn, E.-H.; Lee, S.R. Hydrogen Sulfide as a Potential Alternative for the Treatment of Myocardial Fibrosis. Oxidative Med. Cell. Longev. 2020, 2020, 4105382. [Google Scholar] [CrossRef]
- Lee, Z.-W.; Teo, X.-Y.; Tay, E.Y.-W.; Tan, C.-H.; Hagen, T.; Moore, P.K.; Deng, L.-W. Utilizing hydrogen sulfide as a novel anti-cancer agent by targeting cancer glycolysis and pH imbalance. J. Cereb. Blood Flow Metab. 2014, 171, 4322–4336. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Tian, Z.; Sun, Y.; Lu, C.; Liu, N.; Gao, Z.; Zhang, L.; Dong, S.; Yang, F.; Zhong, X.; et al. Exogenous H2S facilitating ubiquitin aggregates clearance via autophagy attenuates type 2 diabetes-induced cardiomyopathy. Cell Death Dis. 2017, 8, e2992-11. [Google Scholar] [CrossRef] [Green Version]
- Munteanu, C.; Munteanu, D.; Onose, G. Hydrogen sulfide (H2S)—Therapeutic relevance in rehabilitation and balneotherapy Systematic literature review and meta-analysis based on the PRISMA paradig. Balneo PRM Res. J. 2021, 12, 176–195. [Google Scholar] [CrossRef]
- Ghashghaeinia, M.; Mrowietz, U. Human erythrocytes, nuclear factor kappaB (NFκB) and hydrogen sulfide (H2S)—From non-genomic to genomic research. Cell Cycle 2021, 20, 2091–2101. [Google Scholar] [CrossRef]
- Marunaka, Y. Roles of interstitial fluid pH in diabetes mellitus: Glycolysis and mitochondrial function. World J. Diabetes 2015, 6, 125. [Google Scholar] [CrossRef]
- Hu, B.; Xu, G.; Zheng, Y.; Tong, F.; Qian, P.; Pan, X.; Zhou, X.; Shen, R. Chelerythrine Attenuates Renal Ischemia/Reperfusion-induced Myocardial Injury by Activating CSE/H2S via PKC/NF-κB Pathway in Diabetic Rats. Kidney Blood Press. Res. 2017, 42, 379–388. [Google Scholar] [CrossRef]
- Yang, C.-T.; Chen, L.; Xu, S.; Day, J.J.; Li, X.; Xian, M. Recent Development of Hydrogen Sulfide Releasing/Stimulating Reagents and Their Potential Applications in Cancer and Glycometabolic Disorders. Front. Pharmacol. 2017, 8, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, S.; Kambis, T.N.; Mishra, P.K. Hydrogen sulfide-mediated regulation of cell death signaling ameliorates adverse cardiac remodeling and diabetic cardiomyopathy. Am. J. Physiol. Circ. Physiol. 2019, 316, H1237–H1252. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.; Dogaru, G.; Rotariu, M.; Onose, G. Therapeutic gases used in balneotherapy and rehabilitation medicine—Scientific relevance in the last ten years (2011–2020)—Synthetic literature review. Balneo PRM Res. J. 2021, 12, 111–122. [Google Scholar] [CrossRef]
- Jeong, N.Y.; Jung, J.; Tabassum, R. Protective effect of hydrogen sulfide on oxidative stress-induced neurodegenerative diseases. Neural Regen. Res. 2020, 15, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Hu, S.; Liu, Q.; Han, M.; Wang, F.; Qiu, M.; Li, S.; Li, X.; Yang, T.; Fu, X.; et al. Hydrogen sulfide upregulates renal AQP-2 protein expression and promotes urine concentration. FASEB J. 2018, 33, 469–483. [Google Scholar] [CrossRef]
- Yang, R.; Jia, Q.; Ma, S.; Wang, Y.; Mehmood, S.; Chen, Y. Exogenous H2S mitigates myocardial fibrosis in diabetic rats through suppression of the canonical Wnt pathway. Int. J. Mol. Med. 2019, 44, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, Y.; Liang, B.; Li, Z.; Jiang, Z.; Chu, C.; Yang, J. Hydrogen sulfide attenuates myocardial fibrosis in diabetic rats through the JAK/STAT signaling pathway. Int. J. Mol. Med. 2018, 41, 1867–1876. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Teng, Z.; Sun, X.; Zhang, L.; Chen, J.; Wang, B.; Lu, F.; Liu, N.; Yu, M.; Peng, S.; et al. Exogenous H2S reduces the acetylation levels of mitochondrial respiratory enzymes via regulating the NAD+-SIRT3 pathway in cardiac tissues of db/db mice. Am. J. Physiol. Metab. 2019, 317, E284–E297. [Google Scholar] [CrossRef]
- Roa-Coria, J.E.; Pineda-Farias, J.B.; Barragán-Iglesias, P.; Quiñonez-Bastidas, G.N.; Zúñiga-Romero, Á.; Huerta-Cruz, J.C.; Reyes-García, J.G.; Flores-Murrieta, F.J.; Granados-Soto, V.; Rocha-González, H.I. Possible involvement of peripheral TRP channels in the hydrogen sulfide-induced hyperalgesia in diabetic rats. BMC Neurosci. 2019, 20, 1. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Yu, X.; Li, T.; Wu, J.; Zhao, Y.; Liu, J.; Sun, A.; Dong, S.; Wu, J.; Zhong, X.; et al. Exogenous H2S regulates endoplasmic reticulum-mitochondria cross-talk to inhibit apoptotic pathways in STZ-induced type I diabetes. Am. J. Physiol.—Endocrinol. Metab. 2017, 312, E190–E203. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, X.; Li, X.; Wei, X.; Wang, H. Hydrogen Sulfide Plays an Important Role in Diabetic Cardiomyopathy. Front. Cell Dev. Biol. 2021, 9, 627336. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wu, J.; Zhang, L.; Gao, Z.; Sun, Y.; Yu, M.; Zhao, Y.; Dong, S.; Lu, F.; Zhang, W. Hydrogen Sulphide modulating mitochondrial morphology to promote mitophagy in endothelial cells under high-glucose and high-palmitate. J. Cell. Mol. Med. 2017, 21, 3190–3203. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Liu, K.; Xiao, L.; Jin, S.; Dong, J.; Teng, X.; Guo, Q.; Chen, Y.; Wu, Y. Alpha-lipoic acid regulates the autophagy of vascular smooth muscle cells in diabetes by elevating hydrogen sulfide level. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2018, 1864, 3723–3738. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xiao, T.; Li, F.; Li, Y.; Zeng, O.; Liu, M.; Liang, B.; Li, Z.; Chu, C.; Yang, J. Hydrogen sulfide reduced renal tissue fibrosis by regulating autophagy in diabetic rats. Mol. Med. Rep. 2017, 16, 1715–1722. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Du, H.; Wang, B.; Chen, J.; Lu, F.; Peng, S.; Sun, Y.; Liu, N.; Sun, X.; Shiyun, D.; et al. Exogenous H2S Induces Hrd1 S-sulfhydration and Prevents CD36 Translocation via VAMP3 Ubiquitylation in Diabetic Hearts. Aging Dis. 2020, 11, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Kar, S.; Shahshahan, H.R.; Kambis, T.N.; Yadav, S.K.; Li, Z.; Lefer, D.J.; Mishra, P.K. Hydrogen Sulfide Ameliorates Homocysteine-Induced Cardiac Remodeling and Dysfunction. Front. Physiol. 2019, 10, 598. [Google Scholar] [CrossRef]
