
May Sulfonamide Inhibitors of Carbonic Anhydrases from Mammaliicoccus sciuri Prevent Antimicrobial Resistance Due to Gene Transfer to Other Harmful Staphylococci?


Abstract
:1. Introduction
2. Results and Discussion
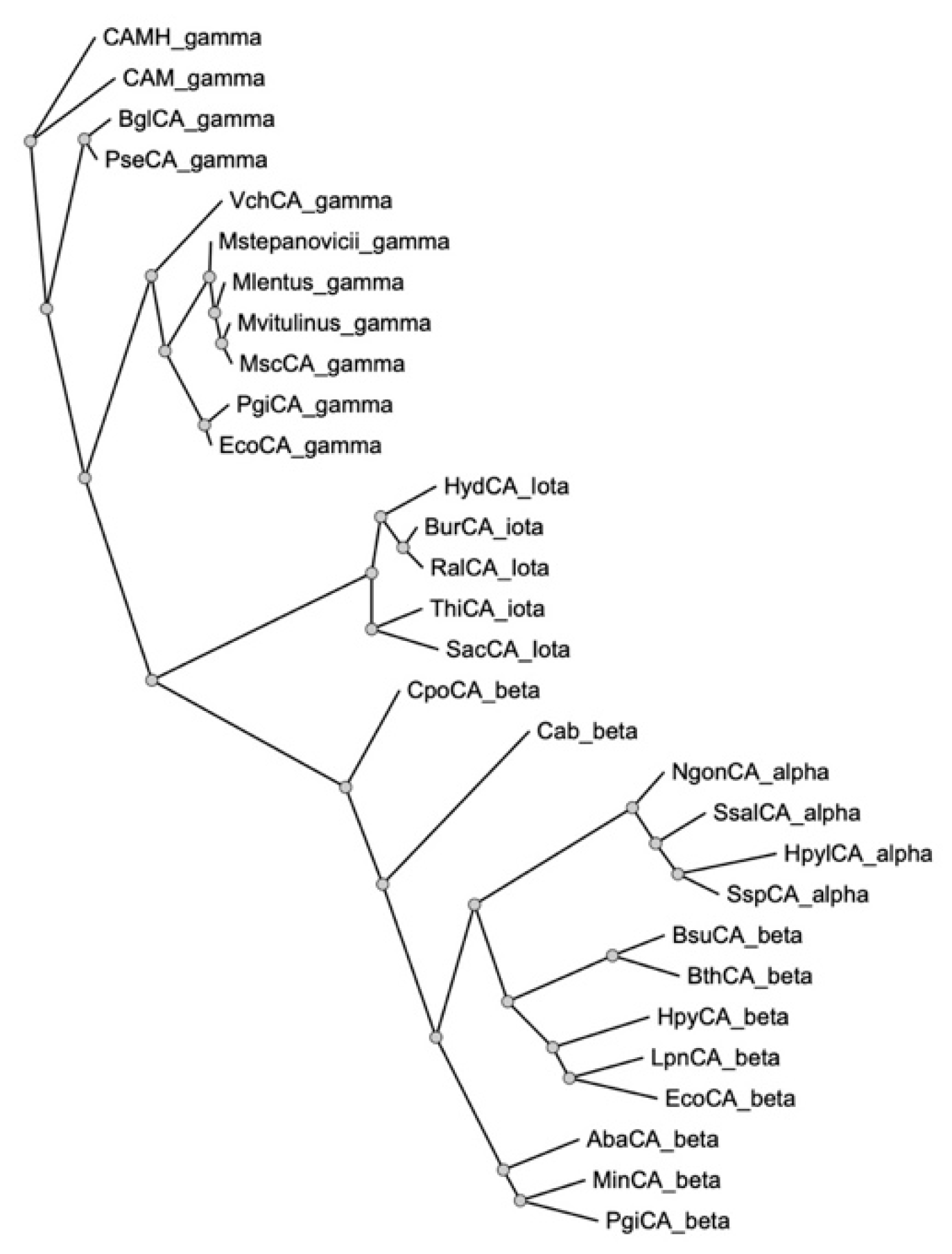
2.1. Gene Identification, Sequence Alignment, and Phylogenetic Analysis of γ-CA Encoded by the M. sciuri Genome
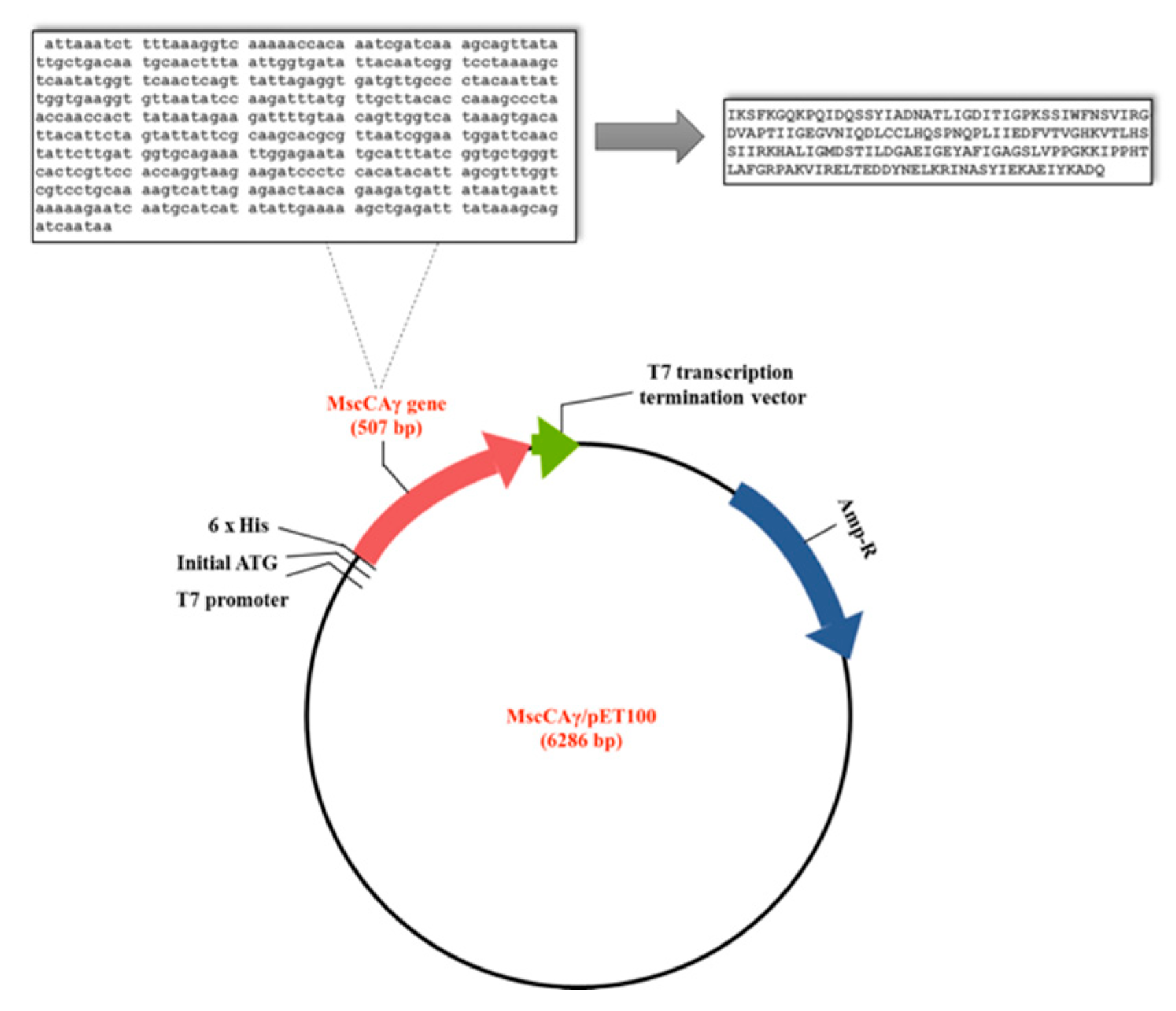
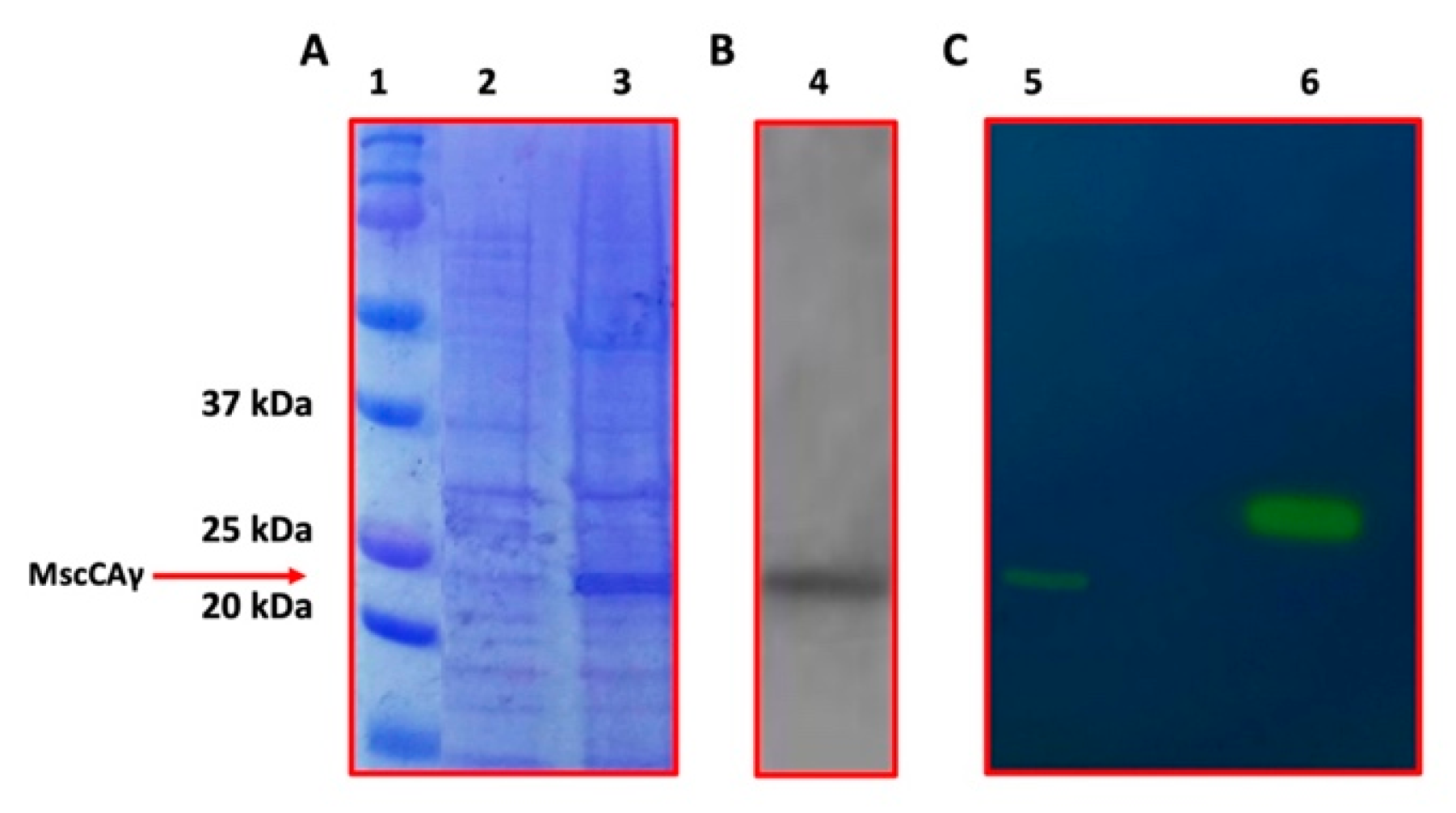
2.2. The Expression of the MscCAγ and Its Subsequent Purification
2.3. Kinetic Parameters Determined Using the Stopped-Flow Method
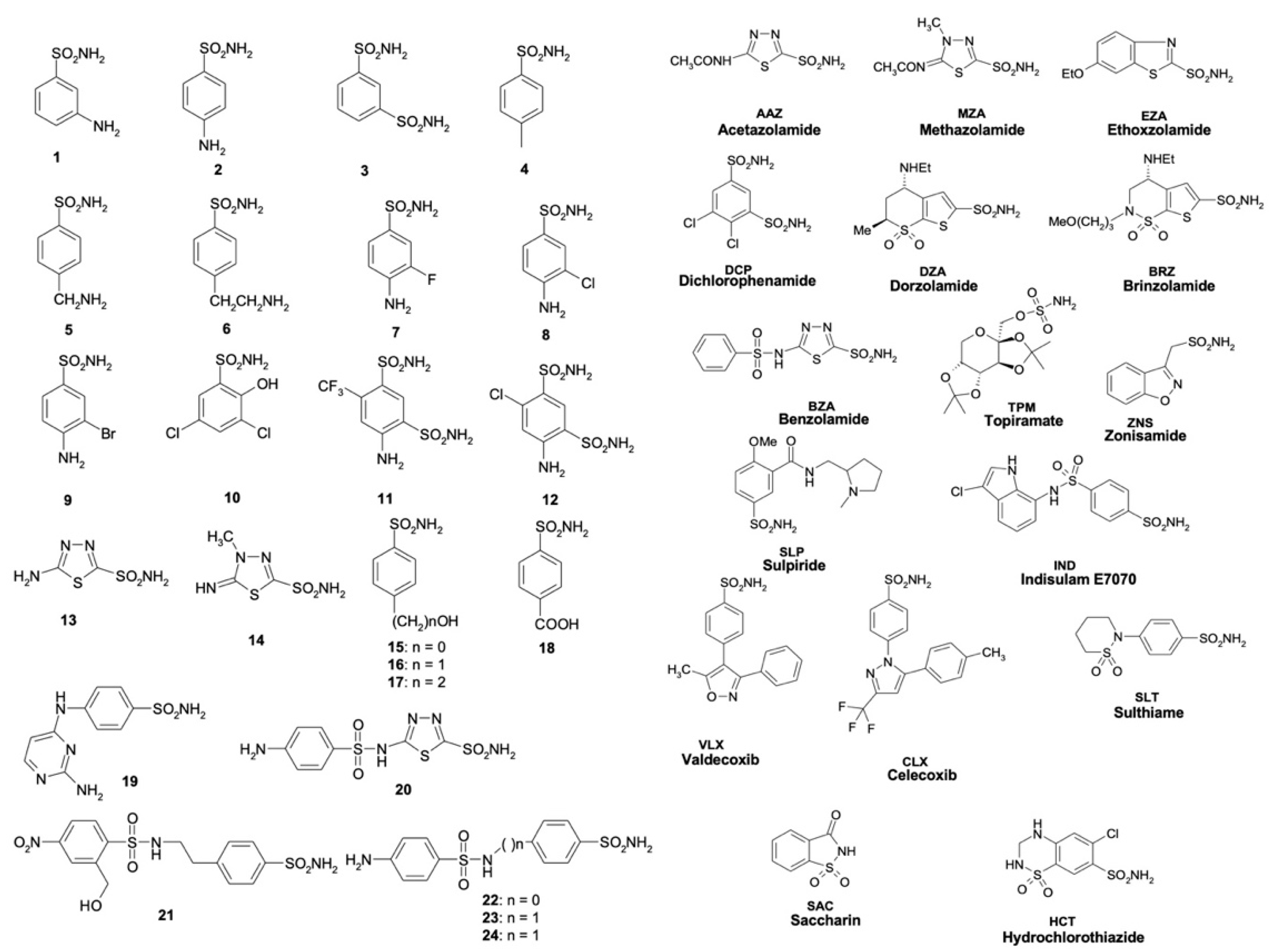
2.4. Sulfonamide Inhibition Profile
3. Materials and Methods
3.1. Bacterial Species, Vectors, and Chemicals
3.2. Screening of Protein Databases, Sequence Analysis, and Phylogenetic Analysis
3.3. Cloning, Expression of the MscCAγ, and Its Subsequent Purification
3.4. Enzyme Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bierowiec, K.; Korzeniowska-Kowal, A.; Wzorek, A.; Rypula, K.; Gamian, A. Prevalence of Staphylococcus Species Colonization in Healthy and Sick Cats. Biomed. Res. Int. 2019, 2019, 4360525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berube, B.J.; Bubeck Wardenburg, J. Staphylococcus aureus alpha-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [Green Version]
- Von Hoven, G.; Qin, Q.; Neukirch, C.; Husmann, M.; Hellmann, N. Staphylococcus aureus alpha-toxin: Small pore, large consequences. Biol. Chem. 2019, 400, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Lee, A.; Hong, J.; Jo, W.Y.; Cho, O.H.; Kim, S.; Bae, I.G. Staphylococcus saprophyticus Bacteremia originating from Urinary Tract Infections: A Case Report and Literature Review. Infect. Chemother. 2016, 48, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.R.; Jiang, J.H.; Hassan, K.A.; Elbourne, L.D.; Tuck, K.L.; Paulsen, I.T.; Peleg, A.Y. Insights on virulence from the complete genome of Staphylococcus capitis. Front. Microbiol. 2015, 6, 980. [Google Scholar] [CrossRef] [Green Version]
