
A Panel of Eight miRNAs Is Deregulated in HTLV-2 Infected PBMCs and BJABGu Cell Line

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
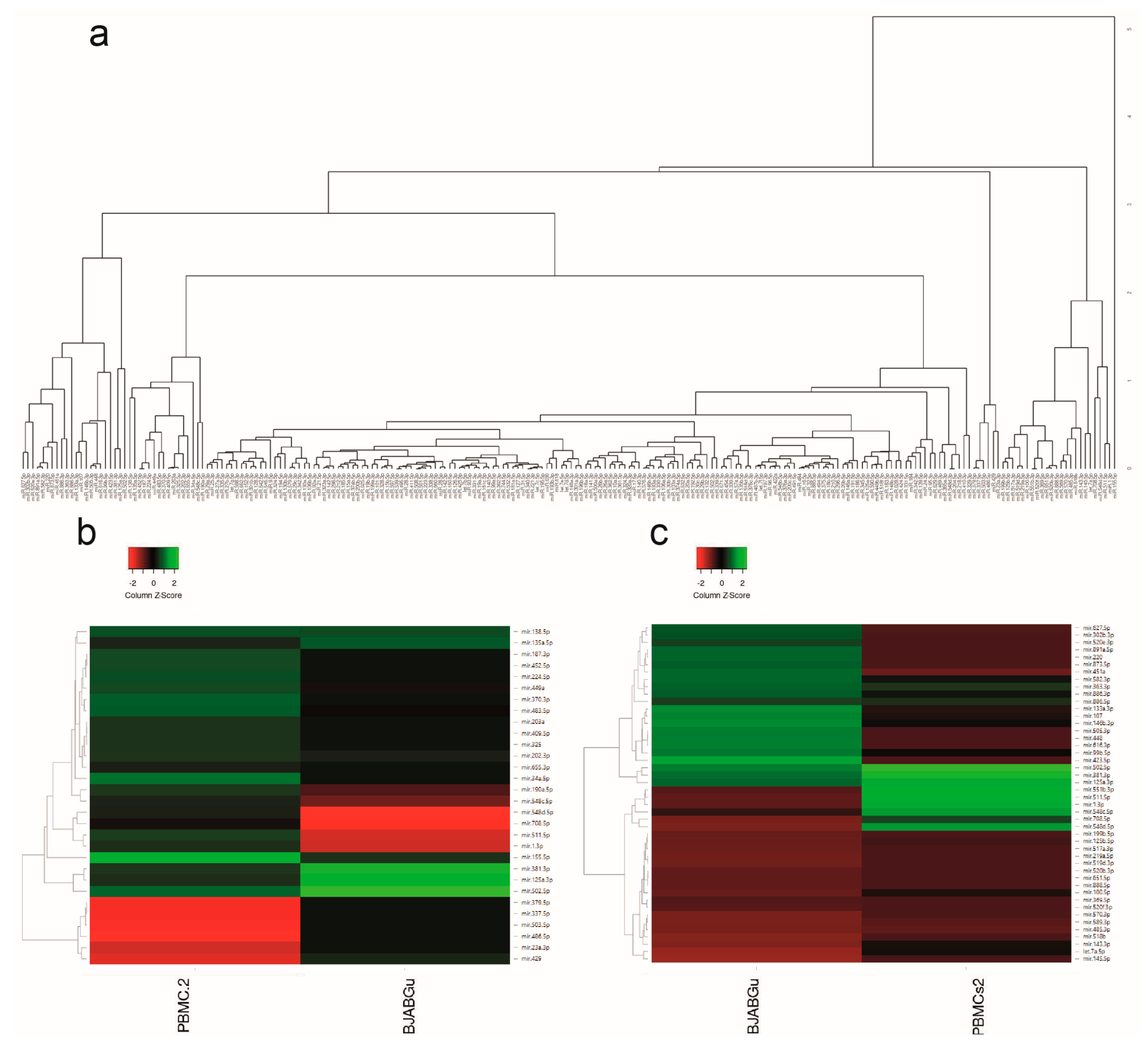
2.1. Identification of Distinct miRNAs Expression Profiles in PBMCs from Mono-HTLV-2 Positive Subjects and in BJABGu Cells
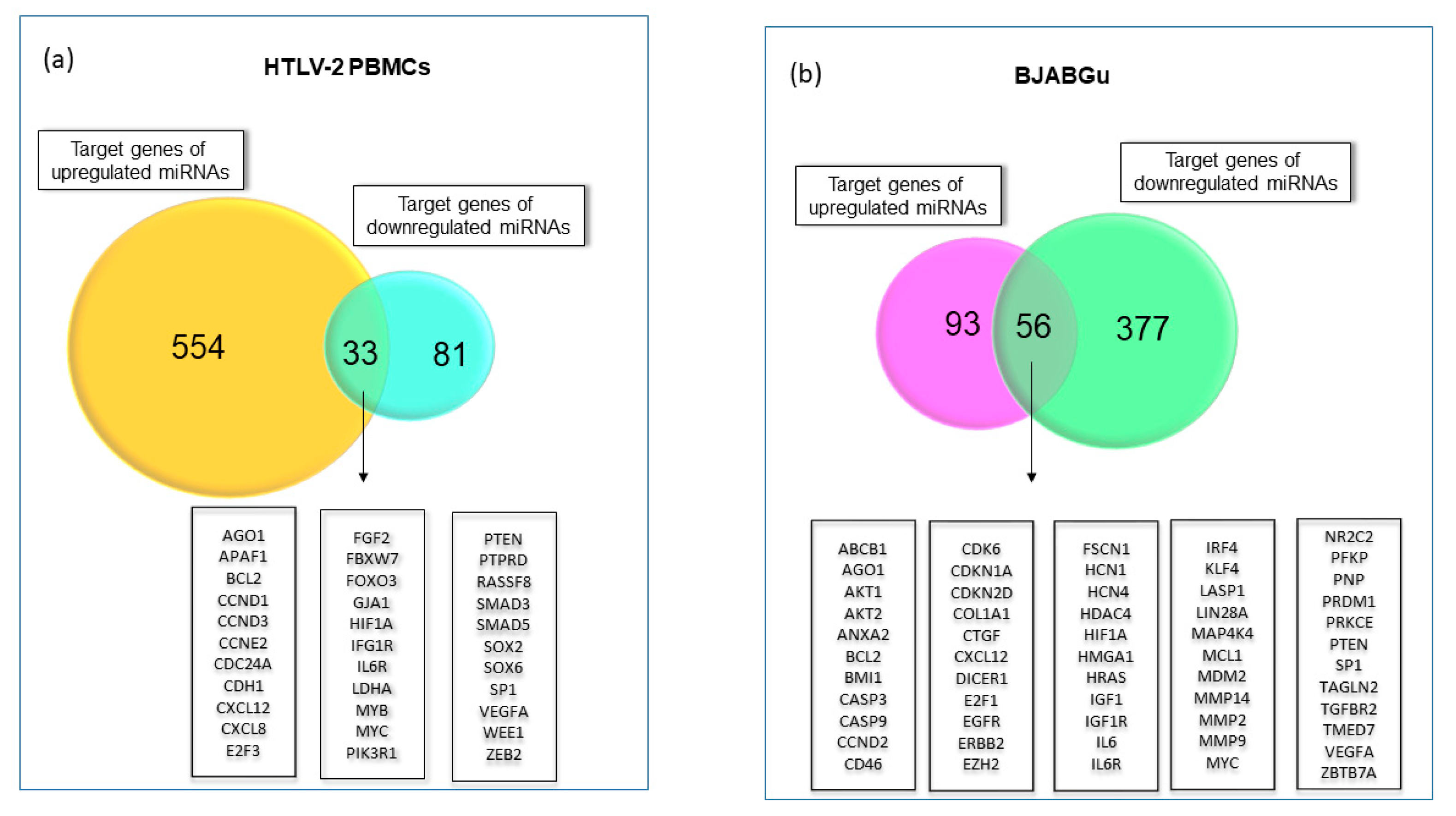
2.2. Exploring miRNA Target Genes
2.3. Top Enriched Pathways Related to miRNAs Affected by HTLV-2 Were Driven by Mediators of Cell Activation and Differentiation
2.4. RUNX2 Is Deregulated in Presence of Tax Proteins
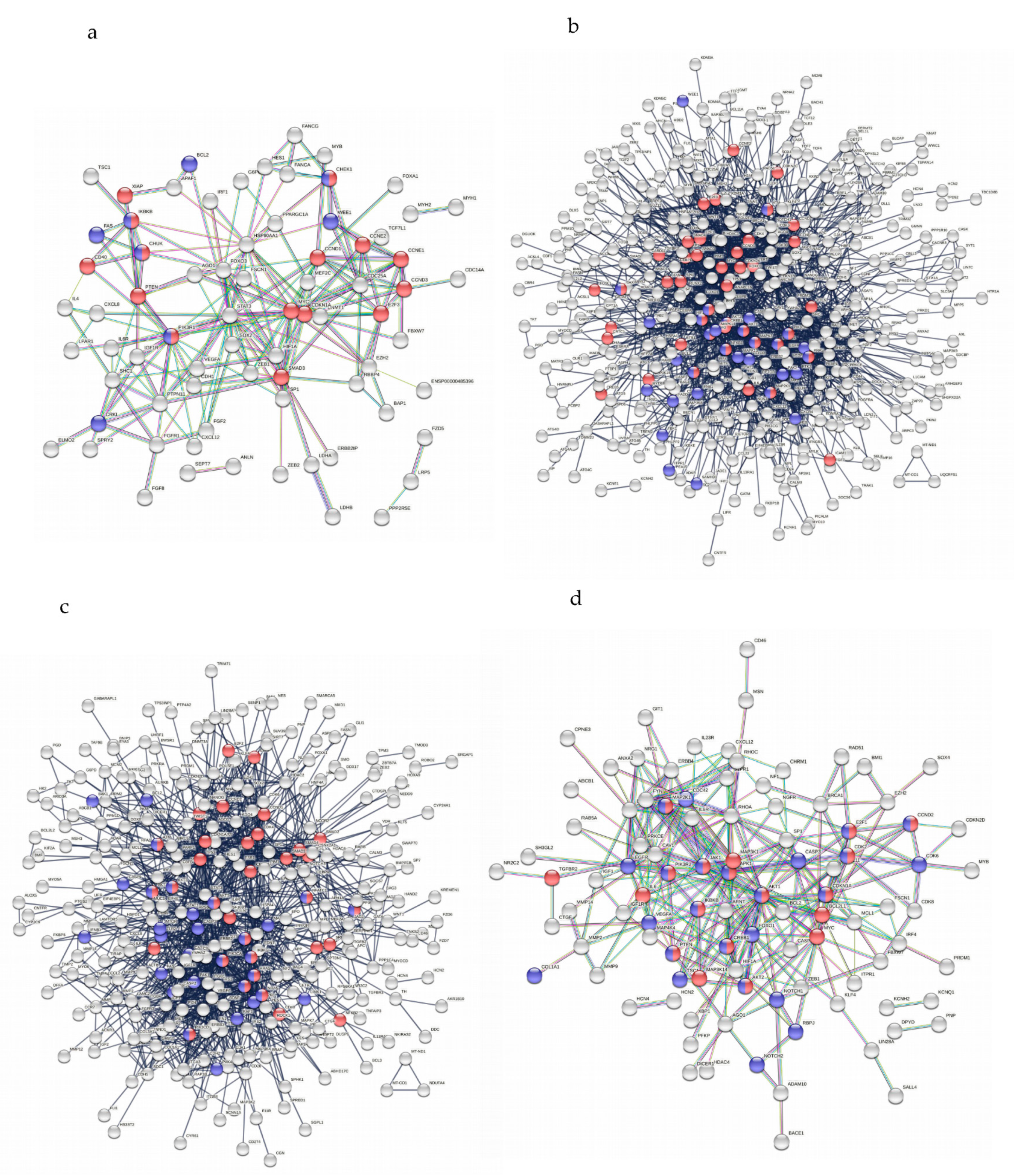
2.5. Multiple Overlapping Pathways in Different Single Node Networks Revealed Potential Strong Functional Relationships
3. Discussion
4. Materials and Methods
4.1. Primary and Immortalized Cells
4.2. miRNA Expression Analysis
4.3. Functional Analysis of miRNAs Target Genes
4.4. Cell Lines and Transfection
4.5. Plasmids
4.6. Protein Extraction
4.7. Western Blotting
4.8. Antibodies
4.9. RT-qPCR Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.P.E.; Lewis, A.P.; Jopling, C.L. The Role of MicroRNAs in Viral Infection. Prog. Mol. Biol. Transl. Sci. 2011, 102, 101–139. [Google Scholar] [CrossRef] [PubMed]
- Vojtechova, Z.; Tachezy, R. The Role of MiRNAs in Virus-Mediated Oncogenesis. Int. J. Mol. Sci. 2018, 19, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramaniam, M.; Pandhare, J.; Dash, C. Are MicroRNAs Important Players in HIV-1 Infection? An Update. Viruses 2018, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Sampey, G.C.; Van Duyne, R.; Currer, R.; Das, R.; Narayanan, A.; Kashanchi, F. Complex Role of MicroRNAs in HTLV-1 Infections. Front. Genet. 2012, 3, 295. [Google Scholar] [CrossRef] [Green Version]
- Sanghvi, V.R.; Steel, L.F. RNA Silencing as a Cellular Defense against HIV-1 Infection: Progress and Issues. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2012, 26, 3937–3945. [Google Scholar] [CrossRef] [PubMed]
- Houzet, L.; Jeang, K.-T. MicroRNAs and Human Retroviruses. Biochim. Biophys. Acta 2011, 1809, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, J.; Aslan, C.; Ahmadi, M.; Zolbanin, N.M.; Kashanchi, F.; Jafari, R. The Versatile Role of Exosomes in Human Retroviral Infections: From Immunopathogenesis to Clinical Application. Cell Biosci. 2021, 11, 19. [Google Scholar] [CrossRef]
- Lairmore, M.D.; Franchini, G. Human T-Cell Leukemia Virus Types I and II. In Fields Virology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2071–2106. ISBN 978-1-4698-6774-8. [Google Scholar]
- Mahieux, R.; Gessain, A. HTLV-3/STLV-3 and HTLV-4 Viruses: Discovery, Epidemiology, Serology and Molecular Aspects. Viruses 2011, 3, 1074–1090. [Google Scholar] [CrossRef]
- Pique, C.; Jones, K.S. Pathways of Cell-Cell Transmission of HTLV-1. Front. Microbiol. 2012, 3, 378. [Google Scholar] [CrossRef] [Green Version]
