

Combating Black Fungus: Using Allicin as a Potent Antifungal Agent against Mucorales
 , , and
, , and
Abstract
:1. Introduction
2. Results
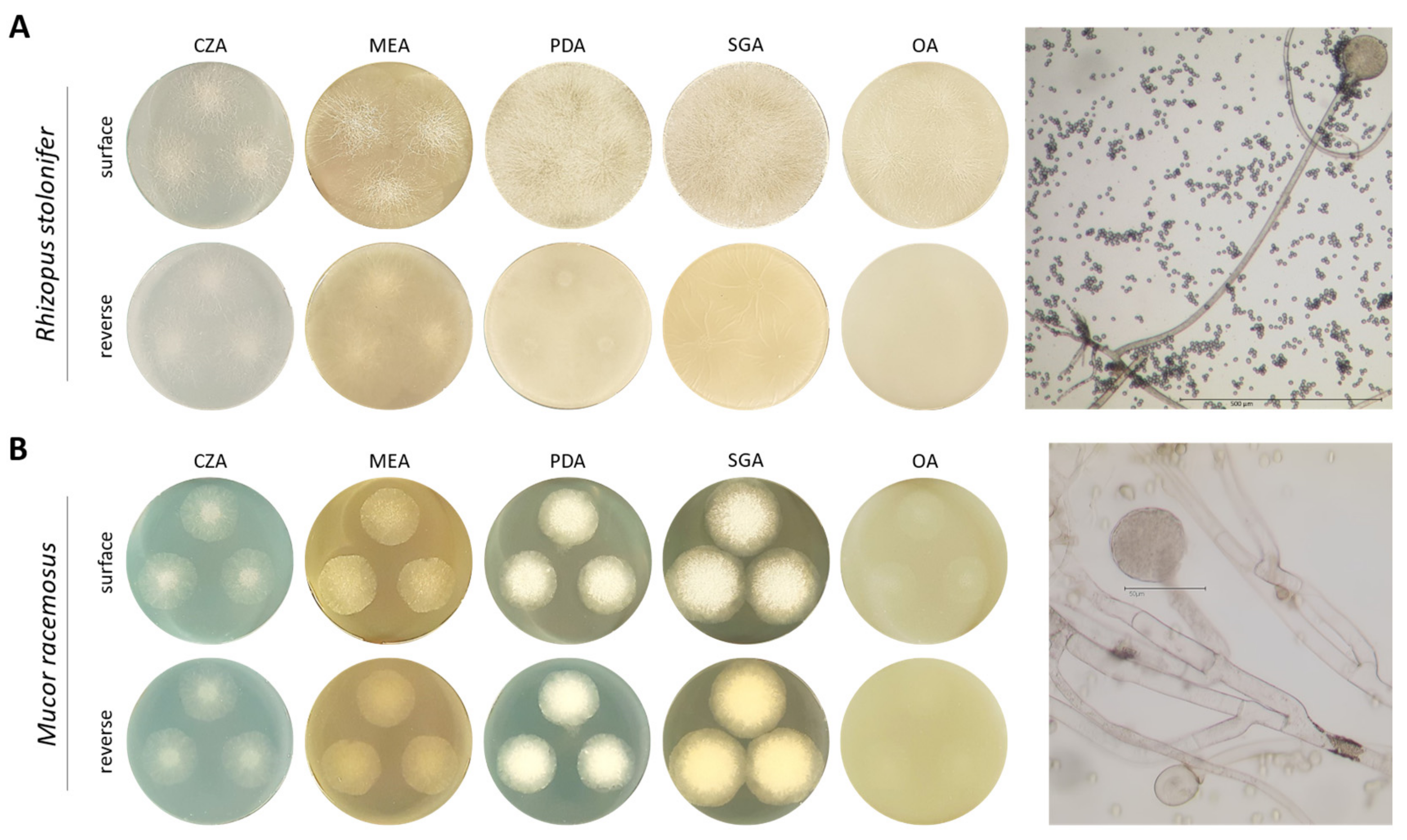
2.1. Morphological Characterisation of Mucorales Species