- Severn, M.M.; Horswill, A.R. Staphylococcus epidermidis and its dual lifestyle in skin health and infection. Nat. Rev. Microbiol. 2022, 1–15. [Google Scholar] [CrossRef]
- Kloos, W.E.; Schleifer, K.H. Staphylococcus-Auricularis Sp-Nov—An Inhabitant of the Human External Ear. Int. J. Syst. Bacteriol. 1983, 33, 9–14. [Google Scholar] [CrossRef]
- Wald, R.; Hess, C.; Urbantke, V.; Wittek, T.; Baumgartner, M. Characterization of Staphylococcus Species Isolated from Bovine Quarter Milk Samples. Animals 2019, 9, 200. [Google Scholar] [CrossRef] [Green Version]
- Antonello, R.M.; Riccardi, N. How we deal with Staphylococcus aureus (MSSA, MRSA) central nervous system infections. Front. Biosci. 2022, 14, 1. [Google Scholar] [CrossRef]
- Cassat, J.E.; Thomsen, I. Staphylococcus aureus infections in children. Curr. Opin. Infect. Dis. 2021, 34, 510–518. [Google Scholar] [CrossRef]
- Kimmig, A.; Hagel, S.; Weis, S.; Bahrs, C.; Loffler, B.; Pletz, M.W. Management of Staphylococcus aureus Bloodstream Infections. Front. Med. 2020, 7, 616524. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, O.; Marasca, S.; Candelora, M.; Rizzetto, G.; Radi, G.; Molinelli, E.; Brescini, L.; Cirioni, O.; Offidani, A. Methicillin-resistant Staphylococcus aureus as a cause of chronic wound infections: Alternative strategies for management. AIMS Microbiol. 2022, 8, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Bakthavatchalam, Y.D.; Rao, S.V.; Isaac, B.; Manesh, A.; Nambi, S.; Swaminathan, S.; Nagvekar, V.; Nangia, V.; John, P.V.; Veeraraghavan, B. A comparative assessment of clinical, pharmacological and antimicrobial profile of novel anti-methicillin-resistant Staphylococcus aureus agent levonadifloxacin: Therapeutic role in nosocomial and community infections. Indian J. Med. Microbiol. 