- Ciminale, V.; Rende, F.; Bertazzoni, U.; Romanelli, M.G. HTLV-1 and HTLV-2: Highly Similar Viruses with Distinct Oncogenic Properties. Front. Microbiol. 2014, 5, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwanaga, M.; Watanabe, T.; Yamaguchi, K. Adult T-Cell Leukemia: A Review of Epidemiological Evidence. Front. Microbiol. 2012, 3, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertazzoni, U.; Ciminale, V.; Romanelli, M.G. Editorial: Molecular Pathology of HTLV-1. Front. Microbiol. 2018, 9, 3069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilotti, E.; Bianchi, M.V.; De Maria, A.; Bozzano, F.; Romanelli, M.G.; Bertazzoni, U.; Casoli, C. HTLV-1/-2 and HIV-1 Co-Infections: Retroviral Interference on Host Immune Status. Front. Microbiol. 2013, 4, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjelle, B.; Appenzeller, O.; Mills, R.; Alexander, S.; Torrez-Martinez, N.; Jahnke, R.; Ross, G. Chronic Neurodegenerative Disease Associated with HTLV-II Infection. Lancet Lond. Engl. 1992, 339, 645–646. [Google Scholar] [CrossRef]
- Casoli, C.; Pilotti, E.; Bertazzoni, U. Molecular and Cellular Interactions of HIV-1/HTLV Coinfection and Impact on AIDS Progression. AIDS Rev. 2007, 9, 140–149. [Google Scholar]
- Beilke, M.A.; Theall, K.P.; O’Brien, M.; Clayton, J.L.; Benjamin, S.M.; Winsor, E.L.; Kissinger, P.J. Clinical Outcomes and Disease Progression among Patients Coinfected with HIV and Human T Lymphotropic Virus Types 1 and 2. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2004, 39, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Melamed, A.; Witkover, A.D.; Laydon, D.J.; Brown, R.; Ladell, K.; Miners, K.; Rowan, A.G.; Gormley, N.; Price, D.A.; Taylor, G.P.; et al. Clonality of HTLV-2 in Natural Infection. PLoS Pathog. 2014, 10, e1004006. [Google Scholar] [CrossRef] [Green Version]
- Forlani, G.; Shallak, M.; Accolla, R.S.; Romanelli, M.G. HTLV-1 Infection and Pathogenesis: New Insights from Cellular and Animal Models. Int. J. Mol. Sci. 2021, 22, 8001. [Google Scholar] [CrossRef]
- Heym, S.; Mohr, C.F.; Engelbrecht, H.C.; Fleckenstein, B.; Thoma-Kress, A.K. Alternative NF-ΚB Signaling Discriminates Induction of the Tumor Marker Fascin by the Viral Oncoproteins Tax-1 and Tax-2 of Human T-Cell Leukemia Viruses. Cancers 2022, 14, 537. [Google Scholar] [CrossRef]
- Martinez, M.P.; Al-Saleem, J.; Green, P.L. Comparative Virology of HTLV-1 and HTLV-2. Retrovirology 2019, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Millen, S.; Meretuk, L.; Göttlicher, T.; Schmitt, S.; Fleckenstein, B.; Thoma-Kress, A.K. A Novel Positive Feedback-Loop between the HTLV-1 Oncoprotein Tax and NF-ΚB Activity in T-Cells. Retrovirology 2020, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, M.G.; Diani, E.; Bergamo, E.; Casoli, C.; Ciminale, V.; Bex, F.; Bertazzoni, U. Highlights on Distinctive Structural and Functional Properties of HTLV Tax Proteins. Front. Microbiol. 2013, 4, 271. [Google Scholar] [CrossRef] [Green Version]
