
TNF-α Activating Osteoclasts in Patients with Psoriatic Arthritis Enhances the Recruitment of Osteoclast Precursors: A Plausible Role of WNT5A-MCP-1 in Osteoclast Engagement in Psoriatic Arthritis

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. The Demographics of Patients with PsA and HC
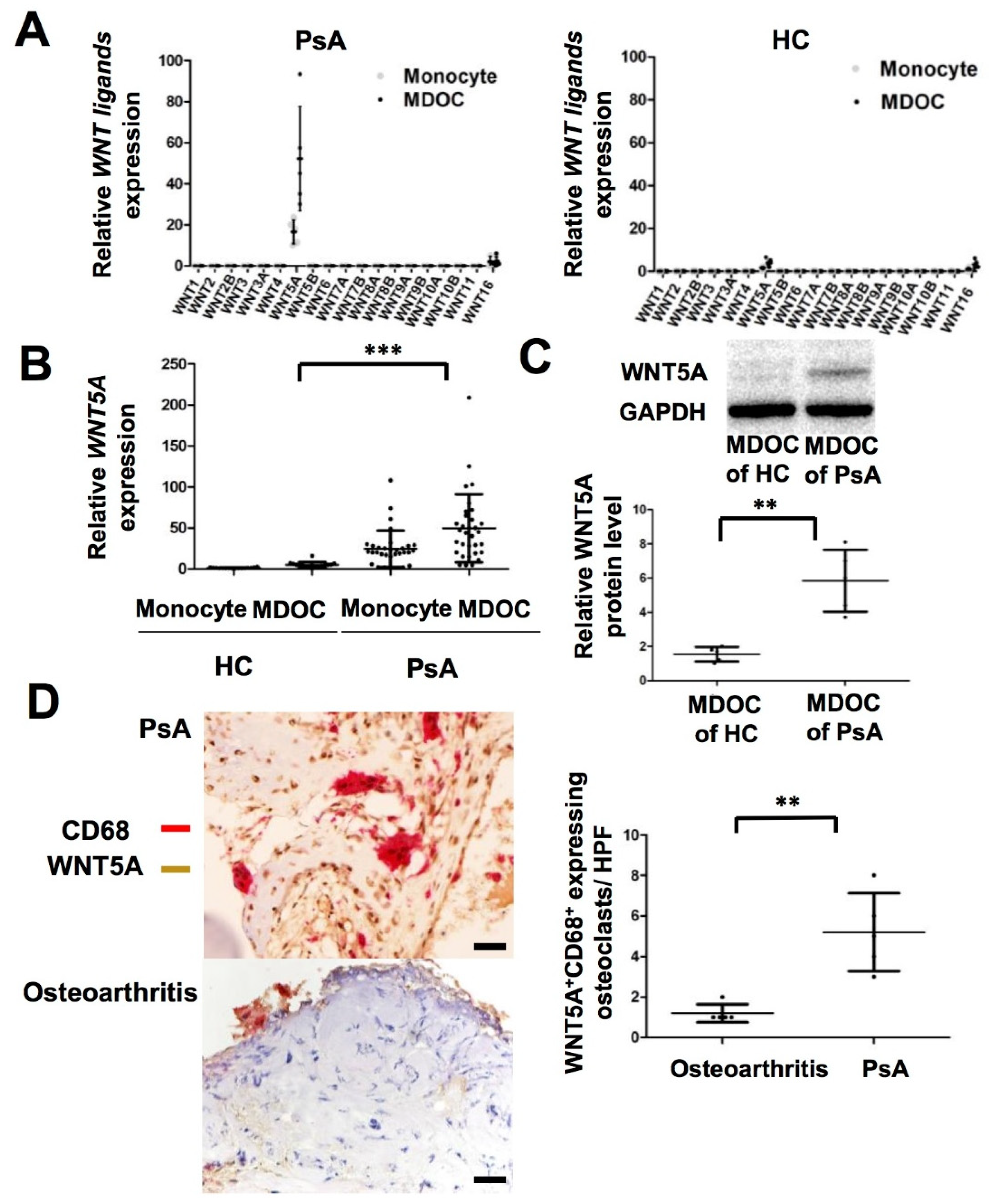
2.2. Increased Transcription and Translation of WNT5A in MDOC and Tissue Osteoclasts from Affected Joints in PsA Patients
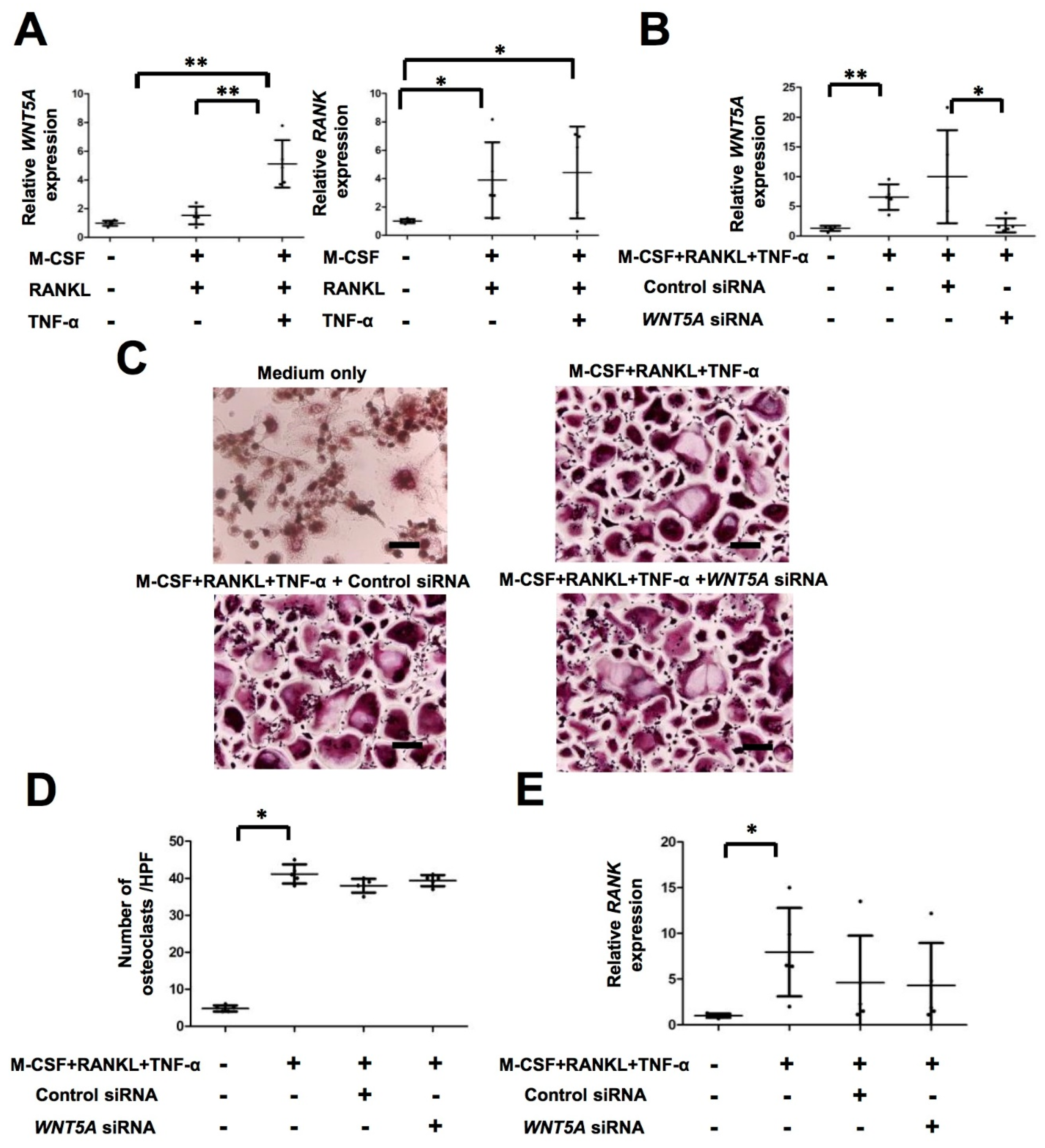
2.3. TNF-α Activates WNT5A Pathway, Which Is Independent of Osteoclastogenesis in PsA
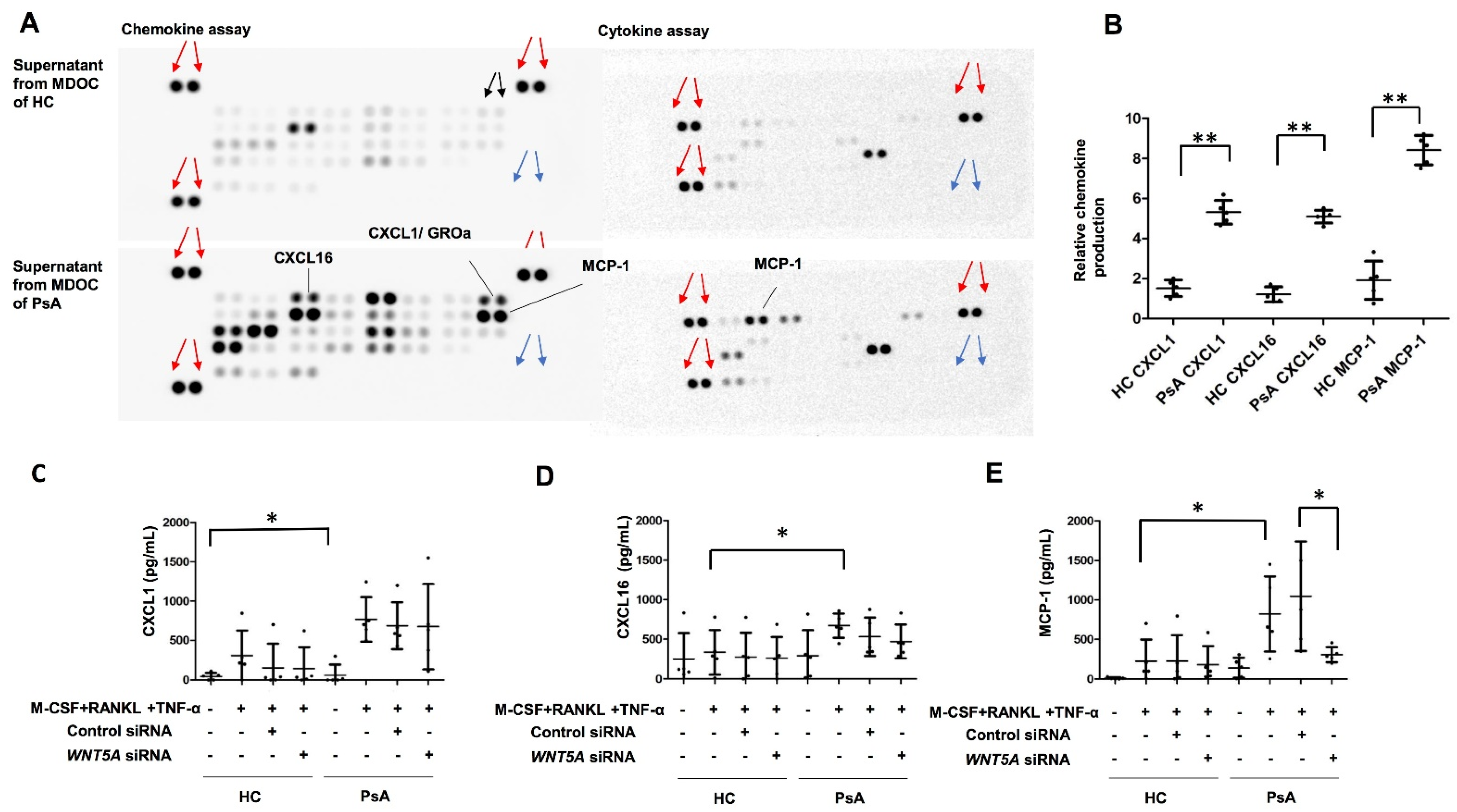
2.4. Selective Induction of MCP-1, but Not CXCL1 or CXCL16, by WNT5A in MDOC from PsA Patients
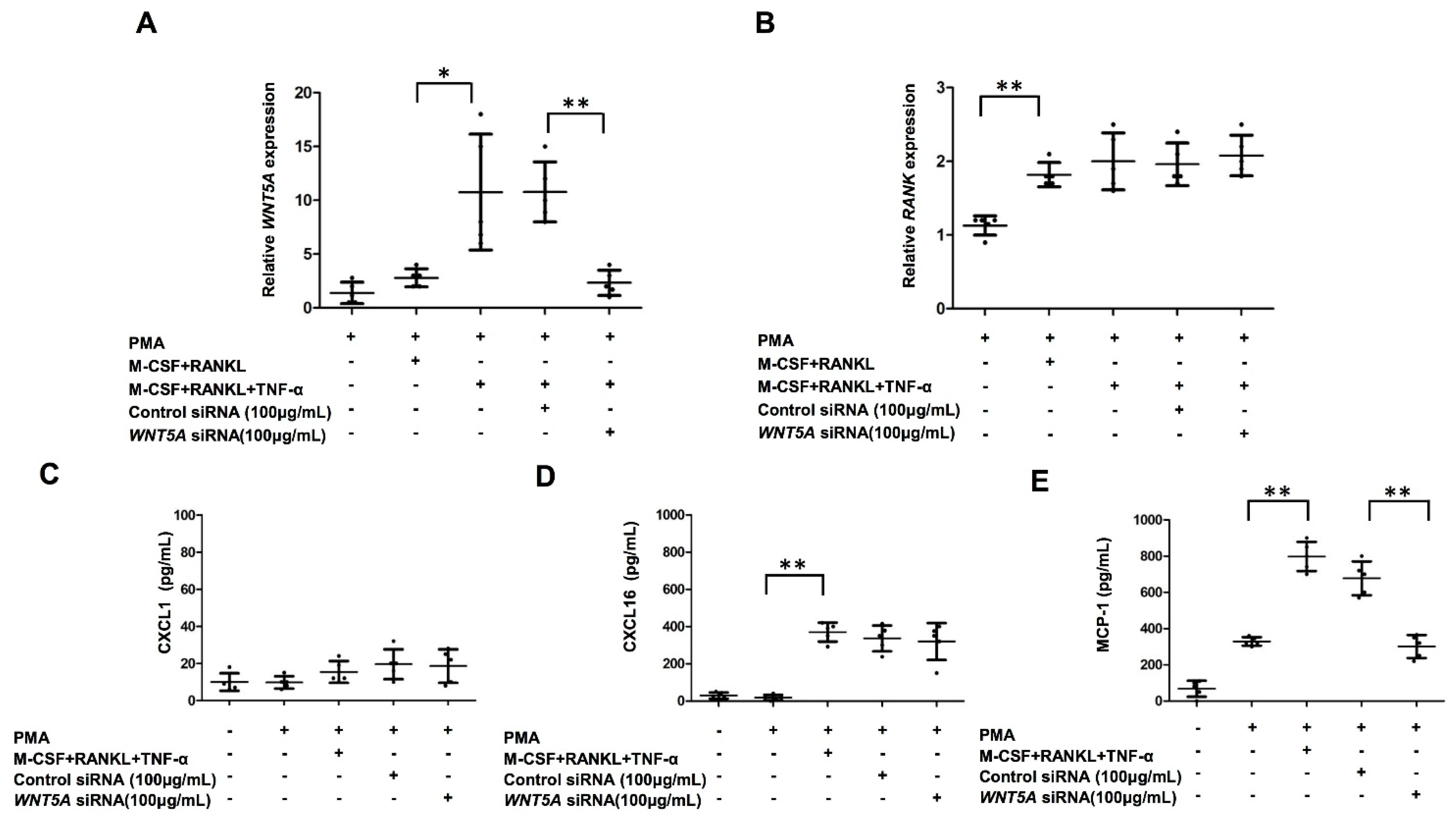
2.5. Increased Production of MCP-1 in THP-1-Cell-Derived Osteoclasts by TNF-α Treatment Was Abrogated by WNT5A Interference
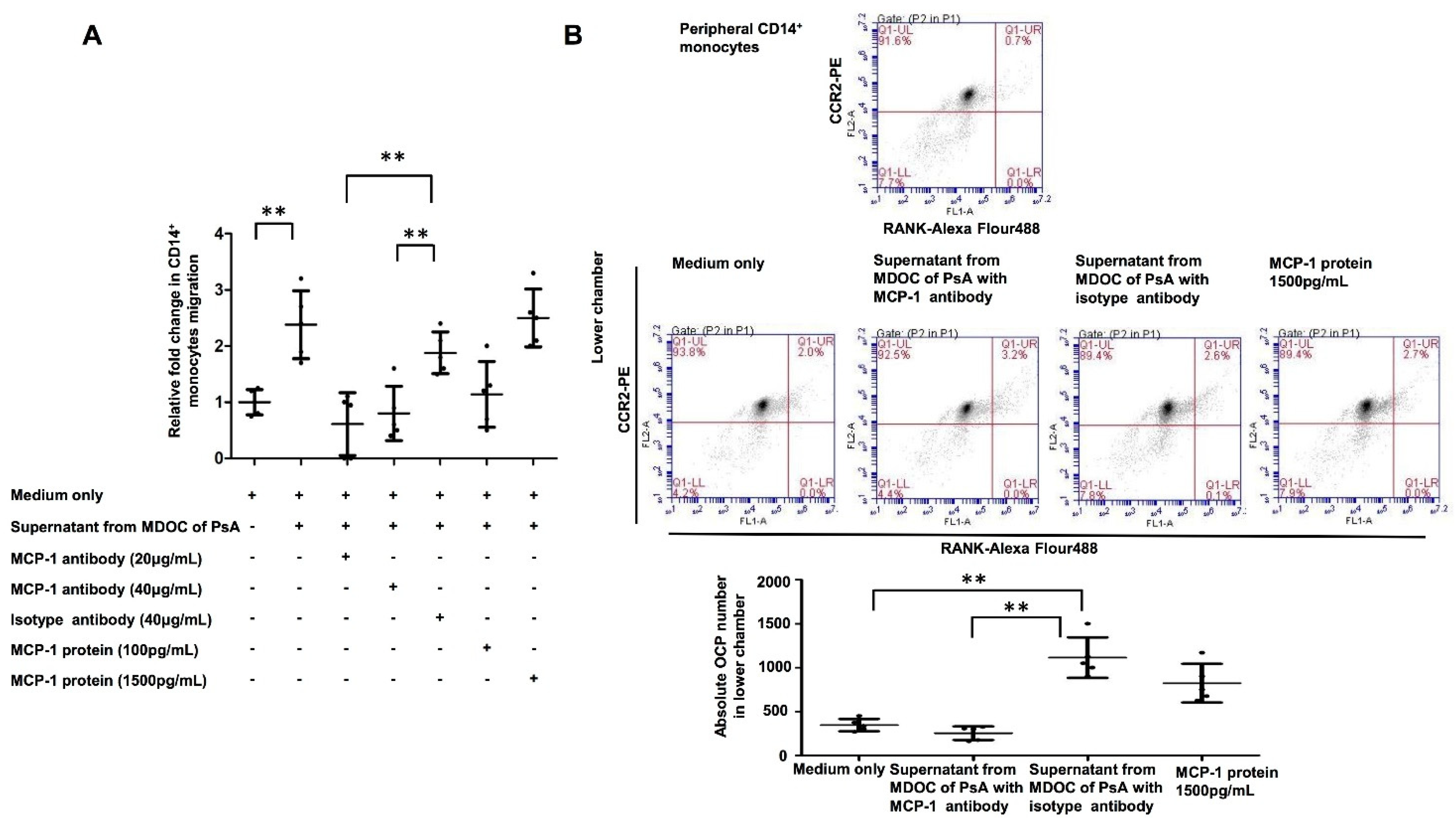
2.6. Increased OCP Recruitment Induced by Supernatants of MDOC in PsA Was Abrogated by MCP-1 Blocking
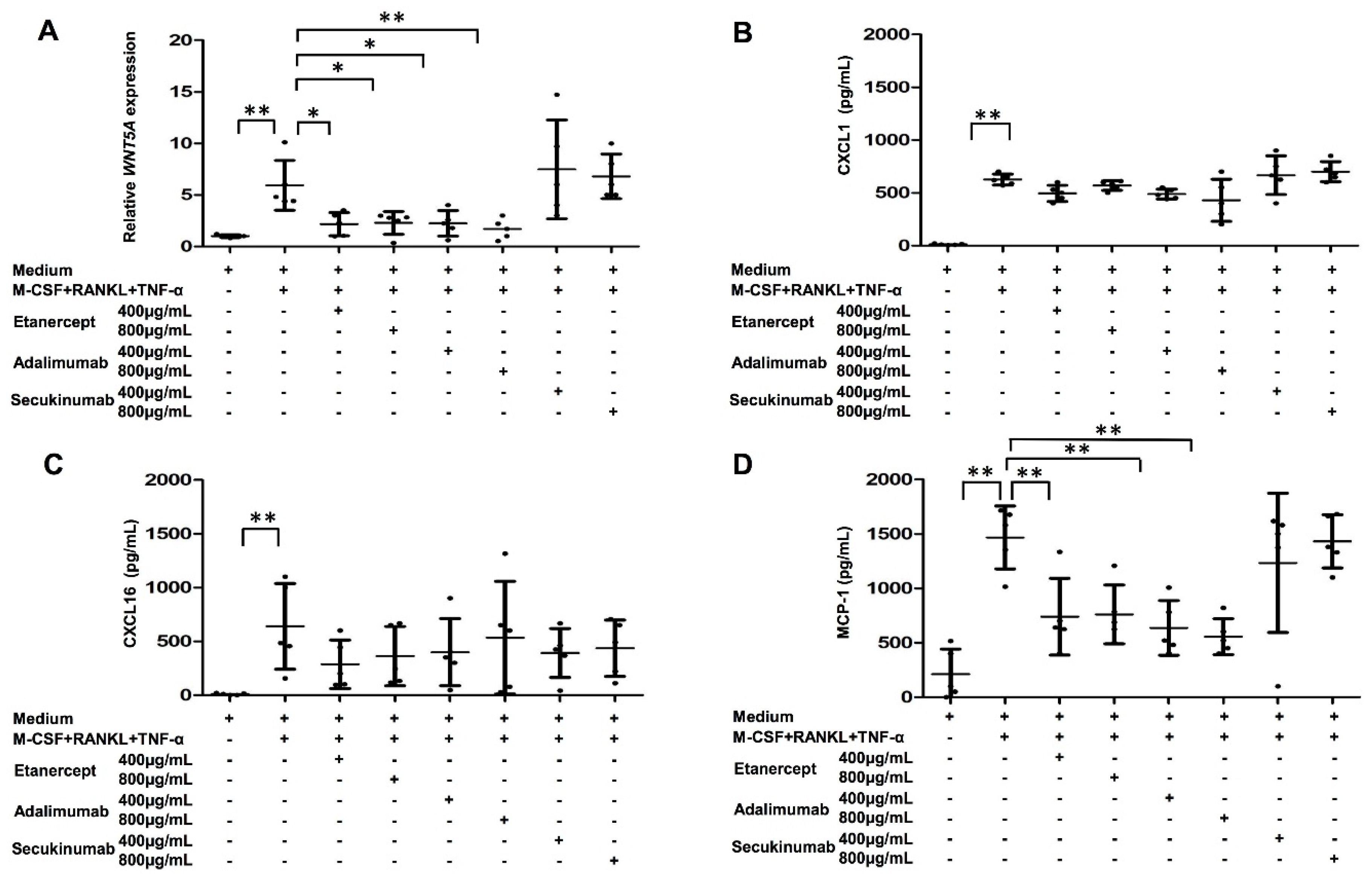
2.7. Both WNT5A Expression and MCP-1 Production in MDOC of PsA Patients Were Decreased by TNF-α Blockade
3. Discussion
4. Materials and Methods
4.1. Isolation of Human Circulatory CD14+ Monocytes to Profile WNT Ligands in Patients with PsA and HC
4.2. Differentiation of Osteoclasts (MDOC) from Human Circulatory CD14+ Monocytes
4.3. Differentiation of Osteoclasts from THP-1 Cells
4.4. Method for RNA Isolation and cDNA Synthesis
4.5. Western Blot Analysis for WNT5A in MDOC
4.6. Immunohistochemistry (IHC) for WNT5A+ and CD68+Expressing Osteoclasts in PsA-Affected Joints
4.7. RNA Interference against WNT5A in MDOC