2019, 37, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Madhaiyan, M.; Wirth, J.S.; Saravanan, V.S. Phylogenomic analyses of the Staphylococcaceae family suggest the reclassification of five species within the genus Staphylococcus as heterotypic synonyms, the promotion of five subspecies to novel species, the taxonomic reassignment of five Staphylococcus species to Mammaliicoccus gen. nov., and the formal assignment of Nosocomiicoccus to the family Staphylococcaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 5926–5936. [Google Scholar]
- Van der Veken, D.; Hollanders, C.; Verce, M.; Michiels, C.; Ballet, S.; Weckx, S.; Leroy, F. Genome-Based Characterization of a Plasmid-Associated Micrococcin P1 Biosynthetic Gene Cluster and Virulence Factors in Mammaliicoccus sciuri IMDO-S72. Appl. Environ. Microbiol. 2022, 88, e0208821. [Google Scholar] [CrossRef]
- Nemeghaire, S.; Argudin, M.A.; Fessler, A.T.; Hauschild, T.; Schwarz, S.; Butaye, P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Vet. Microbiol. 2014, 171, 342–356. [Google Scholar] [CrossRef]
- Lienen, T.; Schnitt, A.; Hammerl, J.A.; Maurischat, S.; Tenhagen, B.A. Mammaliicoccus spp. from German Dairy Farms Exhibit a Wide Range of Antimicrobial Resistance Genes and Non-Wildtype Phenotypes to Several Antibiotic Classes. Biology 2022, 11, 152. [Google Scholar] [CrossRef]
- Abutaleb, N.S.; Elhassanny, A.E.M.; Seleem, M.N. In vivo efficacy of acetazolamide in a mouse model of Neisseria gonorrhoeae infection. Microb. Pathog. 2022, 164, 105454. [Google Scholar] [CrossRef]
- Buzas, G.M. Helicobacter pylori—2010. Orv. Hetil. 2010, 151, 2003–2010. [Google Scholar] [CrossRef]
- Buzas, G.M.; Supuran, C.T. The history and rationale of using carbonic anhydrase inhibitors in the treatment of peptic ulcers. In memoriam Ioan Puscas (1932–2015). J. Enzym. Inhib. Med. Chem. 2016, 31, 527–533. [Google Scholar] [CrossRef]
- Kaur, J.; Cao, X.; Abutaleb, N.S.; Elkashif, A.; Graboski, A.L.; Krabill, A.D.; AbdelKhalek, A.H.; An, W.; Bhardwaj, A.; Seleem, M.N.; et al. Optimization of Acetazolamide-Based Scaffold as Potent Inhibitors of Vancomycin-Resistant Enterococcus. J. Med. Chem. 2020, 63, 9540–9562. [Google Scholar] [CrossRef] [PubMed]