- Shirinian, M.; Kfoury, Y.; Dassouki, Z.; El-Hajj, H.; Bazarbachi, A. Tax-1 and Tax-2 Similarities and Differences: Focus on Post-Translational Modifications and NF-ΚB Activation. Front. Microbiol. 2013, 4, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avesani, F.; Romanelli, M.G.; Turci, M.; Di Gennaro, G.; Sampaio, C.; Bidoia, C.; Bertazzoni, U.; Bex, F. Association of HTLV Tax Proteins with TAK1-Binding Protein 2 and RelA in Calreticulin-Containing Cytoplasmic Structures Participates in Tax-Mediated NF-ΚB Activation. Virology 2010, 408, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Fochi, S.; Mutascio, S.; Bertazzoni, U.; Zipeto, D.; Romanelli, M.G. HTLV Deregulation of the NF-ΚB Pathway: An Update on Tax and Antisense Proteins Role. Front. Microbiol. 2018, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fochi, S.; Ciminale, V.; Trabetti, E.; Bertazzoni, U.; D’Agostino, D.M.; Zipeto, D.; Romanelli, M.G. NF-ΚB and MicroRNA Deregulation Mediated by HTLV-1 Tax and HBZ. Pathogens 2019, 8, 290. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, M.; Tsubata, C.; Kondo, R.; Yoshida, S.; Takahashi, M.; Oie, M.; Tanaka, Y.; Mahieux, R.; Matsuoka, M.; Fujii, M. Cooperation of NF-KappaB2/P100 Activation and the PDZ Domain Binding Motif Signal in Human T-Cell Leukemia Virus Type 1 (HTLV-1) Tax1 but Not HTLV-2 Tax2 Is Crucial for Interleukin-2-Independent Growth Transformation of a T-Cell Line. J. Virol. 2007, 81, 11900–11907. [Google Scholar] [CrossRef] [Green Version]
- Sheehy, N.; Lillis, L.; Watters, K.; Lewis, M.; Gautier, V.; Hall, W. Functional Analysis of Human T Lymphotropic Virus Type 2 Tax Proteins. Retrovirology 2006, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- Bertazzoni, U.; Turci, M.; Avesani, F.; Di Gennaro, G.; Bidoia, C.; Romanelli, M.G. Intracellular Localization and Cellular Factors Interaction of HTLV-1 and HTLV-2 Tax Proteins: Similarities and Functional Differences. Viruses 2011, 3, 541–560. [Google Scholar] [CrossRef] [Green Version]
- Meertens, L.; Chevalier, S.; Weil, R.; Gessain, A.; Mahieux, R. A 10-Amino Acid Domain within Human T-Cell Leukemia Virus Type 1 and Type 2 Tax Protein Sequences Is Responsible for Their Divergent Subcellular Distribution. J. Biol. Chem. 2004, 279, 43307–43320. [Google Scholar] [CrossRef] [Green Version]
- Turci, M.; Romanelli, M.G.; Lorenzi, P.; Righi, P.; Bertazzoni, U. Localization of Human T-Cell Lymphotropic Virus Type II Tax Protein Is Dependent upon a Nuclear Localization Determinant in the N-Terminal Region. Gene 2006, 365, 119–124. [Google Scholar] [CrossRef]