4.8. Human Cytokine and Chemokine Antibody Array
4.9. Enzyme-Linked Immunosorbent Assay (ELISA)
4.10. Cell Migration
4.11. Flow Cytometry for RANK in OCP
4.12. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchlin, C.T.; Colbert, R.A.; Gladman, D.D. Psoriatic Arthritis. N. Engl. J. Med. 2017, 376, 957–970. [Google Scholar] [CrossRef] [Green Version]
- Veale, D.J.; Fearon, U. The pathogenesis of psoriatic arthritis. Lancet 2018, 391, 2273–2284. [Google Scholar] [CrossRef]
- Anandarajah, A.P.; Ritchlin, C.T. The diagnosis and treatment of early psoriatic arthritis. Nature reviews. Rheumatology 2009, 5, 634–641. [Google Scholar] [PubMed]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Massey, H.M.; Flanagan, A.M. Human osteoclasts derive from CD14-positive monocytes. Br. J. Haematol. 1999, 106, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Takayanagi, H. Osteoimmunology: Crosstalk between the immune and bone systems. J. Clin. Immunol. 2009, 29, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.H.; Mensah, K.A.; Schwarz, E.M.; Ju, Y.; Takahata, M.; Feng, C.; McMahon, L.A.; Hicks, D.G.; Panepento, B.; Keng, P.C.; et al. Regulation of human osteoclast development by dendritic cell-specific transmembrane protein (DC-STAMP). J. Bone Miner. Res. 2012, 27, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Ritchlin, C.T.; Haas-Smith, S.A.; Li, P.; Hicks, D.G.; Schwarz, E.M. Mechanisms of TNF-alpha- and RANKL-mediated osteoclastogenesis and bone resorption in psoriatic arthritis. J. Clin. Investig. 2003, 111, 821–831. [Google Scholar] [CrossRef]