- Lotlikar, S.R.; Kayastha, B.B.; Vullo, D.; Khanam, S.S.; Braga, R.E.; Murray, A.B.; McKenna, R.; Supuran, C.T.; Patrauchan, M.A. Pseudomonas aeruginosa beta-carbonic anhydrase, psCA1, is required for calcium deposition and contributes to virulence. Cell Calcium 2019, 84, 102080. [Google Scholar] [CrossRef] [PubMed]
- Modak, J.K.; Tikhomirova, A.; Gorrell, R.J.; Rahman, M.M.; Kotsanas, D.; Korman, T.M.; Garcia-Bustos, J.; Kwok, T.; Ferrero, R.L.; Supuran, C.T.; et al. Anti-Helicobacter pylori activity of ethoxzolamide. J. Enzym. Inhib. Med. Chem. 2019, 34, 1660–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimori, I.; Minakuchi, T.; Vullo, D.; Scozzafava, A.; Supuran, C.T. Inhibition studies of the beta-carbonic anhydrases from the bacterial pathogen Salmonella enterica serovar Typhimurium with sulfonamides and sulfamates. Bioorg. Med. Chem. 2011, 19, 5023–5030. [Google Scholar] [CrossRef] [PubMed]
- Rollenhagen, C.; Bumann, D. Salmonella enterica highly expressed genes are disease specific. Infect. Immun. 2006, 74, 1649–1660. [Google Scholar] [CrossRef] [Green Version]
- Ronci, M.; Del Prete, S.; Puca, V.; Carradori, S.; Carginale, V.; Muraro, R.; Mincione, G.; Aceto, A.; Sisto, F.; Supuran, C.T.; et al. Identification and characterization of the alpha-CA in the outer membrane vesicles produced by Helicobacter pylori. J. Enzym. Inhib. Med. Chem. 2019, 34, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Nishimori, I.; Minakuchi, T.; Scozzafava, A.; Supuran, C.T. Inhibition studies with anions and small molecules of two novel beta-carbonic anhydrases from the bacterial pathogen Salmonella enterica serovar Typhimurium. Bioorg. Med. Chem. Lett. 2011, 21, 3591–3595. [Google Scholar] [CrossRef]