- Lewis, M.J.; Sheehy, N.; Salemi, M.; VanDamme, A.-M.; Hall, W.W. Comparison of CREB- and NF-KappaB-Mediated Transactivation by Human T Lymphotropic Virus Type II (HTLV-II) and Type I (HTLV-I) Tax Proteins. Virology 2002, 295, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Bellon, M.; Lepelletier, Y.; Hermine, O.; Nicot, C. Deregulation of MicroRNA Involved in Hematopoiesis and the Immune Response in HTLV-I Adult T-Cell Leukemia. Blood 2009, 113, 4914–4917. [Google Scholar] [CrossRef] [Green Version]
- Moles, R.; Nicot, C. The Emerging Role of MiRNAs in HTLV-1 Infection and ATLL Pathogenesis. Viruses 2015, 7, 4047–4074. [Google Scholar] [CrossRef] [Green Version]
- Ruggero, K.; Corradin, A.; Zanovello, P.; Amadori, A.; Bronte, V.; Ciminale, V.; D’Agostino, D.M. Role of MicroRNAs in HTLV-1 Infection and Transformation. Mol. Aspects Med. 2010, 31, 367–382. [Google Scholar] [CrossRef]
- Shadabi, S.; Delrish, N.; Norouzi, M.; Ehteshami, M.; Habibian-Sezavar, F.; Pourrezaei, S.; Madihi, M.; Ostadali, M.; Akhgar, F.; Shayeghpour, A.; et al. Comprehensive High-Throughput Meta-Analysis of Differentially Expressed MicroRNAs in Transcriptomic Datasets Reveals Significant Disruption of MAPK/JNK Signal Transduction Pathway in Adult T-Cell Leukemia/Lymphoma. Infect. Agent. Cancer 2021, 16, 49. [Google Scholar] [CrossRef]
- Yamagishi, M.; Nakano, K.; Miyake, A.; Yamochi, T.; Kagami, Y.; Tsutsumi, A.; Matsuda, Y.; Sato-Otsubo, A.; Muto, S.; Utsunomiya, A.; et al. Polycomb-Mediated Loss of MiR-31 Activates NIK-Dependent NF-ΚB Pathway in Adult T Cell Leukemia and Other Cancers. Cancer Cell 2012, 21, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Bignami, F.; Pilotti, E.; Bertoncelli, L.; Ronzi, P.; Gulli, M.; Marmiroli, N.; Magnani, G.; Pinti, M.; Lopalco, L.; Mussini, C.; et al. Stable Changes in CD4+ T Lymphocyte MiRNA Expression after Exposure to HIV-1. Blood 2012, 119, 6259–6267. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.; Zhang, W.; Li, H.; Shen, X.; Guo, Z. Separate Enrichment Analysis of Pathways for Up- and Downregulated Genes. J. R. Soc. Interface 2014, 11, 20130950. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Solal, K.A.; Boregowda, R.K.; Lasfar, A. RUNX2 and the PI3K/AKT Axis Reciprocal Activation as a Driving Force for Tumor Progression. Mol. Cancer 2015, 14, 137. [Google Scholar] [CrossRef] [Green Version]
- Blyth, K.; Vaillant, F.; Hanlon, L.; Mackay, N.; Bell, M.; Jenkins, A.; Neil, J.C.; Cameron, E.R. Runx2 and MYC Collaborate in Lymphoma Development by Suppressing Apoptotic and Growth Arrest Pathways In Vivo. Cancer Res. 2006, 66, 2195–2201. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.-H.; Zaidi, S.K.; Gornostaeva, S.; Komori, T.; Stein, G.S.; Castilla, L.H. Runx2 Induces Acute Myeloid Leukemia in Cooperation with Cbfbeta-SMMHC in Mice. Blood 2009, 113, 3323–3332. [Google Scholar] [CrossRef] [Green Version]