- Dominic, P.; Ahmad, J.; Bhandari, R.; Pardue, S.; Solorzano, J.; Jaisingh, K.; Watts, M.; Bailey, S.R.; Orr, A.W.; Kevil, C.G.; et al. Decreased availability of nitric oxide and hydrogen sulfide is a hallmark of COVID-19. Redox Biol. 2021, 43, 101982. [Google Scholar] [CrossRef]
- Loiselle, J.J.; Yang, G.; Wu, L. Hydrogen sulfide and hepatic lipid metabolism—A critical pairing for liver health. J. Cereb. Blood Flow Metab. 2020, 177, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Zhong, D.; Ma, P.; Li, G.; Hua, W.; Sun, Y.; Liu, N.; Zhang, L.; Zhang, W. Exogenous hydrogen sulfide ameliorates diabetes-associated cognitive decline by regulating the mitochondria-mediated apoptotic pathway and IL-23/IL-17 expression in db/db mice. Cell Physiol. Biochem. 2017, 41, 1838–1850. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Yang, W.; Zhang, H.; Song, Z.; Liu, T.; Lv, X. Hydrogen Sulfide Ameliorates Lung Ischemia-Reperfusion Injury Through SIRT1 Signaling Pathway in Type 2 Diabetic Rats. Front. Physiol. 2020, 11, 596. [Google Scholar] [CrossRef]
- Wu, L.; Chen, Y.; Wang, C.-Y.; Tang, Y.-Y.; Huang, H.-L.; Kang, X.; Li, X.; Xie, Y.-R.; Tang, X.-Q. Hydrogen Sulfide Inhibits High Glucose-Induced Neuronal Senescence by Improving Autophagic Flux via up-regulation of SIRTFront. Mol. Neurosci. 2019, 12, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Wu, Y.; Chen, J.; Zhong, J.; Zeng, F.; Wu, S. A Turn-On Optoacoustic Probe for Imaging Metformin-Induced Upregulation of Hepatic Hydrogen Sulfide and Subsequent Liver Injury. Theranostics 2019, 9, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C.; Papapetropoulos, A. International Union of Basic and Clinical Pharmacology. CII: Pharmacological Modulation of H2S Levels: H2S Donors and H2S Biosynthesis Inhibitors. Pharmacol. Rev. 2017, 69, 497–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.-J.; Wu, Z.-Y.; Nie, X.-W.; Bian, J.-S. Role of Endothelial Dysfunction in Cardiovascular Diseases: The Link Between Inflammation and Hydrogen Sulfide. Front. Pharmacol. 2020, 10, 1568. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Pan, L.; Ji, Y. H 2S protects against diabetes-accelerated atherosclerosis by preventing the activation of NLRP3 inflammasome. J. Biomed. Res. 2019, 34, 94–102. [Google Scholar] [CrossRef]
- Jia, Q.; Mehmood, S.; Liu, X.; Ma, S.; Yang, R. Hydrogen sulfide mitigates myocardial inflammation by inhibiting nucleotide-binding oligomerization domain-like receptor protein 3 inflammasome activation in diabetic rats. Exp. Biol. Med. 2020, 245, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, L.; Zeng, O.; Liu, J.M.; Yang, J. H2S improves renal fibrosis in STZ-induced diabetic rats by ameliorating TGF-β1 expression. Ren. Fail. 2016, 39, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Kar, S.; Shahshahan, H.R.; Hackfort, B.T.; Yadav, S.K.; Yadav, R.; Kambis, T.N.; Lefer, D.J.; Mishra, P.K. Exercise Training Promotes Cardiac Hydrogen Sulfide Biosynthesis and Mitigates Pyroptosis to Prevent High-Fat Diet-Induced Diabetic Cardiomyopathy. Antioxidants 2019, 11, 638. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yuan, Y.; Zhang, L.; Zhang, H.; Zhang, S.; Zhang, Y.; Xuan, X.-X.; Wang, M.-J.; Zhang, J.-Y. Exogenous hydrogen sulfide protects against high glucose-induced apoptosis and oxidative stress by inhibiting the STAT3/HIF-1α pathway in H9c2 cardiomyocytes. Exp. Ther. Med. 2019, 18, 3948–3958. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Huang, Y.; Chen, S.; Tang, C.; Wang, G.; Du, J.; Jin, H. Hydrogen sulfide regulates insulin secretion and insulin resistance in diabetes mellitus, a new promising target for diabetes mellitus treatment? A review. J. Adv. Res. 2021, 27, 19–30. [Google Scholar] [CrossRef]
- Chen, H.-J.; Ngowi, E.E.; Qian, L.; Li, T.; Qin, Y.-Z.; Zhou, J.-J.; Li, K.; Ji, X.-Y.; Wu, D.-D. Role of Hydrogen Sulfide in the Endocrine System. Front. Endocrinol. 2021, 12, 704620. [Google Scholar] [CrossRef] [PubMed]
- Gheibi, S.; Jeddi, S.; Kashfi, K.; Ghasemi, A. Effects of Hydrogen Sulfide on Carbohydrate Metabolism in Obese Type 2 Diabetic Rats. Molecules 2019, 24, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.-L.; Ren, J.-D.; Huang, Z.; Wang, T.; Xiang, K.; Cheng, L.; Tang, L.-J. The Role of Exogenous Hydrogen Sulfide in Free Fatty Acids Induced Inflammation in Macrophages. Cell. Physiol. Biochem. 2017, 42, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Comas, F.; Moreno-Navarrete, J.M. The Impact of H2S on Obesity-Associated Metabolic Disturbances. Antioxidants 2021, 10, 633. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Sagara, M.; Aoki, C.; Tanaka, S.; Aso, Y. Clinical Implication of Plasma Hydrogen Sulfide Levels in Japanese Patients with Type 2 Diabetes. Intern. Med. 2017, 56, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.B.; Zhou, H.; Li, L.; Kang, Y.; Cao, X.; Wu, Z.Y.; Ding, L.; Sethi, G.; Bian, J.-S. Hydrogen sulfide prevents elastin loss and attenuates calcification induced by high glucose in smooth muscle cells through suppression of stat3/cathepsin s signaling pathway. Int. J. Mol. Sci. 2019, 20, 4202. [Google Scholar] [CrossRef] [Green Version]
- John, A.M.S.P.; Kundu, S.; Pushpakumar, S.; Fordham, M.; Weber, G.; Mukhopadhyay, M.; Sen, U. GYY4137, a Hydrogen Sulfide Donor Modulates miR194-Dependent Collagen Realignment in Diabetic Kidney. Sci. Rep. 2017, 7, 879. [Google Scholar] [CrossRef]
- Bitar, M.S.; Nader, J.; Al-Ali, W.; Al Madhoun, A.; Arefanian, H.; Al-Mulla, F. Hydrogen Sulfide Donor NaHS Improves Metabolism and Reduces Muscle Atrophy in Type 2 Diabetes: Implication for Understanding Sarcopenic Pathophysiology. Oxidative Med. Cell. Longev. 2018, 2018, 6825452. [Google Scholar] [CrossRef]
- Ding, T.; Chen, W.; Li, J.; Ding, J.; Mei, X.; Hu, H. High Glucose Induces Mouse Mesangial Cell Overproliferation via Inhibition of Hydrogen Sulfide Synthesis in a TLR-4-Dependent Manner. Cell. Physiol. Biochem. 2017, 41, 1035–1043. [Google Scholar] [CrossRef]
- Lia, Q.; Lancaster, J.R. Chemical Foundations of Hydrogen Sulfide Biology. Nitric Oxide. 2013. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC3624763/pdf/nihms412728.pdf (accessed on 20 May 2022).