- Supuran, C.T. Emerging role of carbonic anhydrase inhibitors. Clin. Sci. 2021, 135, 1233–1249. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrase inhibitors: An update on experimental agents for the treatment and imaging of hypoxic tumors. Expert Opin. Investig. Drugs 2021, 30, 1197–1208. [Google Scholar] [CrossRef]
- Giovannuzzi, S.; Hewitt, C.S.; Nocentini, A.; Capasso, C.; Flaherty, D.P.; Supuran, C.T. Coumarins effectively inhibit bacterial alpha-carbonic anhydrases. J. Enzym. Inhib. Med. Chem. 2022, 37, 333–338. [Google Scholar] [CrossRef]
- Giovannuzzi, S.; Hewitt, C.S.; Nocentini, A.; Capasso, C.; Costantino, G.; Flaherty, D.P.; Supuran, C.T. Inhibition studies of bacterial alpha-carbonic anhydrases with phenols. J. Enzym. Inhib. Med. Chem. 2022, 37, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta- and gamma-carbonic anhydrases from Bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzym. Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Nocentini, A.; Supuran, C.T.; Capasso, C. An overview on the recently discovered iota-carbonic anhydrases. J. Enzym. Inhib. Med. Chem. 2021, 36, 1988–1995. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. New light on bacterial carbonic anhydrases phylogeny based on the analysis of signal peptide sequences. J. Enzym. Inhib. Med. Chem. 2016, 31, 1254–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T.; Capasso, C. An Overview of the Bacterial Carbonic Anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T.; Capasso, C. Biomedical applications of prokaryotic carbonic anhydrases. Expert Opin. Ther. Pat. 2018, 28, 745–754. [Google Scholar] [CrossRef]
- De Luca, V.; Carginale, V.; Supuran, C.T.; Capasso, C. The gram-negative bacterium Escherichia coli as a model for testing the effect of carbonic anhydrase inhibition on bacterial growth. J. Enzym. Inhib. Med. Chem. 2022, 37, 2092–2098. [Google Scholar] [CrossRef]
- Urbanski, L.J.; Bua, S.; Angeli, A.; Kuuslahti, M.; Hytonen, V.P.; Supuran, C.T.; Parkkila, S. Sulphonamide inhibition profile of Staphylococcus aureus beta-carbonic anhydrase. J. Enzym. Inhib. Med. Chem. 2020, 35, 1834–1839. [Google Scholar] [CrossRef]