- Chopin, M.; Preston, S.P.; Lun, A.T.L.; Tellier, J.; Smyth, G.K.; Pellegrini, M.; Belz, G.T.; Corcoran, L.M.; Visvader, J.E.; Wu, L.; et al. RUNX2 Mediates Plasmacytoid Dendritic Cell Egress from the Bone Marrow and Controls Viral Immunity. Cell Rep. 2016, 15, 866–878. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Dong, X.; Liu, X.; Wang, G.; Dong, Z.; Shen, W.; Zheng, G.; Lu, J.; Chen, J.; Wang, Y.; et al. High-Throughput Functional MicroRNAs Profiling by Recombinant AAV-Based MicroRNA Sensor Arrays. PLoS ONE 2012, 7, e29551. [Google Scholar] [CrossRef] [Green Version]
- Bernier, A.; Sagan, S.M. The Diverse Roles of MicroRNAs at the Host–Virus Interface. Viruses 2018, 10, 440. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Wang, N.; Li, Z.; Zhang, Y.; Zheng, Y.; Yi, P.; Chen, J. MiR-125a-3p Suppresses the Growth and Progression of Papillary Thyroid Carcinoma Cell by Targeting MMP11. J. Cell. Biochem. 2020, 121, 984–995. [Google Scholar] [CrossRef]
- Ying, Y.; Li, J.; Xie, H.; Yan, H.; Jin, K.; He, L.; Ma, X.; Wu, J.; Xu, X.; Fang, J.; et al. CCND1, NOP14 and DNMT3B Are Involved in MiR-502-5p-Mediated Inhibition of Cell Migration and Proliferation in Bladder Cancer. Cell Prolif. 2020, 53, e12751. [Google Scholar] [CrossRef] [Green Version]
- Zhan, L.; Yang, J.; Liu, Y.; Cheng, Y.; Liu, H. MicroRNA MiR-502-5p Inhibits Ovarian Cancer Genesis by Downregulation of GINS Complex Subunit 2. Bioengineered 2021, 12, 3336–3347. [Google Scholar] [CrossRef]
- Bovolenta, C.; Pilotti, E.; Mauri, M.; Turci, M.; Ciancianaini, P.; Fisicaro, P.; Bertazzoni, U.; Poli, G.; Casoli, C. Human T-Cell Leukemia Virus Type 2 Induces Survival and Proliferation of CD34(+) TF-1 Cells through Activation of STAT1 and STAT5 by Secretion of Interferon-Gamma and Granulocyte Macrophage-Colony-Stimulating Factor. Blood 2002, 99, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Galán, A.; Fernández-Messina, L.; Sánchez-Madrid, F. Control of Immunoregulatory Molecules by MiRNAs in T Cell Activation. Front. Immunol. 2018, 9, 2148. [Google Scholar] [CrossRef] [PubMed]
- Falivene, J.; Ghiglione, Y.; Laufer, N.; Socías, M.E.; Holgado, M.P.; Ruiz, M.J.; Maeto, C.; Figueroa, M.I.; Giavedoni, L.D.; Cahn, P.; et al. Th17 and Th17/Treg Ratio at Early HIV Infection Associate with Protective HIV-Specific CD8(+) T-Cell Responses and Disease Progression. Sci. Rep. 2015, 5, 11511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, C.; Veyrenche, N.; Mennechet, F.; Bedin, A.-S.; Routy, J.-P.; Van de Perre, P.; Reynes, J.; Tuaillon, E. Th17 CD4+ T-Cell as a Preferential Target for HIV Reservoirs. Front. Immunol. 2022, 13, 822576. [Google Scholar] [CrossRef] [PubMed]
- Okkenhaug, K.; Turner, M.; Gold, M.R. PI3K Signaling in B Cell and T Cell Biology. Front. Immunol. 2014, 5, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panfil, A.R.; Dissinger, N.J.; Howard, C.M.; Murphy, B.M.; Landes, K.; Fernandez, S.A.; Green, P.L. Functional Comparison of HBZ and the Related APH-2 Protein Provides Insight into Human T-Cell Leukemia Virus Type 1 Pathogenesis. J. Virol. 2016, 90, 3760–3772. [Google Scholar] [CrossRef] [Green Version]