- Cheng, Z.; Kishore, R. Potential role of hydrogen sulfide in diabetes-impaired angiogenesis and ischemic tissue repair. Redox Biol. 2020, 37, 101704. [Google Scholar] [CrossRef]
- Roorda, M.; Miljkovic, J.L.; Van Goor, H.; Henning, R.H.; Bouma, H.R. Spatiotemporal regulation of hydrogen sulfide signaling in the kidney. Redox Biol. 2021, 43, 101961. [Google Scholar] [CrossRef] [PubMed]
- Bulboaca, A.E.; Boarescu, P.-M.; Porfire, A.S.; Dogaru, G.; Barbalata, C.; Valeanu, M.; Munteanu, C.; Râjnoveanu, R.M.; Nicula, C.A.; Stanescu, I.C. The Effect of Nano-Epigallocatechin-Gallate on Oxidative Stress and Matrix Metalloproteinases in Experimental Diabetes Mellitus. Antioxidants 2020, 9, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Shang, Q.; Yao, J.; Ji, Y. Hydrogen sulfide: A gaseous signaling molecule modulates tissue homeostasis: Implications in ophthalmic diseases. Cell Death Dis. 2019, 10, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, J.; Bhavsar, P.; Zhang, Q.; Hui, C.; Menzies-Gow, A.; Fan Chung, K. Sputum and serum hydrogen sulfide (H2S) as novel biomarker of asthma. Clin. Transl. Allergy 2013, 3. Available online: http://www.ctajournal.com/content/3/S1/P3 (accessed on 20 May 2022). [CrossRef] [Green Version]
- Lu, M.; Zhao, F.-F.; Tang, J.-J.; Su, C.-J.; Fan, Y.; Ding, J.-H.; Bian, J.; Hu, G. The Neuroprotection of Hydrogen Sulfide Against MPTP-Induced Dopaminergic Neuron Degeneration Involves Uncoupling Protein 2 Rather Than ATP-Sensitive Potassium Channels. Antioxid. Redox Signal. 2012, 17, 849–859. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Asahina, R.; Fujioka, M.; Matsui, T.K.; Kato, S.; Mori, E.; Hioki, H.; Yamamoto, T.; Kobayashi, K.; Tsuboi, A. Ras-like Gem GTPase induced by Npas4 promotes activity-dependent neuronal tolerance for ischemic stroke. Proc. Natl. Acad. Sci. USA 2021, 118, e2018850118. [Google Scholar] [CrossRef]
- Vela-Anero, Á.; Hermida-Gómez, T.; Gato-Calvo, L.; Vaamonde-García, C.; Díaz-Prado, S.; Meijide-Faílde, R.; Blanco, F.; Burguera, E. Long-term effects of hydrogen sulfide on the anabolic-catabolic balance of articular cartilage in vitro. Nitric Oxide 2017, 70, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, A.; Kurian, G.A. Hydrogen Sulfide Promotes Proliferation of HT-29 Colon Cancer Cells in a Mitochondria-independent Pathway. Indian J. Pharm. Sci. 2019, 81, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Tang, G.; Zhang, L.; Wu, L.; Wang, R. The Pathogenic Role of Cystathionine γ-Lyase/Hydrogen Sulfide in Streptozotocin-Induced Diabetes in Mice. Am. J. Pathol. 2011, 179, 869–879. [Google Scholar] [CrossRef]
- Xue, R.; Hao, D.-D.; Sun, J.-P.; Li, W.-W.; Zhao, M.-M.; Li, X.-H.; Chen, Y.; Zhu, J.-H.; Ding, Y.-J.; Liu, J.; et al. Hydrogen Sulfide Treatment Promotes Glucose Uptake by Increasing Insulin Receptor Sensitivity and Ameliorates Kidney Lesions in Type 2 Diabetes. Antioxid. Redox Signal. 2013, 19, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Ngowi, E.E.; Afzal, A.; Sarfraz, M.; Khattak, S.; Zaman, S.U.; Khan, N.H.; Li, T.; Jiang, Q.-Y.; Zhang, X.; Duan, S.-F.; et al. Role of hydrogen sulfide donors in cancer development and progression. Int. J. Biol. Sci. 2020, 17, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Potenza, M.A.; Sgarra, L.; Desantis, V.; Nacci, C.; Montagnani, M. Diabetes and alzheimer’s disease: Might mitochondrial dysfunction help deciphering the common path? Antioxidants 2021, 10, 1257. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C. Hydrogen Sulfide, an Endogenous Stimulator of Mitochondrial Function in Cancer Cells. Cells 2021, 10, 220. [Google Scholar] [CrossRef]
- Monteiro, B.S.; Freire-Brito, L.; Carrageta, D.F.; Oliveira, P.F.; Alves, M.G. Mitochondrial Uncoupling Proteins (UCPs) as Key Modulators of ROS Homeostasis: A Crosstalk between Diabesity and Male Infertility? Antioxidants 2021, 10, 1746. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, R.; Jeong, N.Y. Potential for therapeutic use of hydrogen sulfide in oxidative stress-induced neurodegenerative diseases. Int. J. Med. Sci. 2019, 16, 1386–1396. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Yan, Q.; Liu, X.; Li, P.; Li, X.; Chen, Y.; Simoncini, T.; Liu, J.; Zhu, D.; Fu, X. 17β-Estradiol nongenomically induces vascular endothelial H2S release by promoting phosphorylation of cystathionine γ-lyase. J. Biol. Chem. 2019, 294, 15577–15592. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, D.; Gao, X.; Lew, K.; Richards, A.M.; Wang, P. mTORC2 Phosphorylation of Akt1: A Possible Mechanism for Hydrogen Sulfide-Induced Cardioprotection. PLoS ONE 2014, 9, e99665. [Google Scholar] [CrossRef] [Green Version]
- Lee, Z.-W.; Teo, X.-Y.; Song, Z.J.; Nin, D.S.; Novera, W.; Choo, B.A.; Dymock, B.W.; Moore, P.K.; Huang, R.Y.-J.; Deng, L.-W. Intracellular Hyper-Acidification Potentiated by Hydrogen Sulfide Mediates Invasive and Therapy Resistant Cancer Cell Death. Front. Pharmacol. 2017, 8, 763. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Mao, Y.-G.; Chen, X.; Zhang, Y. Hydrogen sulfide therapy: A narrative overview of current research and possible therapeutic implications in future. Med. Gas Res. 2020, 10, 185–188. [Google Scholar] [CrossRef]