- Geissmann, F.; Jung, S.; Littman, D.R. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity 2003, 19, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Komano, Y.; Nanki, T.; Hayashida, K.; Taniguchi, K.; Miyasaka, N. Identification of a human peripheral blood monocyte subset that differentiates into osteoclasts. Arthritis Res. Ther. 2006, 8, R152. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, R.; Kukita, T.; Ichigi, Y.; Takigawa, I.; Qu, P.F.; Funakubo, N.; Miyamoto, H.; Nonaka, K.; Kukita, A. Characterization and identification of subpopulations of mononuclear preosteoclasts induced by TNF-alpha in combination with TGF-beta in rats. PLoS ONE 2012, 7, e47930. [Google Scholar] [CrossRef] [Green Version]
- Ross, E.L.; D’Cruz, D.; Morrow, W.J. Localized monocyte chemotactic protein-1 production correlates with T cell infiltration of synovium in patients with psoriatic arthritis. J. Rheumatol. 2000, 27, 2432–2443. [Google Scholar]
- Haringman, J.J.; Ludikhuize, J.; Tak, P.P. Chemokines in joint disease: The key to inflammation? Ann. Rheum. Dis. 2004, 63, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Lerner, U.H.; Ohlsson, C. The WNT system: Background and its role in bone. J. Intern. Med. 2015, 277, 630–649. [Google Scholar] [CrossRef]
- Maeda, K.; Kobayashi, Y.; Udagawa, N.; Uehara, S.; Ishihara, A.; Mizoguchi, T.; Kikuchi, Y.; Takada, I.; Kato, S.; Kani, S.; et al. Wnt5a-Ror2 signaling between osteoblast-lineage cells and osteoclast precursors enhances osteoclastogenesis. Nat. Med. 2012, 18, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Pederson, L.; Ruan, M.; Westendorf, J.J.; Khosla, S.; Oursler, M.J. Regulation of bone formation by osteoclasts involves Wnt/BMP signaling and the chemokine sphingosine-1-phosphate. Proc. Natl. Acad. Sci. USA 2008, 105, 20764–20769. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Zeve, D.; Suh, J.M.; Wang, X.; Du, Y.; Zerwekh, J.E.; Dechow, P.C.; Graff, J.M.; Wan, Y. Biphasic and dosage-dependent regulation of osteoclastogenesis by beta-catenin. Mol. Cell. Biol. 2011, 31, 4706–4719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, F.; Oguma, J.; Brown, A.M.; Laurence, J. Noncanonical Wnt signaling promotes osteoclast differentiation and is facilitated by the human immunodeficiency virus protease inhibitor ritonavir. Biochem. Biophys. Res. Commun. 2012, 417, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Sen, M.; Lauterbach, K.; El-Gabalawy, H.; Firestein, G.S.; Corr, M.; Carson, D.A. Expression and function of wingless and frizzled homologs in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2000, 97, 2791–2796. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Mauro, T.M.; Li, Z. The pathological role of Wnt5a in psoriasis and psoriatic arthritis. J. Cell Mol. Med. 2019, 23, 5876–5883. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.H.; Chuang, H.Y.; Ho, J.C.; Lee, C.H.; Hsiao, C.C. Treatment with TNF-alpha inhibitor rectifies M1 macrophage polarization from blood CD14+ monocytes in patients with psoriasis independent of STAT1 and IRF-1 activation. J. Dermatol. Sci. 2018, 91, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Zhan, Z.; Li, S.; Zheng, Z.; Wang, T.; Zhang, K.; Pan, H.; Li, Z.; Zhang, N.; et al. Inflammation Intensity-Dependent Expression of Osteoinductive Wnt Proteins Is Critical for Ectopic New Bone Formation in Ankylosing Spondylitis. Arthritis Rheumatol. 2018, 70, 1056–1070. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, C.L.; Li, R.M.; Hui, T.Q.; Su, Y.Y.; Yuan, Q.; Zhou, X.D.; Ye, L. Wnt5a promotes inflammatory responses via nuclear factor kappaB (NF-kappaB) and mitogen-activated protein kinase (MAPK) pathways in human dental pulp cells. J. Biol. Chem. 2014, 289, 21028–21039. [Google Scholar] [CrossRef] [Green Version]
- Bougault, C.; Briolay, A.; Boutet, M.A.; Pilet, P.; Delplace, S.; Le Goff, B.; Guicheux, J.; Blanchard, F.; Magne, D. Wnt5a is expressed in spondyloarthritis and exerts opposite effects on enthesis and bone in murine organ and cell cultures. Transl. Res. 2015, 166, 627–638. [Google Scholar] [CrossRef]
- Uehara, S.; Udagawa, N.; Mukai, H.; Ishihara, A.; Maeda, K.; Yamashita, T.; Murakami, K.; Nishita, M.; Nakamura, T.; Kato, S.; et al. Protein kinase N3 promotes bone resorption by osteoclasts in response to Wnt5a-Ror2 signaling. Sci. Signal. 2017, 10, eaan0023. [Google Scholar] [CrossRef] [Green Version]
- Uehara, S.; Udagawa, N.; Kobayashi, Y. Non-canonical Wnt signals regulate cytoskeletal remodeling in osteoclasts. Cell. Mol. Life Sci. CMLS 2018, 75, 3683–3692. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Bu, X.; Wang, W.; Ma, T.; Ma, H. GEC-derived SFRP5 inhibits Wnt5a-induced macrophage chemotaxis and activation. PLoS ONE 2014, 9, e85058. [Google Scholar] [CrossRef]
- Konig, A.; Krenn, V.; Toksoy, A.; Gerhard, N.; Gillitzer, R. Mig, GRO alpha and RANTES messenger RNA expression in lining layer, infiltrates and different leucocyte populations of synovial tissue from patients with rheumatoid arthritis, psoriatic arthritis and osteoarthritis. Virchows Arch. 2000, 436, 449–458. [Google Scholar] [PubMed]