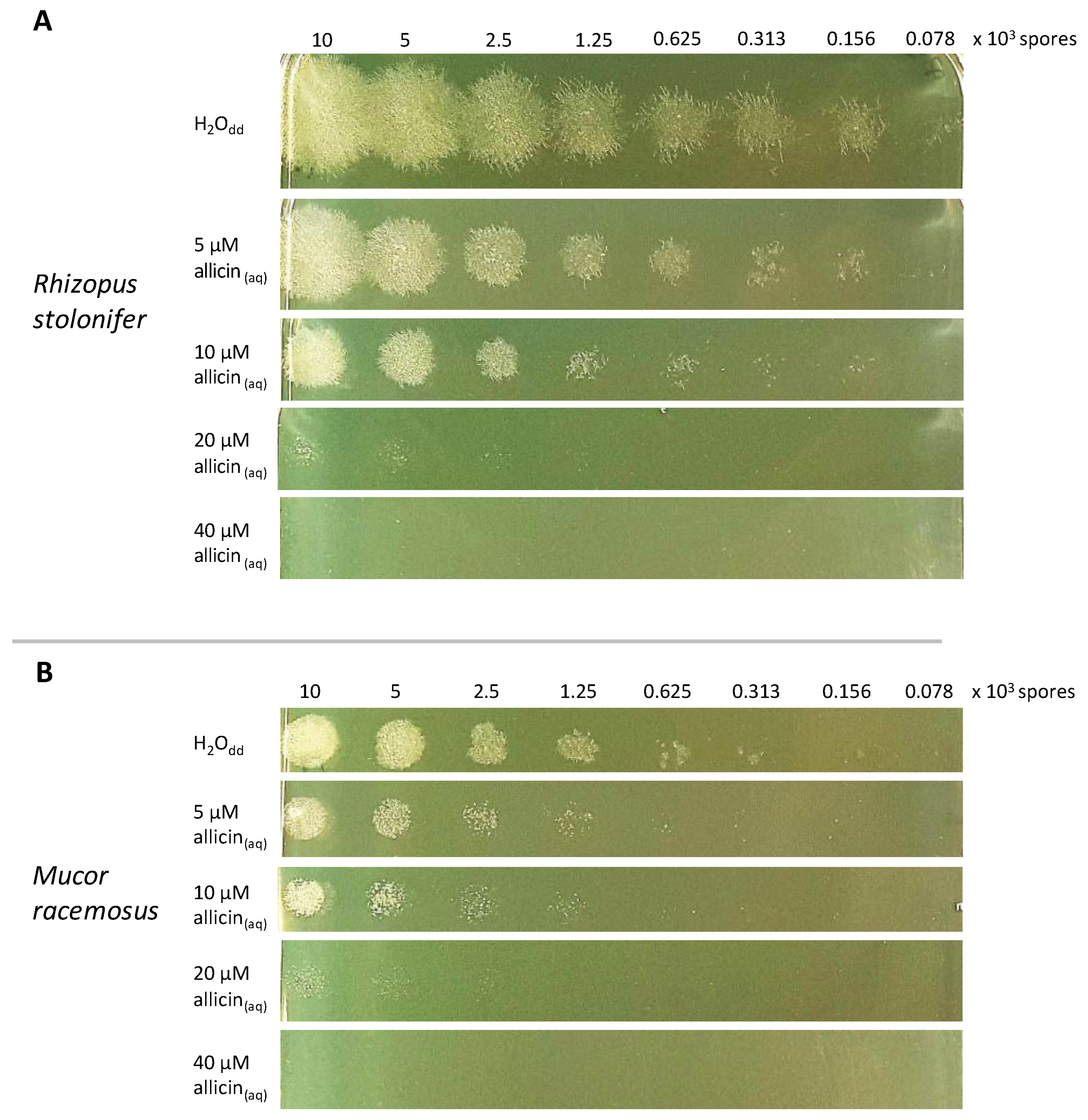
2.2. Allicin Susceptibility Depends on Spore Amount
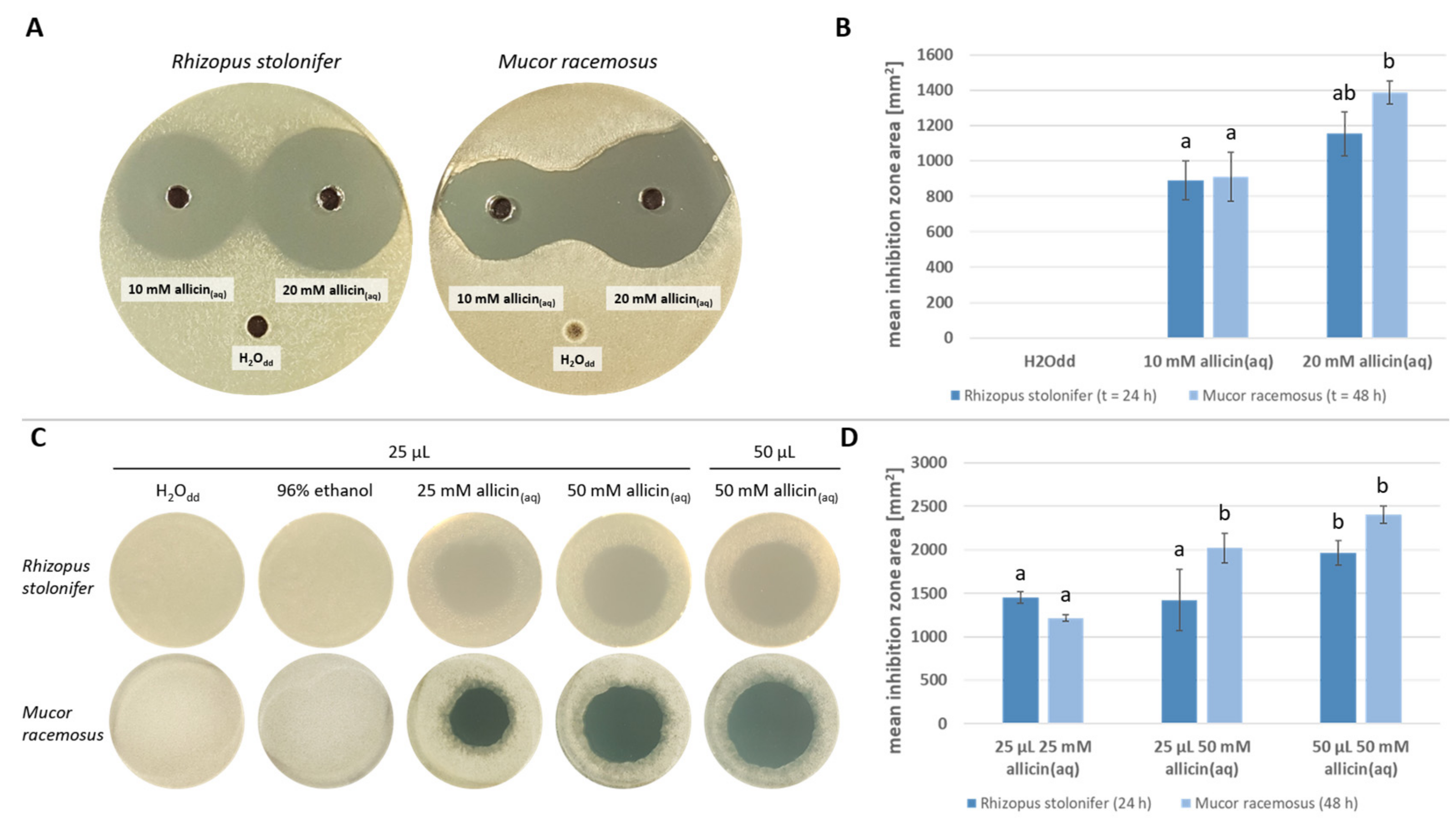
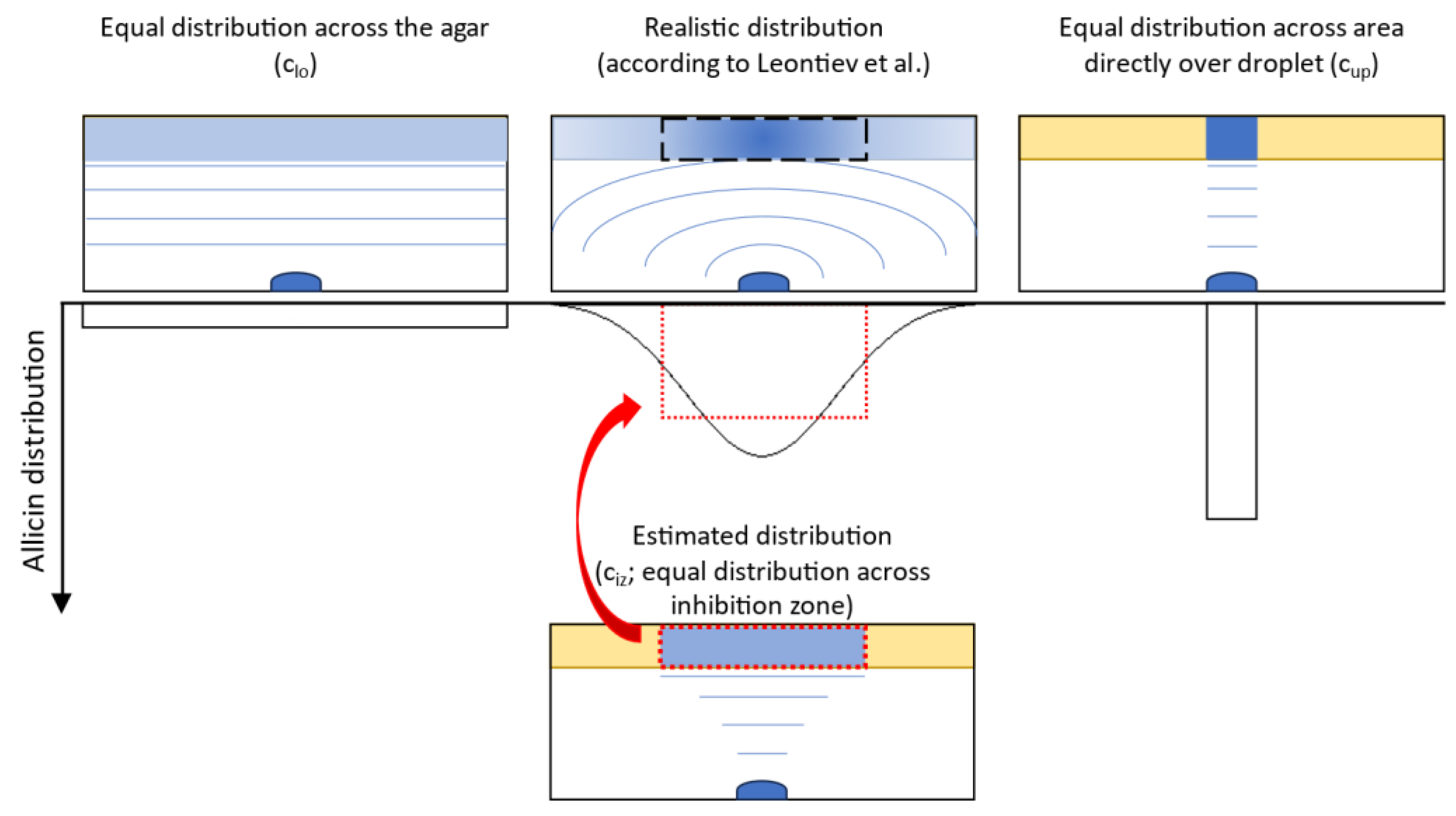
2.3. Allicin Inhibits Mucorales Spore Germination by Diffusion and as a Vapour