- Gülich, A.F.; Rica, R.; Tizian, C.; Viczenczova, C.; Khamina, K.; Faux, T.; Hainberger, D.; Penz, T.; Bosselut, R.; Bock, C.; et al. Complex Interplay Between MAZR and Runx3 Regulates the Generation of Cytotoxic T Lymphocyte and Memory T Cells. Front. Immunol. 2021, 12, 535039. [Google Scholar] [CrossRef]
- Jadhav, R.R.; Im, S.J.; Hu, B.; Hashimoto, M.; Li, P.; Lin, J.-X.; Leonard, W.J.; Greenleaf, W.J.; Ahmed, R.; Goronzy, J.J. Epigenetic Signature of PD-1+ TCF1+ CD8 T Cells That Act as Resource Cells during Chronic Viral Infection and Respond to PD-1 Blockade. Proc. Natl. Acad. Sci. USA 2019, 116, 14113–14118. [Google Scholar] [CrossRef] [Green Version]
- Olesin, E.; Nayar, R.; Saikumar-Lakshmi, P.; Berg, L.J. The Transcription Factor Runx2 Is Required for Long-Term Persistence of Antiviral CD8+ Memory T Cells. ImmunoHorizons 2018, 2, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Qin, X. MicroRNA-708 Targeting ZNF549 Regulates Colon Adenocarcinoma Development through PI3K/AKt Pathway. Sci. Rep. 2020, 10, 16729. [Google Scholar] [CrossRef]
- Ma, Y.; Zhou, G.; Li, M.; Hu, D.; Zhang, L.; Liu, P.; Lin, K. Long Noncoding RNA DANCR Mediates Cisplatin Resistance in Glioma Cells via Activating AXL/PI3K/Akt/NF-ΚB Signaling Pathway. Neurochem. Int. 2018, 118, 233–241. [Google Scholar] [CrossRef]
- Salemi, M.; Vandamme, A.M.; Guano, F.; Gradozzi, C.; Cattaneo, E.; Casoli, C.; Bertazzoni, U. Complete Nucleotide Sequence of the Italian Human T-Cell Lymphotropic Virus Type II Isolate Gu and Phylogenetic Identification of a Possible Origin of South European Epidemics. J. Gen. Virol. 1996, 77 Pt 6, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-Enabled Heat Mapping for All. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Aoki-Kinoshita, K.F.; Kanehisa, M. Gene Annotation and Pathway Mapping in KEGG. Methods Mol. Biol. 2007, 396, 71–91. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Turci, M.; Lodewick, J.; Di Gennaro, G.; Rinaldi, A.S.; Marin, O.; Diani, E.; Sampaio, C.; Bex, F.; Bertazzoni, U.; Romanelli, M.G. Ubiquitination and Sumoylation of the HTLV-2 Tax-2B Protein Regulate Its NF-ΚB Activity: A Comparative Study with the HTLV-1 Tax-1 Protein. Retrovirology 2012, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Fochi, S.; Orlandi, E.; Ceccuzzi, L.; Rodolfo, M.; Vergani, E.; Turco, A.; Romanelli, M.G.; Gomez-Lira, M. Identification of Suitable MRNAs and MicroRNAs as Reference Genes for Expression Analyses in Skin Cells under Sex Hormone Exposure. Gene 2021, 769, 145336. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactome Terms | Down miRNAs in Infected PBMCs | Up miRNAs in Infected PBMCs | Down miRNAs in BjabGu | Up miRNAs in BjabGu |
|---|---|---|---|---|