- Chandran, V.; Abji, F.; Perruccio, A.V.; Gandhi, R.; Li, S.; Cook, R.J.; Gladman, D.D. Serum-based soluble markers differentiate psoriatic arthritis from osteoarthritis. Ann. Rheum. Dis. 2019, 78, 796–801. [Google Scholar] [CrossRef]
- Steel, K.J.A.; Srenathan, U.; Ridley, M.; Durham, L.E.; Wu, S.Y.; Ryan, S.E.; Hughes, C.D.; Chan, E.; Kirkham, B.W.; Taams, L.S. Polyfunctional, Proinflammatory, Tissue-Resident Memory Phenotype and Function of Synovial Interleukin-17A+CD8+ T Cells in Psoriatic Arthritis. Arthritis Rheumatol. 2020, 72, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardaway, A.L.; Herroon, M.K.; Rajagurubandara, E.; Podgorski, I. Marrow adipocyte-derived CXCL1 and CXCL2 contribute to osteolysis in metastatic prostate cancer. Clin. Exp. Metastasis 2015, 32, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Hounoki, H.; Sugiyama, E.; Mohamed, S.G.; Shinoda, K.; Taki, H.; Abdel-Aziz, H.O.; Maruyama, M.; Kobayashi, M.; Miyahara, T. Activation of peroxisome proliferator-activated receptor gamma inhibits TNF-alpha-mediated osteoclast differentiation in human peripheral monocytes in part via suppression of monocyte chemoattractant protein-1 expression. Bone 2008, 42, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, P.; Wang, C.Y.; Stashenko, P.; Lee, S.K.; Lorenzo, J.A.; Graves, D.T. Monocyte chemoattractant protein-1 expression and monocyte recruitment in osseous inflammation in the mouse. Endocrinology 1995, 136, 2752–2759. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon. Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Cambre, I.; Gaublomme, D.; Burssens, A.; Jacques, P.; Schryvers, N.; De Muynck, A.; Meuris, L.; Lambrecht, S.; Carter, S.; de Bleser, P.; et al. Mechanical strain determines the site-specific localization of inflammation and tissue damage in arthritis. Nat. Commun. 2018, 9, 4613. [Google Scholar] [CrossRef] [Green Version]