- Chen, G.; Dou, Y.; Wang, Z. The role of hydrogen sulfide in stroke. Med. Gas Res. 2016, 6, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Huang, L. Molecular hydrogen: A therapeutic antioxidant and beyond. Med. Gas Res. 2016, 6, 219–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.-S.; Song, H.; Kim, E.-H.; Choi, J.-H.; Hong, H.; Han, Y.-M.; Hahm, K.-B. Inhibition of Hydrogen Sulfide-induced Angiogenesis and Inflammation in Vascular Endothelial Cells: Potential Mechanisms of Gastric Cancer Prevention by Korean Red Ginseng. J. Ginseng. Res. 2012, 36, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, B.D.; Snyder, S.H. H2S signalling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Stier-Jarmer, M.; Frisch, D.; Oberhauser, C.; Immich, G.; Kirschneck, M.; Schuh, A. Effects of single moor baths on physiological stress response and psychological state: A pilot study. Int. J. Biometeorol. 2017, 61, 1957–1964. [Google Scholar] [CrossRef]
- Zhan, Y.; Li, M.Z.; Yang, L.; Feng, X.F.; Zhang, Q.X.; Zhang, N.; Zhao, Y.-Y.; Zhao, H. An MRI study of neurovascular restorative after combination treatment with xiaoshuanenteric-coated capsule and enriched environment in rats after stroke. Front. Neurosci. 2019, 13, 701. [Google Scholar] [CrossRef]
- Streeter, E.Y.; Badoer, E.; Woodman, O.L.; Hart, J.L. Effect of type 1 diabetes on the production and vasoactivity of hydrogen sulfide in rat middle cerebral arteries. Physiol. Rep. 2013, 1, e00111. [Google Scholar] [CrossRef]
- Allen, C.L.; Wolanska, K.; Malhi, N.K.; Benest, A.V.; Wood, M.E.; Amoaku, W.; Torregrossa, R.; Whiteman, M.; Bates, D.O.; Whatmore, J.L. Hydrogen Sulfide Is a Novel Protector of the Retinal Glycocalyx and Endothelial Permeability Barrier. Front. Cell Dev. Biol. 2021, 9, 724905. [Google Scholar] [CrossRef]
- Liu, Z.; Ran, Y.; Qie, S.; Gong, W.; Gao, F.; Ding, Z.; Xi, J. Melatonin protects against ischemic stroke by modulating microglia/macrophage polarization toward anti-inflammatory phenotype through STAT3 pathway. CNS Neurosci. Ther. 2019, 25, 1353–1362. [Google Scholar] [CrossRef] [Green Version]
- Predmore, B.L.; Lefer, D.J.; Gojon, G. Hydrogen Sulfide in Biochemistry and Medicine. Antioxid. Redox Signal. 2012, 17, 119–140. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, S.; Minamino, S.; Takase, M.; Ishiyama, Y.; Hosokura, H.; Kohda, T.; Ikeda, Y.; Fujimoto, Y. Hydrogen sulfide donor GYY4137 suppresses proliferation of human colorectal cancer Caco-2 cells by inducing both cell cycle arrest and cell death. Heliyon 2019, 5, e02244. [Google Scholar] [CrossRef] [Green Version]
- Narne, P.; Pandey, V.; Phanithi, P.B. Role of Nitric Oxide and Hydrogen Sulfide in Ischemic Stroke and the Emergent Epigenetic Underpinnings. Mol. Neurobiol. 2019, 56, 1749–1769. [Google Scholar] [CrossRef] [PubMed]
- Dilek, N.; Papapetropoulos, A.; Toliver-Kinsky, T.; Szabo, C. Hydrogen sulfide: An endogenous regulator of the immune system. Pharmacol. Res. 2020, 161, 105119. [Google Scholar] [CrossRef] [PubMed]
- Ortega, E.; Gálvez, I.; Hinchado, M.D.; Guerrero, J.; Martín-Cordero, L.; Torres-Piles, S. Anti-inflammatory effect as a mechanism of effectiveness underlying the clinical benefits of pelotherapy in osteoarthritis patients: Regulation of the altered inflammatory and stress feedback response. Int. J. Biometeorol. 2017, 61, 1777–1785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.; Zhao, G.; Sun, A.; Zong, N.C.; Li, Z.; Zhu, H.; Zou, Y.; Yang, X.; Ge, J. Hydrogen Sulfide Attenuates the Recruitment of CD11b+Gr-1+ Myeloid Cells and Regulates Bax/Bcl-2 Signaling in Myocardial Ischemia Injury. Sci. Rep. 2014, 4, 4774. [Google Scholar] [CrossRef]
- Merighi, S.; Gessi, S.; Varani, K.; Fazzi, D.; Borea, P.A. Hydrogen sulfide modulates the release of nitric oxide and VEGF in human keratinocytes. Pharmacol. Res. 2012, 66, 428–436. [Google Scholar] [CrossRef]
- Wu, L.; Yang, W.; Jia, X.; Yang, G.; Duridanova, D.; Cao, K.; Wang, R. Pancreatic islet overproduction of H2S and suppressed insulin release in Zucker diabetic rats. Lab. Investig. 2009, 89, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.; Ling, X.; Xi, W.; Wang, P.; Sun, J.; Yang, Q.; Xiao, J. Exogenous hydrogen sulfide reduces atrial remodeling and atrial fibrillation induced by diabetes mellitus via activation of the PI3K/Akt/eNOS pathway. Mol. Med. Rep. 2020, 22, 1759–1766. [Google Scholar] [CrossRef]
- Malagrinò, F.; Zuhra, K.; Mascolo, L.; Mastronicola, D.; Vicente, J.B.; Forte, E.; Giuffrè, A. Hydrogen Sulfide Oxidation: Adaptive Changes in Mitochondria of SW480 Colorectal Cancer Cells upon Exposure to Hypoxia. Oxidative Med. Cell. Longev. 2019, 2019, 8102936. [Google Scholar] [CrossRef] [Green Version]