| Regulation of RUNX1 Expression and Activity | 1.61 *; 0.0018 ** | 1.14 *; 0.00055 ** | 1.21 *; 0.0009 ** | n.d. ‡ |
| Transcriptional regulation by RUNX2 | n.d | 0.83 *; 1.89 × 10−9 ** | 0.89 *; 9.55 × 10−9 ** | 0.9 *; 0.008 * |
| Transcriptional regulation by RUNX3 | 1.11 *; 0.00067 ** | 0.73 *; 0.0001 ** | 0.8 *; 8.8 × 10−5 ** | 1 *; 0.0023 ** |
| RUNX2 regulates genes involved in cell migration | n.d. | n.d. | 1.45 *; 0.00066 ** | n.d. |
| RUNX3 regulates WNT signaling | 1.81 *; 0.0071 ** | 1.32 *; 0.0025 ** | n.d. | n.d. |
| RUNX3 regulates CDKN1A transcription | n.d. | n.d | 1.41 *; 0.0057 ** | n.d |
| RUNX3 regulates p14-ARF | n.d. | n.d. | 1.31 *; 0.0103 ** | n.d. |
| Interleukin-6 signaling | 1.67 *; 0.0133 ** | 1.18 *; 0.0068 ** | 1.32 *; 0.0018 ** | 1.56 *; 0.0226 ** |
| Interleukin-4 and Interleukin-13 signaling | 1.44 *; 4.86 × 10−15 ** | 1.08 *; 1.7 × 10−22 ** | 1.13 *; 1.25 × 10−20 ** | 1.37 *; 8.07 × 10−16 ** |
| Interleukin-37 signaling | 1.39 *; 0.0487 ** | n.d. | n.d. | n.d. |
| Interleukin-3, Interleukin-5 and GM-CSF signaling | 1.16 *; 0.0372 ** | n.d. | 0.82 *; 0.0128 ** | n.d. |
| Interleukin-1 family signaling | 0.88 *; 0.0296 ** | n.d. | n.d. | n.d. |
| Regulation of IFNG signaling | n.d. | 1.08 *; 0.0143 ** | n.d. | n.d. |
| Interleukin-17 signaling | n.d. | 0.75 *; 0.00066 ** | n.d. | n.d. |
| Interleukin-2 family signaling | n.d. | n.d. | 0.87 *; 0.0078 ** | n.d. |
| TNF signaling | n.d. | n.d. | 0.79 *; 0.04 ** | n.d. |
| Interleukin-10 signaling | n.d. | n.d. | 0.78 *; 0.0439 ** | n.d. |
| Interleukin-12 signaling | n.d. | n.d. | n.d. | 1.16 *; 0.0075 ** |
| PI3k cascade: FGFR1 | 1.61 *; 0.00018 ** | n.d. | n.d. | n.d. |
| PI3k cascade: FGFR3 | 1.58 *; 0.0021 ** | n.d. | n.d. | n.d. |
| PI3k cascade: FGFR4 | 1.54 *; 0.0029 ** | 0.92 *; 0.041 ** | n.d. | n.d. |
| PI3k cascade: FGFR2 | 1.47 *; 0.0044 ** | n.d. | n.d. | n.d. |
| PI3k events in ERBB2 signaling | n.d. | 1.02 *; 0.0211 ** | 1.06 *; 0.0443 ** | 1.52 *; 0.0028 ** |
| AKT-mediated inactivation of FOXO1A | 1.4 *; 0.0461 ** | 1.53 *; 0.0217 ** | 2 *; 0.0029 ** | |
| Negative regulation of the PI3K/AKT network | 1.03 *; 0.0018 ** | 0.87 *; 1.28 × 10−10 ** | 0.93 *; 8.3 × 10−10 ** | 1.07 *; 1.58 × 10−5 ** |
| PI5P, PP2A and IER3 Regulate PI3K/AKT Signaling | 0.99 *; 0.0099 ** | 0.83 *; 4.31 × 10−8 ** | 0.89 *; 7.46 × 10−8 ** | 1.01 *; 0.00057 ** |
| CD28 dependent PI3K/Akt signaling | n.d. | 1.14 *; 3.68 × 10−5 ** | 1.09 *; 0.0026 ** | 1.48 *; 0.00045 ** |
| Activated NTRK3 signals through PI3K | n.d. | n.d. | 1.36 *; 0.0439 ** | n.d. |
| Erythropoietin activates Phosphoinositide-3-kinase (PI3K) | n.d. | n.d. | 1.42 *; 1.87 × 10−5 ** | n.d. |
| AKT phosphorylates targets in the cytosol | n.d. | n.d. | 1.29 *; 0.00043 ** | 1.58 *; 0.0019 ** |
| AKT phosphorylates targets in the nucleus | n.d. | n.d. | n.d. | 1.73 *; 0.00072 ** |