- Sumi, K.; Abe, T.; Kunimatsu, R.; Oki, N.; Tsuka, Y.; Awada, T.; Nakajima, K.; Ando, K.; Tanimoto, K. The effect of mesenchymal stem cells on chemotaxis of osteoclast precursor cells. J. Oral. Sci 2018, 60, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Schett, G.; Elewaut, D.; McInnes, I.B.; Dayer, J.M.; Neurath, M.F. How cytokine networks fuel inflammation: Toward a cytokine-based disease taxonomy. Nat. Med. 2013, 19, 822–824. [Google Scholar] [CrossRef] [PubMed]
- Anandarajah, A.P.; Schwarz, E.M.; Totterman, S.; Monu, J.; Feng, C.Y.; Shao, T.; Haas-Smith, S.A.; Ritchlin, C.T. The effect of etanercept on osteoclast precursor frequency and enhancing bone marrow oedema in patients with psoriatic arthritis. Ann. Rheum. Dis. 2008, 67, 296–301. [Google Scholar] [CrossRef]
- Nielsen, M.A.; Lomholt, S.; Mellemkjaer, A.; Andersen, M.N.; Buckley, C.D.; Kragstrup, T.W. Responses to Cytokine Inhibitors Associated with Cellular Composition in Models of Immune-Mediated Inflammatory Arthritis. ACR Open Rheumatol. 2020, 2, 3–10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients with PsA (n = 32) | HC (n = 16) | |
|---|---|---|
| Age (years) | 48.3 ± 11.0 | 46.4 ± 12.5 |
| Female sex (no. (%)) | 16 (50%) | 8 (50%) |
| Weight (kg) | 71.6 ± 11.0 | 69.3 ± 13.6 |
| Psoriasis (years) | 15.3 ± 9.3 | |
| Psoriatic arthritis (years) | 9.3 ± 7.9 | |
| Previous drug use | ||
| Anti-TNF drugs, anti-IL-12/23, or anti-IL-17 (no. (%)) | 4 (10) | |
| Use of methotrexate (no. (%)) | 21/32 (65.6) | |
| Use of leflunomide, (no. (%)) | 10 (25) | |
| Use of NSAID, (no. (%)) | 38 (95) | |