- Giuffrè, A.; Vicente, J.B. Hydrogen Sulfide Biochemistry and Interplay with Other Gaseous Mediators in Mammalian Physiology. Oxidative Med. Cell. Longev. 2018, 2018, 6290931. [Google Scholar] [CrossRef]
- Li, P.; Liu, H.; Shi, X.; Prokosch, V. Hydrogen sulfide: Novel endogenous and exogenous modulator of oxidative stress in retinal degeneration diseases. Molecules 2021, 26, 2411. [Google Scholar] [CrossRef]
- Li, H.; Xu, F.; Gao, G.; Gao, X.; Wu, B.; Zheng, C.; Wang, P.; Li, Z.; Hua, H.; Li, D. Hydrogen sulfide and its donors: Novel antitumor and antimetastatic therapies for triple-negative breast cancer. Redox Biol. 2020, 34, 101564. [Google Scholar] [CrossRef] [PubMed]
- Hellmich, M.R.; Szabo, C. Hydrogen Sulfide and Cancer. Handb. Exp. Pharmacol. 2015, 230, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, L.; Shen, J.; Chuai, Y.; Cai, J. Hydrogen as a New Class of Radioprotective Agent. Int. J. Biol. Sci. 2013, 9, 887–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.-H.; Ho, J.Y.; Fan, L.; Richardson, D.J.; Yuan, Z.; Bond, P.L. Antimicrobial Effects of Free Nitrous Acid on Desulfovibrio vulgaris: Implications for Sulfide-Induced Corrosion of Concrete. Appl. Environ. Microbiol. 2016, 82, 5563–5575. [Google Scholar] [CrossRef] [Green Version]
- Shan, H.; Qiu, J.; Chang, P.; Chu, Y.; Gao, C.; Wang, H.; Chen, G.; Luo, C.; Wang, T.; Chen, X.; et al. Exogenous Hydrogen Sulfide Offers Neuroprotection on Intracerebral Hemorrhage Injury Through Modulating Endogenous H2S Metabolism in Mice. Front. Cell. Neurosci. 2019, 13, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scammahorn, J.J.; Nguyen, I.T.N.; Bos, E.M.; Van Goor, H.; Joles, J.A. Fighting Oxidative Stress with Sulfur: Hydrogen Sulfide in the Renal and Cardiovascular Systems. Antioxidants 2021, 10, 373. [Google Scholar] [CrossRef]
- Xie, L.; Gu, Y.; Wen, M.; Zhao, S.; Wang, W.; Ma, Y.; Meng, G.; Han, Y.; Wang, Y.; Liu, G.; et al. Hydrogen Sulfide Induces Keap1 S-sulfhydration and Suppresses Diabetes-Accelerated Atherosclerosis via Nrf2 Activation. Diabetes 2016, 65, 3171–3184. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Luo, N.; Wang, L.; Zhao, Z.; Bu, H.; Xu, G.; Yan, Y.; Che, X.; Jiao, Z.; Zhao, T.; et al. Hydrogen sulfide ameliorates chronic renal failure in rats by inhibiting apoptosis and inflammation through ROS/MAPK and NF-κB signaling pathways. Sci. Rep. 2017, 7, 455. [Google Scholar] [CrossRef]
- Ji, J.; Xiang, P.; Li, T.; Lan, L.; Xu, X.; Lu, G.; Ji, H.; Zhang, Y.; Li, Y. NOSH-NBP, a Novel Nitric Oxide and Hydrogen Sulfide- Releasing Hybrid, Attenuates Ischemic Stroke-Induced Neuroinflammatory Injury by Modulating Microglia Polarization. Front. Cell. Neurosci. 2017, 11, 154. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, C.; Sun, W.; Li, L.; Wu, B.; Bai, S.; Li, H.; Zhong, X.; Wang, R.; Wu, L.; et al. Exogenous hydrogen sulfide restores cardioprotection of ischemic post-conditioning via inhibition of mPTP opening in the aging cardiomyocytes. Cell Biosci. 2015, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Xuan, A.; Long, D.; Li, J.; Ji, W.; Zhang, M.; Hong, L.; Liu, J. Hydrogen sulfide attenuates spatial memory impairment and hippocampal neuroinflammation in beta-amyloid rat model of Alzheimer’s disease. J. Neuroinflamm. 2012, 9, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.-Y.; Shen, Z.; He, P.; Jiang, L.; Hou, W.-W.; Shen, Y.; Zhang, X.-N.; Chen, Z. A Novel Neuroprotective Strategy for Ischemic Stroke: Transient Mild Acidosis Treatment by CO2 Inhalation at Reperfusion. J. Cereb. Blood Flow Metab. 2014, 34, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bełtowski, J.; Wójcicka, G.; Jamroz-Wiśniewska, A. Hydrogen sulfide in the regulation of insulin secretion and insulin sensitivity: Implications for the pathogenesis and treatment of diabetes mellitus. Biochem. Pharmacol. 2018, 149, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, P.; Ravindran, S.; Boovarahan, S.R.; Kurian, G.A. Hydrogen sulfide-mediated cardioprotection against ischemia reperfusion is linked to KATP channel for mitochondrial preservation but not for its distinct preference on interfibrillar mitochondria. Bangladesh J. Pharmacol. 2019, 14, 107–115. [Google Scholar] [CrossRef]
- Kamat, P.K.; Kyles, P.; Kalani, A.; Tyagi, N. Hydrogen Sulfide Ameliorates Homocysteine-Induced Alzheimer’s Disease-Like Pathology, Blood–Brain Barrier Disruption, and Synaptic Disorder. Mol. Neurobiol. 2016, 53, 2451–2467. [Google Scholar] [CrossRef] [PubMed]
- Borlongan, C.V.; Gonzales-Portillo, B.; Lippert, T.; Nguyen, H.; Lee, J.-Y. Hyperbaric oxygen therapy: A new look on treating stroke and traumatic brain injury. Brain Circ. 2019, 5, 101–105. [Google Scholar] [CrossRef]
- Durante, W. Hydrogen Sulfide Therapy in Diabetes-Accelerated Atherosclerosis: A Whiff of Success. Diabetes 2016, 65, 2832–2834. [Google Scholar] [CrossRef] [Green Version]