- Urbanski, L.J.; Vullo, D.; Parkkila, S.; Supuran, C.T. An anion and small molecule inhibition study of the beta-carbonic anhydrase from Staphylococcus aureus. J. Enzym. Inhib. Med. Chem. 2021, 36, 1088–1092. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Tripp, B.C.; Smith, K.; Ferry, J.G. Carbonic anhydrase: New insights for an ancient enzyme. J. Biol. Chem. 2001, 276, 48615–48618. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; Del Prete, S.; Supuran, C.T.; Capasso, C. Protonography, a new technique for the analysis of carbonic anhydrase activity. J. Enzym. Inhib. Med. Chem. 2015, 30, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; Iandolo, E.; Supuran, C.T.; Capasso, C. Protonography, a powerful tool for analyzing the activity and the oligomeric state of the gamma-carbonic anhydrase identified in the genome of Porphyromonas gingivalis. Bioorg. Med. Chem. 2015, 23, 3747–3750. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; Supuran, C.T.; Capasso, C. Protonography, a technique applicable for the analysis of eta-carbonic anhydrase activity. J. Enzym. Inhib. Med. Chem. 2015, 30, 920–924. [Google Scholar] [CrossRef] [PubMed]
- Akocak, S.; Lolak, N.; Bua, S.; Nocentini, A.; Supuran, C.T. Activation of human alpha-carbonic anhydrase isoforms I, II, IV and VII with bis-histamine schiff bases and bis-spinaceamine substituted derivatives. J. Enzym. Inhib. Med. Chem. 2019, 34, 1193–1198. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T.; Capasso, C. Antibacterial carbonic anhydrase inhibitors: An update on the recent literature. Expert Opin. Ther. Pat. 2020, 30, 963–982. [Google Scholar] [CrossRef]
- Abutaleb, N.S.; Elkashif, A.; Flaherty, D.P.; Seleem, M.N. In Vivo Antibacterial Activity of Acetazolamide. Antimicrob. Agents Chemother. 2021, 65, e01715-20. [Google Scholar] [CrossRef]