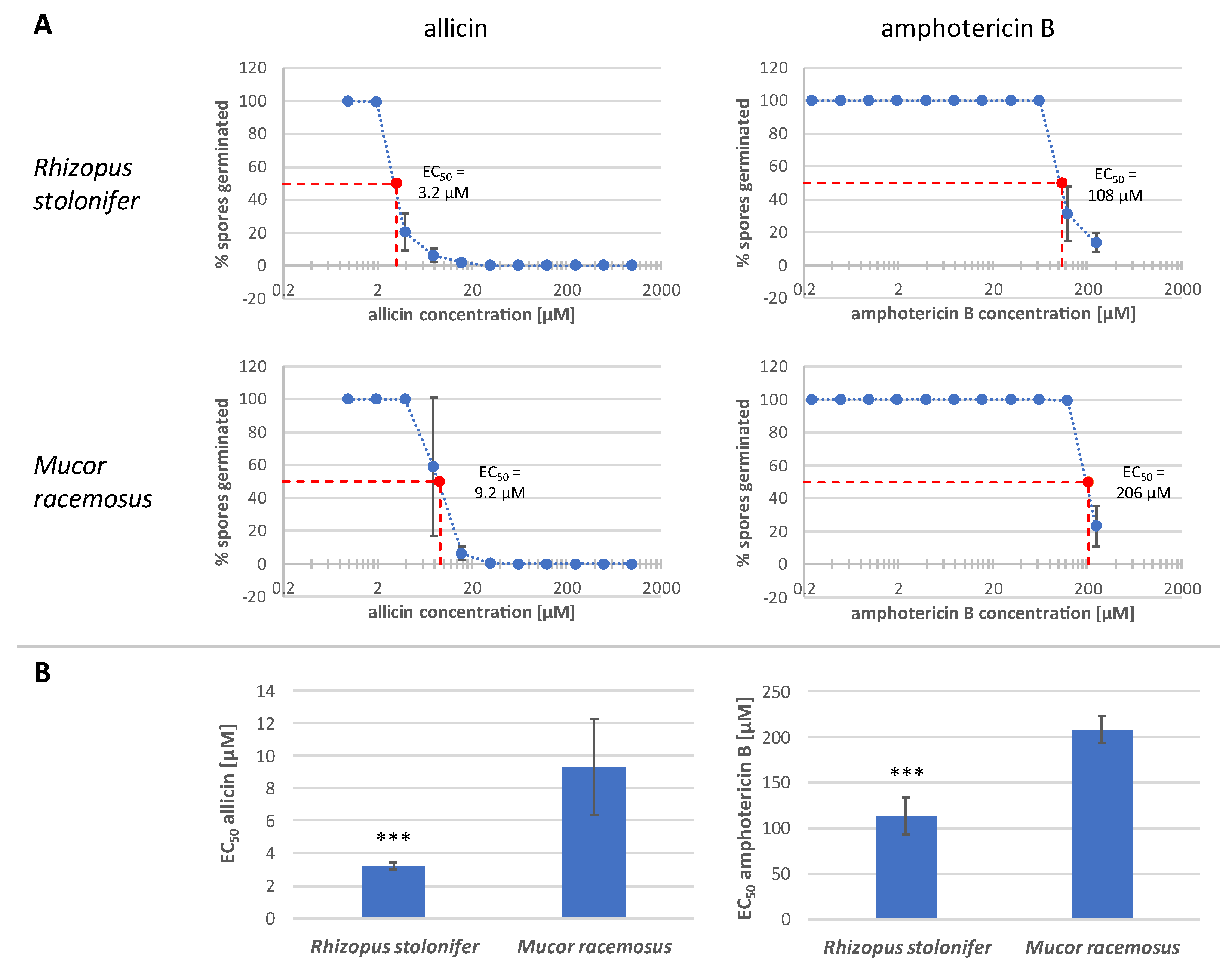
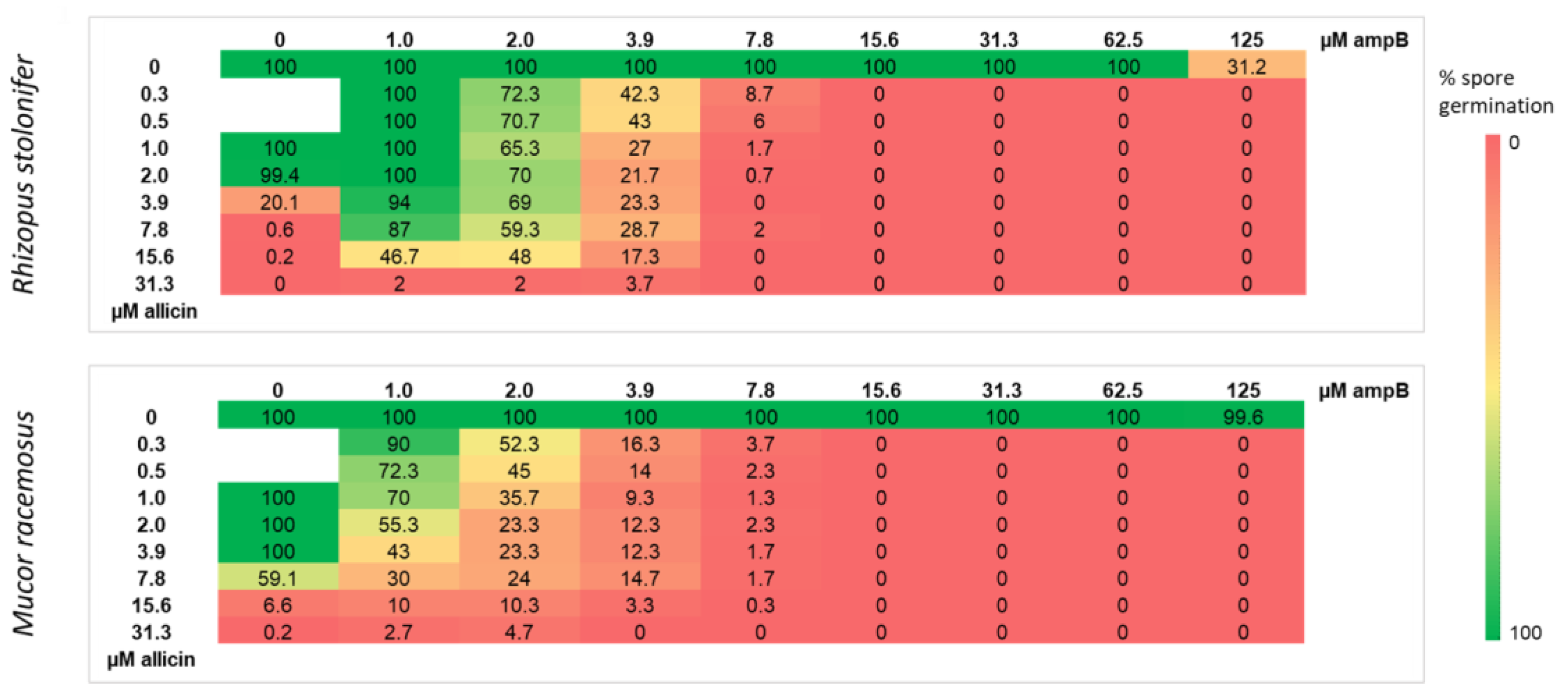
2.4. Allicin Inhibits Spore Germination More Effectively Than Amphotericin B upon Direct Contact
2.5. The Efficacy of Allicin as a Vapour in Inhibiting Spore Germination Is Comparable to That of a Direct Application of Amphotericin B
- R. stolonifer seems to be less susceptible to allicin than M. racemosus for vapor and diffusion treatment.
- Allicin application via diffusion is approx. 1.8 times more effective than vapor regardless of the species.
- Allicin vapor reaches effective concentrations of the same order of magnitude as ampBs’ direct application. Thus, inhibition of spore germination is more or equally effective with allicin vapor.