- Bredthauer, A.; Lehle, K.; Scheuerle, A.; Schelzig, H.; McCook, O.; Radermacher, P.; Szabo, C.; Wepler, M.; Simon, F. Intravenous hydrogen sulfide does not induce neuroprotection after aortic balloon occlusion-induced spinal cord ischemia/reperfusion injury in a human-like porcine model of ubiquitous arteriosclerosis. Intensiv. Care Med. Exp. 2018, 6, 44. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Shen, X.; Jiang, X.; Shan, H.; Cimini, M.; Fang, P.; Ji, Y.; Park, J.Y.; Drosatos, K.; Yang, X.; et al. Hyperhomocysteinemia potentiates diabetes-impaired EDHF-induced vascular relaxation: Role of insufficient hydrogen sulfide. Redox Biol. 2018, 16, 215–225. [Google Scholar] [CrossRef]
- Wang, R. Physiological Implications of Hydrogen Sulfide: A Whiff Exploration That Blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [Green Version]
- Lambooy, S.P.H.; Bidadkosh, A.; Nakladal, D.; Van Buiten, A.; Girgis, R.A.T.; Van Der Graaf, A.C.; Wiedenmann, T.J.; Koster, R.A.; Vogelaar, P.; Buikema, H.; et al. The Novel Compound Sul-121 Preserves Endothelial Function and Inhibits Progression of Kidney Damage in Type 2 Diabetes Mellitus in Mice. Sci. Rep. 2017, 7, 11165. [Google Scholar] [CrossRef] [PubMed]
- Dogaru, G.; Bulboaca, A.; Boarescu, P.M.; Ciumarnean, L.; Rus, V.; Sitar-Taut, A.-V.; Munteanu, C.; Bodisz, G.; Stanescu, I. The Effect of Mofettes on Oxidative Stress/Antioxidant Balance in Experimental Myocardial Ischemia. In Vivo 2019, 33, 1911–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambari, L.; Grigolo, B.; Filardo, G.; Grassi, F. Sulfurous thermal waters stimulate the osteogenic differentiation of human mesenchymal stromal cells—An in vitro study. Biomed. Pharmacother. 2020, 129, 110344. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, Y.; Wang, Q.; Wang, Z.; Cui, L.; Xu, Y.; Guan, K. Levels of nasal exhaled hydrogen sulfide in the general population and allergic rhinitis patients. J. Clin. Lab. Anal. 2021, 35, e23678. [Google Scholar] [CrossRef] [PubMed]
- Carubbi, C.; Gobbi, G.; Bucci, G.; Gesi, M.; Vitale, M.; Mirandola, P. Skin, Inflammation and Sulfurous Waters: What is Known, What is Believed. Eur. J. Inflamm. 2013, 11, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Hoteteu, M.; Romanian Association of Balneology; Munteanu, C.; Ionescu, E.V.; Almășan, R.E. Techirghiol Balnear and Rehabilitation Sanatorium Bioactive substances of the Techirghiol therapeutic mud. Balneo Res. J. 2018, 9, 5–10. [Google Scholar] [CrossRef]
- Carbajo, J.M.; Maraver, F. Sulphurous Mineral Waters: New Applications for Health. Evidence-Based Complement. Altern. Med. 2017, 2017, 8034084. [Google Scholar] [CrossRef]
- Viegas, J.; Esteves, A.F.; Cardoso, E.M.; Arosa, F.A.; Vitale, M.; Taborda-Barata, L. Biological Effects of Thermal Water-Associated Hydrogen Sulfide on Human Airways and Associated Immune Cells: Implications for Respiratory Diseases. Front. Public Health 2019, 7, 128. [Google Scholar] [CrossRef] [Green Version]
- Kida, K.; Yamada, M.; Tokuda, K.; Marutani, E.; Kakinohana, M.; Kaneki, M.; Ichinose, F. Inhaled Hydrogen Sulfide Prevents Neurodegeneration and Movement Disorder in a Mouse Model of Parkinson’s Disease. Antioxid. Redox Signal. 2011, 15, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Kloesch, B.; Liszt, M.; Krehan, D.; Broell, J.; Kiener, H.; Steiner, G. High concentrations of hydrogen sulphide elevate the expression of a series of pro-inflammatory genes in fibroblast-like synoviocytes derived from rheumatoid and osteoarthritis patients. Immunol. Lett. 2011, 141, 197–203. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Li, Y.; Li, L.; Xu, S.; Feng, X.; Liu, S. Hydrogen Sulfide (H2S)-Releasing Compounds: Therapeutic Potential in Cardiovascular Diseases. Front. Pharmacol. 2018, 9, 1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanderson, T.H.; Wider, J.M.; Lee, I.; Reynolds, C.A.; Liu, J.; Lepore, B.; Tousignant, R.; Bukowski, M.J.; Johnston, H.; Fite, A.; et al. Inhibitory modulation of cytochrome c oxidase activity with specific near-infrared light wavelengths attenuates brain ischemia/reperfusion injury. Sci. Rep. 2018, 8, 3481. [Google Scholar] [CrossRef] [PubMed]
- Mirandola, P.; Gobbi, G.; Micheloni, C.; Vaccarezza, M.; Di Marcantonio, D.; Ruscitti, F.; De Panfilis, G.; Vitale, M. Hydrogen sulfide inhibits IL-8 expression in human keratinocytes via MAP kinase signaling. Lab. Investig. 2011, 91, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Kashfi, K.; Chattopadhyay, M.; Kodela, R. NOSH-sulindac (AVT-18A) is a novel nitric oxide- and hydrogen sulfide-releasing hybrid that is gastrointestinal safe and has potent anti-inflammatory, analgesic, antipyretic, anti-platelet, and anti-cancer properties. Redox Biol. 2015, 6, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological Profile for Hydrogen Sulfide and Carbonyl Sulfide; U.S. Department of Health and Human Services, Public Health Service: Atlanta, GA, USA, 2016. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp114.pdf (accessed on 20 May 2022).