- An, W.; Holly, K.J.; Nocentini, A.; Imhoff, R.D.; Hewitt, C.S.; Abutaleb, N.S.; Cao, X.; Seleem, M.N.; Supuran, C.T.; Flaherty, D.P. Structure-activity relationship studies for inhibitors for vancomycin-resistant Enterococcus and human carbonic anhydrases. J. Enzym. Inhib. Med. Chem. 2022, 37, 1838–1844. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Mount, D.W. Using the Basic Local Alignment Search Tool (BLAST). CSH Protoc. 2007, 2007, pdb.top17. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. The Clustal Omega Multiple Alignment Package. Methods Mol. Biol. 2021, 2231, 3–16. [Google Scholar]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Del Prete, S.; De Luca, V.; Nocentini, A.; Scaloni, A.; Mastrolorenzo, M.D.; Supuran, C.T.; Capasso, C. Anion Inhibition Studies of the Beta-Carbonic Anhydrase from Escherichia coli. Molecules 2020, 25, 2564. [Google Scholar] [CrossRef]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar] [CrossRef]
- Carta, F.; Temperini, C.; Innocenti, A.; Scozzafava, A.; Kaila, K.; Supuran, C.T. Polyamines inhibit carbonic anhydrases by anchoring to the zinc-coordinated water molecule. J. Med. Chem. 2010, 53, 5511–5522. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Acronym | Class | kcat (s−1) | kcat/Km (M−1 × s−1) | KI (Acetazolamide) (nM) |
|---|---|---|---|---|---|
| Homo sapiens | hCA I | α | 2.0 × 105 | 5.0 × 107 | 250 |
| hCA II | α | 1.4 × 106 | 1.5 × 108 | 12 | |
| Mammaliicoccus sciuri | MscCAβ (ex SauBCA) | β | 1.5 × 105 | 2.6 × 107 | 628 |
| MscCAγ | γ | 6.2 × 105 | 9.5 × 106 | 245 |
| Inhibitor | KI (nM) a | |||
|---|---|---|---|---|
| hCA I | hCA II | MscCAγ | MscCAβ (ex SauBCA) | |
| 1 | 28,000 | 300 | 797 | 355 |
| 2 | 25,000 | 240 | 888 | 409 |
| 3 | 79.0 | 8.0 | 552 | 95 |
| 4 | 78,500 | 320 | 647 | 83 |
| 5 | 25,000 | 170 | 354 | 193 |
| 6 | 21,000 | 160 | 176 | 253 |
| 7 | 8300 | 60.0 | 2010 | 93 |
| 8 | 9800 | 110 | 426 | 95 |
| 9 | 6500 | 40.0 | 478 | 75 |
| 10 | 7300 | 54.0 | 2429 | 202 |
| 11 | 5800 | 63.0 | 32,978 | 81 |
| 12 | 8400 | 75.0 | 17,200 | 79 |
| 13 | 8600 | 60.0 | 539 | 417 |
| 14 | 9300 | 19.0 | 820 | 553 |