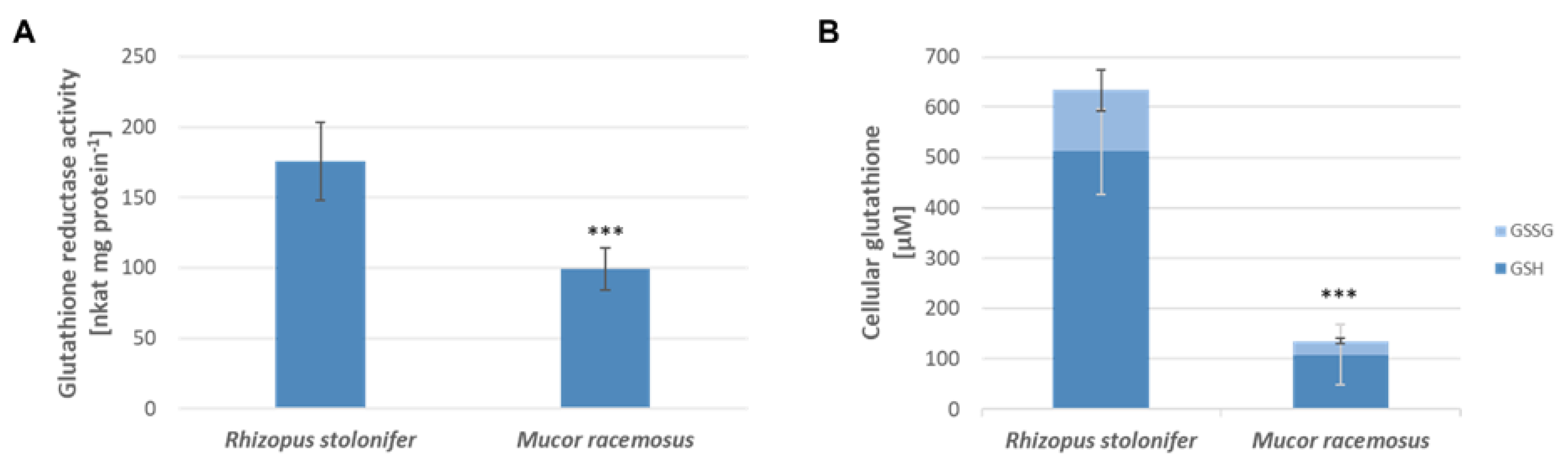
2.6. Correlation between Allicin Resistance, Glutathione Reductase Activity and Cellular Glutathione Levels
2.7. Allicin and Amphotericin B Show a Synergistic Effect against Mucorales
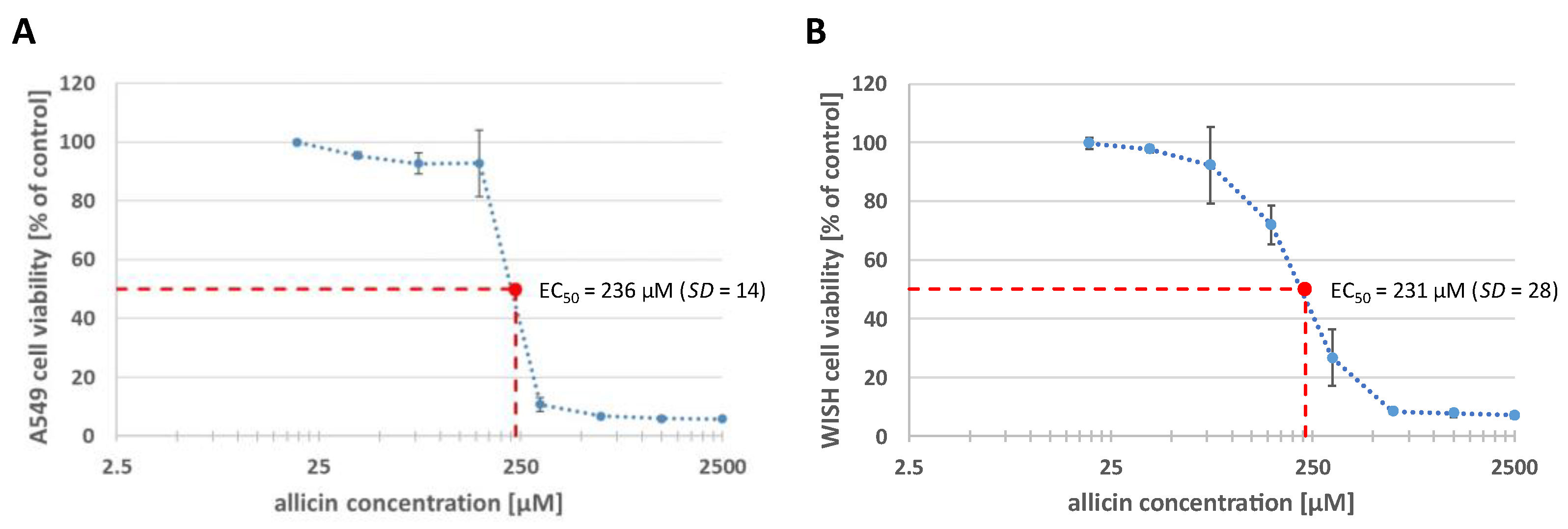
2.8. Allicin Has a Cytotoxic Effect on Human Epithelial Cell Lines
3. Discussion
4. Materials and Methods
4.1. Allicin Synthesis
4.2. Fungi Procurement and Cultivation Methods
4.3. Growth Assay and Macroscopic Characterization
- Potato dextrose agar (PDA): 39 g potato glucose agar (Carl Roth GmbH + Co. KG, Karlsruhe, Germany);
- Czapek agar (CZA): 30 g saccharose, 2.5 g NaNO3, 1 g K2HPO4, 0.5 g KCl, 0.5 g MgSO4 · 7H2O, 0.01 g FeSO4 · 7H2O;
- Malt extract agar (MEA): 15 g malt extract (Carl Roth GmbH + Co. KG, Karlsruhe, Germany);
- Sabouraud glucose agar (SGA): 40 g glucose monohydrate, 10 g peptone;
- Oatmeal agar (OA): 30 g of oat flakes were brought to a boil in 1 L deionised water and allowed to simmer gently for 2 h. The oat flakes were then suspended. The suspension was filtered and agar was added to the filtrate.