- Jia, J.; Xiao, Y.; Wang, W.; Qing, L.; Xu, Y.; Song, H.; Zhen, X.; Ao, G.; Alkayed, N.J.; Cheng, J. Differential mechanisms underlying neuroprotection of hydrogen sulfide donors against oxidative stress. Neurochem. Int. 2013, 62, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Diao, J.; Yi, H.; Xu, L.; Xu, J.; Xu, W. Signaling pathways involved in HSP32 induction by hyperbaric oxygen in rat spinal neurons. Redox Biol. 2016, 10, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Wu, L.; Montaut, S.; Yang, G. Hydrogen Sulfide Signaling Axis as a Target for Prostate Cancer Therapeutics. Prostate Cancer 2016, 2016, 8108549. [Google Scholar] [CrossRef] [Green Version]
- Stoica, S.I.; Bleotu, C.; Ciobanu, V.; Ionescu, A.M.; Albadi, I.; Onose, G.; Munteanu, C. Considerations about Hypoxic Changes in Neuraxis Tissue Injuries and Recovery. Biomedicines 2022, 10, 481. [Google Scholar] [CrossRef]
- Onose, G.; Anghelescu, A.; Blendea, D.; Ciobanu, V.; Daia, C.; Firan, F.C.; Oprea, M.; Spinu, A.; Popescu, C.; Ionescu, A.; et al. Cellular and Molecular Targets for Non-Invasive, Non-Pharmacological Therapeutic/Rehabilitative Interventions in Acute Ischemic Stroke. Int. J. Mol. Sci. 2022, 23, 907. [Google Scholar] [CrossRef]
- Bithi, N.; Link, C.; Henderson, Y.O.; Kim, S.; Yang, J.; Li, L.; Wang, R.; Willard, B.; Hine, C. Dietary restriction transforms the mammalian protein persulfidome in a tissue-specific and cystathionine γ-lyase-dependent manner. Nat. Commun. 2021, 12, 1745. [Google Scholar] [CrossRef]
- Wang, W.-J.; Cai, G.-Y.; Ning, Y.-C.; Cui, J.; Hong, Q.; Bai, X.-Y.; Xu, X.-M.; Bu, R.; Sun, X.-F.; Chen, X.-M. Hydrogen sulfide mediates the protection of dietary restriction against renal senescence in aged F344 rats. Sci. Rep. 2016, 6, 30292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renieris, G.; Katrini, K.; Damoulari, C.; Akinosoglou, K.; Psarrakis, C.; Kyriakopoulou, M.; Dimopoulos, G.; Lada, M.; Koufargyris, P.; Giamarellos-Bourboulis, E.J. Serum Hydrogen Sulfide and Outcome Association in Pneumonia by the SARS-CoV-2 Coronavirus. Shock 2020, 54, 633–763. [Google Scholar] [CrossRef] [PubMed]
- Citi, V.; Martelli, A.; Brancaleone, V.; Brogi, S.; Gojon, G.; Montanaro, R.; Morales, G.; Testai, L.; Calderone, V. Anti-inflammatory and antiviral roles of hydrogen sulfide: Rationale for considering H2S donors in COVID-19 therapy. Br. J. Pharmacol. 2020, 177, 4931–4941. [Google Scholar] [CrossRef] [PubMed]
- Evgen’ev, M.B.; Frenkel, A. Possible application of H2S-producing compounds in therapy of coronavirus (COVID-19) infection and pneumonia. Cell Stress Chaperones 2020, 25, 713–715. [Google Scholar] [CrossRef]
- Datzmann, T.; Merz, T.; McCook, O.; Szabo, C.; Radermacher, P. H2S as a Therapeutic Adjuvant Against COVID-19: Why and How? Shock 2021, 56, 865–867. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Keywords in Title, Abstract or Author-Specified Keywords | Elsevier | PubMed | PMC | ISI | Total |
|---|---|---|---|---|---|
| “Hydrogen sulfide” AND “Diabetes” | 8 | 63 | 45 | 59 | 175 |
| “H2S” AND “Diabetes” | 1 | 14 | 8 | 14 | 37 |
| Total | 9 | 77 | 53 | 73 | 212 |
| Physiological Properties of H2S | ||
|---|---|---|
| Authors | Ref. No. | Subject-Data |
| (Sun, 2021) | [3] | An Updated Insight Into Molecular Mechanism of H2S in Cardiomyopathy |
| (George, 2018) | [6] | Treating inflammation and oxidative stress with H2S during age-related macular degeneration |
| (Zou, 2017) | [10] | H2S ameliorates cognitive dysfunction in streptozotocin-induced diabetic rats |
| (Rey, 2021) | [11] | Mitochondrial metabolism as target of the neuroprotective role of erythropoietin in Parkinson’s disease. |
| (Testai, 2021) | [12] | Modulation of EndMT by H2S in the Prevention of Cardiovascular Fibrosis |
| (Ciccone, 2021) | [13] | Endothelium as a Source and Target of H2S to Improve Its Trophism and Function |
| (Wu, 2017) | [16] | Exogenous H2S facilitating ubiquitin aggregates clearance via autophagy |
| (Hu, 2017) | [20] | Chelerythrine Attenuates Renal Ischemia/Reperfusion-induced Myocardial Injury |
| (Kar, 2019) | [22] | H2S -mediated regulation of cell death signaling ameliorates adverse cardiac remodeling |
| (Jeong, 2020) | [24] | Protective effect of H2S on oxidative stress-induced neurodegenerative diseases |
| (Luo, 2019) | [25] | H2S upregulates renal AQP-2 protein expression and promotes urine concentration |
| (Yang, 2019) | [26] | Exogenous H2S mitigates myocardial fibrosis through suppression of Wnt pathway |
| (Liu, 2018) | [27] | H2S attenuates myocardial fibrosis through the JAK/STAT signaling pathway |
| (Sun, 2019) | [28] | Exogenous H2S reduces the acetylation levels of mitochondrial respiratory enzymes |
| (Roa-Coria, 2019) | [29] | Possible involvement of peripheral TRP channels in the H2S-induced hyperalgesia |
| (Yang, 2017) | [30] | Exogenous H2S regulates endoplasmic reticulum-mitochondria crosstalk to inhibit apoptosis |
| (Zhao, 2021) | [31] | H2S Plays an Important Role in Diabetic Cardiomyopathy |
| (Liu, 2017) | [32] | H2S modulating mitochondrial morphology to promote mitophagy in endothelial cells |
| (Qiu, 2018) | [33] | Alpha-lipoic acid regulates the autophagy of vascular smooth muscle cells elevating H2S level |
| (Li, 2017) | [34] | H2S reduced renal tissue fibrosis by regulating autophagy in diabetic rats |
| (Yu, 2020) | [35] | Exogenous H2S Induces Hrd1 S-sulfhydration and Prevents CD36 Translocation via VAMP3 |