| 15 | 5500 | 80.0 | 2931 | 619 |
| 16 | 9500 | 94.0 | 915 | 603 |
| 17 | 21,000 | 125 | 736 | 232 |
| 18 | 164 | 46.0 | 722 | 555 |
| 19 | 109 | 33.0 | 1893 | 909 |
| 20 | 6.0 | 2.0 | 1626 | 92 |
| 21 | 69.0 | 11.0 | 83.4 | 85 |
| 22 | 164 | 46.0 | 72.5 | 83 |
| 23 | 109 | 33.0 | 217 | 92 |
| 24 | 95.0 | 30.0 | 45.7 | 96 |
| AAZ | 250 | 12.0 | 245 | 628 |
| MZA | 50.0 | 14.0 | 94.5 | 863 |
| EZA | 25.0 | 8.0 | 96.6 | 698 |
| DCP | 1200 | 38.0 | 8580 | - |
| DZA | 50,000 | 9.0 | 43.8 | 909 |
| BRZ | 45,000 | 3.0 | 100 | 815 |
| BZA | 15.0 | 9.0 | 444 | 501 |
| TPM | 250 | 10.0 | 578 | 466 |
| ZNS | 56.0 | 35.0 | 923 | 4551 |
| SLP | 1200 | 40.0 | 2203 | 807 |
| IND | 31.0 | 15.0 | 369 | 588 |
| VLX | 54,000 | 43.0 | 1540 | 509 |
| CLX | 50,000 | 21.0 | 3024 | 871 |
| SLT | 374 | 9.0 | 405 | 824 |
| SAC | 18,540 | 5959 | 29,687 | 667 |
| HCT | 328 | 290 | 18,346 | 593 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, V.; Giovannuzzi, S.; Supuran, C.T.; Capasso, C. May Sulfonamide Inhibitors of Carbonic Anhydrases from Mammaliicoccus sciuri Prevent Antimicrobial Resistance Due to Gene Transfer to Other Harmful Staphylococci? Int. J. Mol. Sci. 2022, 23, 13827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213827
De Luca V, Giovannuzzi S, Supuran CT, Capasso C. May Sulfonamide Inhibitors of Carbonic Anhydrases from Mammaliicoccus sciuri Prevent Antimicrobial Resistance Due to Gene Transfer to Other Harmful Staphylococci? International Journal of Molecular Sciences. 2022; 23(22):13827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213827
Chicago/Turabian StyleDe Luca, Viviana, Simone Giovannuzzi, Claudiu T. Supuran, and Clemente Capasso. 2022. "May Sulfonamide Inhibitors of Carbonic Anhydrases from Mammaliicoccus sciuri Prevent Antimicrobial Resistance Due to Gene Transfer to Other Harmful Staphylococci?" International Journal of Molecular Sciences 23, no. 22: 13827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232213827