4.4. Inhibition Zone Tests
4.5. Drop Test
4.6. Determination of the Half Maximal Effective Concentration (EC50) and the Minimum Inhibitory Concentration (MIC)
4.7. Investigation of Synergistic Effect of Allicin and AmpB
4.8. Calibration of the Glutathione Reductase (GR) and Glutathione Determination Assay
4.9. Preparation of Cell Lysates
4.10. Determination of Protein Levels by Bradford Assay
4.11. Glutathione Reductase Enzyme Activity Assay
4.12. Determination of Cellular Glutathione Levels
4.13. Allicin Toxicity on Human Epithelial Cell Lines
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden Killers: Human Fungal Infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed]
- Hoenigl, M.; Seidel, D.; Sprute, R.; Cunha, C.; Oliverio, M.; Goldman, G.H.; Ibrahim, A.S.; Carvalho, A. COVID-19-Associated Fungal Infections. Nat. Microbiol. 2022, 7, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; WHO: Geneva, Switzerland, 2022; Volume 1, ISBN 9789240060241. [Google Scholar]
- Vermeulen, E.; Lagrou, K.; Verweij, P.E. Azole Resistance in Aspergillus Fumigatus: A Growing Public Health Concern. Curr. Opin. Infect. Dis. 2013, 26, 493–500. [Google Scholar] [CrossRef]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the Emerging Threat of Antifungal Resistance to Human Health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef]
- Denning, D.W.; Bromley, M.J. How to Bolster the Antifungal Pipeline. Science 2015, 347, 1414–1416. [Google Scholar] [CrossRef]
- Hoenigl, M.; Sprute, R.; Egger, M.; Arastehfar, A.; Cornely, O.A.; Krause, R.; Lass-Flörl, C.; Prattes, J.; Spec, A.; Thompson, G.R.; et al. The Antifungal Pipeline: Fosmanogepix, Ibrexafungerp, Olorofim, Opelconazole, and Rezafungin. Drugs 2021, 81, 1703–1729. [Google Scholar] [CrossRef] [PubMed]
- Roden, M.M.; Zaoutis, T.E.; Buchanan, W.L.; Knudsen, T.A.; Sarkisova, T.A.; Schaufele, R.L.; Sein, M.; Sein, T.; Chiou, C.C.; Chu, J.H.; et al. Epidemiology and Outcome of Zygomycosis: A Review of 929 Reported Cases. Clin. Infect. Dis. 2005, 41, 634–653. [Google Scholar] [CrossRef]
- Jeong, W.; Keighley, C.; Wolfe, R.; Lee, W.L.; Slavin, M.A.; Kong, D.C.M.; Chen, S.C.A. The Epidemiology and Clinical Manifestations of Mucormycosis: A Systematic Review and Meta-Analysis of Case Reports. Clin. Microbiol. Infect. 2019, 25, 26–34. [Google Scholar] [CrossRef]
- Raut, A.; Huy, N.T. Rising Incidence of Mucormycosis in Patients with COVID-19: Another Challenge for India amidst the Second Wave? Lancet Respir. Med. 2021, 9, e77. [Google Scholar] [CrossRef]
- Hoenigl, M.; Seidel, D.; Carvalho, A.; Rudramurthy, S.M.; Arastehfar, A.; Gangneux, J.-P.; Nasir, N.; Bonifaz, A.; Araiza, J.; Klimko, N.; et al. The Emergence of COVID-19 Associated Mucormycosis: A Review of Cases from 18 Countries. Lancet Microbe 2022, 5247, E543–E552. [Google Scholar] [CrossRef]
- Tayabali, K.; Pothiwalla, H.; Narayanan, S. Epidemiology of COVID-19–Associated Mucormycosis. Curr. Fungal Infect. Rep. 2023, 17, 156–175. [Google Scholar] [CrossRef] [PubMed]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E. Global Guideline for the Diagnosis and Management of Mucormycosis: An Initiative of the European Confederation of Medical Mycology in Cooperation with the Mycoses Study Group Education and Research Consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef] [PubMed]
- Muthu, V.; Agarwal, R.; Dhooria, S.; Sehgal, I.S.; Prasad, K.T.; Aggarwal, A.N.; Chakrabarti, A. Has the Mortality from Pulmonary Mucormycosis Changed over Time? A Systematic Review and Meta-Analysis. Clin. Microbiol. Infect. 2021, 27, 538–549. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Edwards, J.E.; Bryant, R.; Spellberg, B. Economic Burden of Mucormycosis in the United States: Can a Vaccine Be Cost-Effective? Med. Mycol. 2009, 47, 592–600. [Google Scholar] [CrossRef]
- Madanagopal, P.; Ramprabhu, N.; Jagadeesan, R. In Silico Prediction and Structure-Based Multitargeted Molecular Docking Analysis of Selected Bioactive Compounds against Mucormycosis. Bull. Natl. Res. Cent. 2022, 46, 1–21. [Google Scholar] [CrossRef]