| (Kar, 2019) | [36] | H2S Ameliorates Homocysteine-Induced Cardiac Remodeling and Dysfunction |
| (Dominic, 2021) | [37] | Decreased availability of nitric oxide and H2S is a hallmark of COVID-19 |
| (Loiselle, 2020) | [38] | H2S and hepatic lipid metabolism-a critical pairing for liver health |
| (Ma, 2017) | [39] | Exogenous H2S Ameliorates Diabetes-Associated Cognitive Decline |
| (Jiang, 2020) | [40] | H2S Ameliorates Lung Ischemia-Reperfusion Injury Through SIRT1 Signaling Pathway |
| (Wu, 2019) | [41] | H2S Inhibits High Glucose-Induced Neuronal Senescence by Improving Autophagic Flux |
| Pathophysiological Properties H2S | ||
| Authors | Ref. No. | Subject-Data |
| (Citi, 2021) | [7] | Role of H2S in endothelial dysfunction: Pathophysiology and therapeutic approaches |
| (Kang, 2020) | [14] | H2S as a Potential Alternative for the Treatment of Myocardial Fibrosis |
| (Sun, 2019) | [42] | H2S and Subsequent Liver Injury |
| (Szabo, 2017) | [43] | Pharmacological Modulation of H2S Levels |
| (Sun, 2020) | [44] | The Link Between Inflammation and H2S |
| (Zheng, 2020) | [45] | H2S protects against diabetes-accelerated atherosclerosis by preventing the activation of NLRP3 |
| (Jia, 2020) | [46] | H2S mitigates myocardial inflammation by inhibiting nucleotide-binding oligomerization domain-like receptor protein 3 inflammasome activation in diabetic rats |
| (Li, 2017) | [47] | H2S improves renal fibrosis in STZ-induced diabetic rats by ameliorating TGF-beta 1 expression |
| (Kar, 20190 | [48] | Exercise Training Promotes Cardiac H2S Biosynthesis and Mitigates Pyroptosis |
| (Li, 2019) | [49] | Exogenous H2S protects against high glucose-induced apoptosis and oxidative stress |
| H2S—Role in Diabetes Mellitus and Associated Vascular Pathology | ||
| Authors | Ref. No. | Subject-Data |
| (Gheibi, 2020) | [8] | Regulation of carbohydrate metabolism by NO and H2S: Implications in diabetes |
| (Zhang, 2021) | [50] | H2S regulates insulin secretion and insulin resistance in diabetes mellitus |
| (Chen, 2021) | [51] | Role of H2S in the Endocrine System |
| (Gheibi, 2019) | [52] | Effects of H2S on Carbohydrate Metabolism in Obese Type 2 Diabetic Rats |
| (Luo, 2017) | [53] | The Role of Exogenous H2S in Free Fatty Acids Induced Inflammation in Macrophages |
| (Comas, 2021) | [54] | The Impact of H2S on Obesity-Associated Metabolic Disturbances |
| (Suzuki, 2017) | [55] | Clinical Implication of Plasma H2S Levels in Japanese Patients with Type 2 Diabetes |
| (Zhou, 2019) | [56] | H2S Prevents Elastin Loss and Attenuates Calcification Induced by High Glucose |
| H2S—As a Natural Therapeutic Factor in DM | ||
| Authors | Ref. No. | Subject-Data |
| (Melino, 2019) | [2] | Natural H2S Donors from Allium sp. as a Nutraceutical Approach in Type 2 Diabetes |
| (Sashi, 2019) | [5] | H2S inhibits Ca2+-induced mitochondrial permeability transition pore opening |
| (Yang, 2017) | [21] | H2S Releasing/Stimulating Reagents |
| (John, 2017) | [57] | GYY4137, an H2S Donor Modulates miR194-Dependent Collagen Realignment |
| (Bitar, 2018) | [58] | H2S Donor NaHS Improves Metabolism and Reduces Muscle Atrophy in Type 2 Diabetes |
| (Ding, 2017) | [59] | High Glucose Induces Mouse Mesangial Cell Overproliferation via Inhibition of H2S Synthesis |
| No. | Study | Start Year | END Year | N-Total Subjects | Diabetes Mellitus | Cardiovascular/ Associated Disease | Neurodegenerative/ Associated Disease | Respiratory/ Associated Disease | Sodium Thiosulfate | Taurine | Captopril/Enalapril/Hydrochlorothiazide | Observational |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | AXIS LEGEND | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |||
| 1 | NCT03410173 | 2017 | 2018 | 200 | 200 | 200 | ||||||
| 2 | NCT03410537 | 2017 | 2018 | 200 | 200 | 200 | ||||||
| 3 | NCT02180074 | 2013 | 2017 | 137 | 137 | 137 | ||||||
| 4 | NCT03829605 | 2019 | 2019 | 50 | 50 | 50 | ||||||
| 5 | NCT05060848 | 2021 | 2027 | 100 | 100 | 100 | ||||||
| 6 | NCT04952883 | 2016 | 2017 | 100 | 100 | 100 | ||||||
| 7 | NCT02899364 | 2018 | 2023 | 380 | 380 | 380 | ||||||
| 8 | NCT03179163 | 2016 | 2021 | 70 | 70 | 70 | ||||||
| TOTAL | 1237 | 400 | 637 | 100 | 100 | 380 | 400 | 70 | 387 | |||
| % | 100 | 32.34 | 51.50 | 8.08 | 8.08 | 30.72 | 32.34 | 5.66 | 31.29 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munteanu, C.; Rotariu, M.; Turnea, M.; Dogaru, G.; Popescu, C.; Spînu, A.; Andone, I.; Postoiu, R.; Ionescu, E.V.; Oprea, C.; et al. Recent Advances in Molecular Research on Hydrogen Sulfide (H2S) Role in Diabetes Mellitus (DM)—A Systematic Review. Int. J. Mol. Sci. 2022, 23, 6720. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126720
Munteanu C, Rotariu M, Turnea M, Dogaru G, Popescu C, Spînu A, Andone I, Postoiu R, Ionescu EV, Oprea C, et al. Recent Advances in Molecular Research on Hydrogen Sulfide (H2S) Role in Diabetes Mellitus (DM)—A Systematic Review. International Journal of Molecular Sciences. 2022; 23(12):6720. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126720
Chicago/Turabian StyleMunteanu, Constantin, Mariana Rotariu, Marius Turnea, Gabriela Dogaru, Cristina Popescu, Aura Spînu, Ioana Andone, Ruxandra Postoiu, Elena Valentina Ionescu, Carmen Oprea, and et al. 2022. "Recent Advances in Molecular Research on Hydrogen Sulfide (H2S) Role in Diabetes Mellitus (DM)—A Systematic Review" International Journal of Molecular Sciences 23, no. 12: 6720. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126720