| miRNAs | HTLV-2+ vs. Uninfected | BJABGu vs. BJAB | Targets | KEGG GEA | Reactome GEA |
|---|---|---|---|---|---|
| 125a-3p | 2.07 (FE) | 2.59 (FE) | GPC4, MTA1, PRDM1, RHOA, FYN, IL23R, GIT1, VEGFA, IRF4, XBP1, EZH2, BRCA1, IL6, NRG1 | EGFR tyrosine kinase inhibitor resistance (1.73 *; 0.0055 **) Inflammatory bowel disease (1.67; 0.0497) Th17 cell differentiation (1.62; 0.0077) | ERBB2 signaling (1.92; 0.0387) IL-4 and IL-13 signaling (1.72; 0.0163) Interleukins signaling (1.2; 0.0435) |
| 708-5p | 1.04 (FE) | −3.13 (FE) | BMI1, ZEB2, BIRC5, AKT2, CD44, TMEM88, EYA3, NNAT, AKT1, CCND1, MMP2, EZH2, PARP1, BCL2, CASP2, CD274, CNTFR, SMAD3, IKBKG, KDM1A | Pancreatic cancer (1.83; 1.8 × 10−6) Chronic myeloid leukemia (1.81; 1.8 × 10−6) AGE-RAGE signaling pathway in diabetic complications (1.78; 3.21 × 10−7) | AKT-mediated inactivation of FOXO1A (2.69; 0.0161) RUNX2 regulates genes involved in cell migration (2.39; 0.0362) Activation of BAD and translocation to mitochondria (2.29; 0.0016) |
| 1-3p | 2.07 (FE) | −2.36 (FE) | CEBPA, MEF2A, GATA4, HCN4, HDAC4, FOXP1, HCN2, PTMA, PTBP1, MET, CAND1, ANXA2, HAND2, IGF1, TMSB4X, KCNJ2, GJA1, XPO6, POGK, TAGLN2, LASP1, ADAR, KCNE1, BDNF, G6PD, SOX6, ATP6V1B2, LARP4, CNN3, PNP, KIF2A, HSPD1, HSPA4, PIM1, CALM3, PPP2R5A, PAX3, TWF1, TWF2, FN1, NOTCH3, SLC8A1, EDN1, PRKCE, FABP3, SNAI2, SOX9, PGD, SRF, IL11, YWHAZ, CCND1, MYOCD, TKT, CCL2, CDK4, SP1, ETS1, FASN, PIK3CA, TH, MPL, API5, SPRED1, ASPH, ND1, COX1, FRS2, FZD7, AGO1, KRAS, NAIP, VEGFA, RARB, BAG4, ABCB1, TWIST1, TNKS2, CXCL12 | Pentose phosphate pathway (1.41; 0.0039) AGE-RAGE signaling pathway in diabetic complications (1.4; 7.86 × 10−9) Bladder cancer (1.38; 0.00071) | RUNX3 regulates p14-ARF (1.92; 0.0123) Signaling by FGFR3 fusions in cancer (1.87; 0.0126) Signaling by FGFR4 in disease (1.83; 0.0128) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilotti, E.; Cannata, A.; Magnani, G.; Bignami, F.; Corsi, A.; Valenti, M.T.; Shallak, M.; Forlani, G.; Romanelli, M.G. A Panel of Eight miRNAs Is Deregulated in HTLV-2 Infected PBMCs and BJABGu Cell Line. Int. J. Mol. Sci. 2022, 23, 7583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147583
Pilotti E, Cannata A, Magnani G, Bignami F, Corsi A, Valenti MT, Shallak M, Forlani G, Romanelli MG. A Panel of Eight miRNAs Is Deregulated in HTLV-2 Infected PBMCs and BJABGu Cell Line. International Journal of Molecular Sciences. 2022; 23(14):7583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147583
Chicago/Turabian StylePilotti, Elisabetta, Attilio Cannata, Giacomo Magnani, Fabio Bignami, Andrea Corsi, Maria Teresa Valenti, Mariam Shallak, Greta Forlani, and Maria Grazia Romanelli. 2022. "A Panel of Eight miRNAs Is Deregulated in HTLV-2 Infected PBMCs and BJABGu Cell Line" International Journal of Molecular Sciences 23, no. 14: 7583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147583