| Patients with specific disease characteristics | ||
| PASI | 12.9 ± 8.8 | |
| Peripheral arthritis, (no. (%)) | 32 (100) | |
| Peripheral and axil arthritis, (no. (%)) | 10 (31.3) | |
| Dactylitis, (no. (%)) | 13/32 (40.6) | |
| Enthesitis, (no. (%)) | 20/32 (62.5) | |
| Tender-joint count (of 78 joints) | 9.9 ± 7.6 | |
| Swollen-joint count (of 76 joints) | 6.0 ± 6.7 | |
| Uveitis, (no. (%)) | 2 (6.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-H.; Ho, J.-C.; Li, S.-C.; Cheng, Y.-W.; Hsu, C.-Y.; Chou, W.-Y.; Hsiao, C.-C.; Lee, C.-H. TNF-α Activating Osteoclasts in Patients with Psoriatic Arthritis Enhances the Recruitment of Osteoclast Precursors: A Plausible Role of WNT5A-MCP-1 in Osteoclast Engagement in Psoriatic Arthritis. Int. J. Mol. Sci. 2022, 23, 921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020921
Lin S-H, Ho J-C, Li S-C, Cheng Y-W, Hsu C-Y, Chou W-Y, Hsiao C-C, Lee C-H. TNF-α Activating Osteoclasts in Patients with Psoriatic Arthritis Enhances the Recruitment of Osteoclast Precursors: A Plausible Role of WNT5A-MCP-1 in Osteoclast Engagement in Psoriatic Arthritis. International Journal of Molecular Sciences. 2022; 23(2):921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020921
Chicago/Turabian StyleLin, Shang-Hung, Ji-Chen Ho, Sung-Chou Li, Yu-Wen Cheng, Chung-Yuan Hsu, Wen-Yi Chou, Chang-Chun Hsiao, and Chih-Hung Lee. 2022. "TNF-α Activating Osteoclasts in Patients with Psoriatic Arthritis Enhances the Recruitment of Osteoclast Precursors: A Plausible Role of WNT5A-MCP-1 in Osteoclast Engagement in Psoriatic Arthritis" International Journal of Molecular Sciences 23, no. 2: 921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020921