- Sharma, A.D.; Kaur, I.; Chauhan, A. Compositional Profiling and Molecular Docking Studies of Eucalyptus polybrachtea Essential Oil against Mucormycosis and Aspergillosis. BioTechnologia 2023, 104, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Nimoshini, G.; Nataraj, P.; Rameshkumar, A.; Krishnan, R.; Savithri, S. Inhibitory Effect of Garlic and Omam Extracts on Mucor circinelloides, a Fungus Causing Mucormycosis: An in Vitro Study. J. Oral Maxillofac. Pathol. 2023, 27, 71–75. [Google Scholar] [CrossRef]
- Cavallito, C.J.; Bailey, J.H. Allicin, the Antibacterial Principle of Allium sativum. I. Isolation, Physical Properties and Antibacterial Action. J. Am. Chem. Soc. 1944, 66, 1950–1951. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Foerster, J.; Kappler, U.; Antelmann, H.; Noll, U.; Gruhlke, M.C.H.; Slusarenko, A.J. Allicin, the Odor of Freshly Crushed Garlic: A Review of Recent Progress in Understanding Allicin’s Effects on Cells. Molecules 2021, 26, 1505. [Google Scholar] [CrossRef]
- Baysal, G.; Olcay, H.S.; Günneç, Ç. Encapsulation and Antibacterial Studies of Goji Berry and Garlic Extract in the Biodegradable Chitosan. J. Bioact. Compat. Polym. 2023, 38, 209–219. [Google Scholar] [CrossRef]
- Schier, C.; Foerster, J.; Heupel, M.; Dörner, P.; Klaas, M.; Schröder, W.; Rink, L.; Slusarenko, A.J.; Gruhlke, M.C.H. Allicin as a Volatile or Nebulisable Antimycotic for the Treatment of Pulmonary Mycoses: In Vitro Studies Using a Lung Flow Test Rig. Int. J. Mol. Sci. 2022, 23, 6607. [Google Scholar] [CrossRef] [PubMed]
- Shadkchan, Y.; Shemesh, E.; Mirelman, D.; Miron, T.; Rabinkov, A.; Wilchek, M.; Osherov, N. Efficacy of Allicin, the Reactive Molecule of Garlic, in Inhibiting Aspergillus Spp. in Vitro, and in a Murine Model of Disseminated Aspergillosis. J. Antimicrob. Chemother. 2004, 53, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Coppi, A.; Cabinian, M.; Mirelman, D.; Sinnis, P. Antimalarial Activity of Allicin, a Biologically Active Compound from Garlic Cloves. Antimicrob. Agents Chemother. 2006, 50, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Ankri, S.; Miron, T.; Rabinkov, A.; Wilchek, M.; Mirelman, D. Allicin from Garlic Strongly Inhibits Cysteine Proteinases and Cytopathic Effects of Entamoeba Histolytica. Antimicrob. Agents Chemother. 1997, 41, 2286–2288. [Google Scholar] [CrossRef] [PubMed]
- Weber, N.D.; Andersen, D.O.; North, J.A.; Murray, B.K.; Lawson, L.D.; Hughes, B.G. In Vitro Virucidal Effects of Allium sativum (Garlic) Extract and Compounds. Planta Med. 1992, 58, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.; Cole, L.L.; Davis, L.E.; Lockwood, S.J.; Simmons, V.; Wild, G.C. Antiviral Properties of Garlic: In Vitro Effects on Influenza B, Herpes Simplex and Coxsackie Viruses. Planta Med. 1985, 51, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Gruhlke, M.C.H.; Antelmann, H.; Bernhardt, J.; Kloubert, V.; Rink, L.; Slusarenko, A.J. The Human Allicin-Proteome: S-Thioallylation of Proteins by the Garlic Defence Substance Allicin and Its Biological Effects. Free Radic. Biol. Med. 2019, 131, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Van Loi, V.; Huyen, N.T.T.; Busche, T.; Tung, Q.N.; Gruhlke, M.C.H.; Kalinowski, J.; Bernhardt, J.; Slusarenko, A.J.; Antelmann, H. Staphylococcus aureus Responds to Allicin by Global S-Thioallylation—Role of the Brx/BSH/YpdA Pathway and the Disulfide Reductase MerA to Overcome Allicin Stress. Free Radic. Biol. Med. 2019, 139, 55–69. [Google Scholar] [CrossRef]
- Willis, E.D. Enzyme Inhibition by Allicin, the Active Principle of Garlic. Biochem. J. 1956, 63, 514–520. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, D.K. Enzyme Inhibition by Allicin, the Molluscicidal Agent of Allium sativum L. (Garlic). Phyther. Res. 1996, 10, 383–386. [Google Scholar] [CrossRef]
- Prager-Khoutorsky, M.; Goncharov, I.; Rabinkov, A.; Mirelman, D.; Geiger, B.; Bershadsky, A.D. Allicin Inhibits Cell Polarization, Migration and Division via Its Direct Effect on Microtubules. Cell Motil. Cytoskelet. 2007, 64, 321–337. [Google Scholar] [CrossRef]
- Mösbauer, K.; Fritsch, V.N.; Adrian, L.; Bernhardt, J.; Gruhlke, M.C.H.; Slusarenko, A.J.; Niemeyer, D.; Antelmann, H. The Effect of Allicin on the Proteome of SARS-CoV-2 Infected Calu-3 Cells. Front. Microbiol. 2021, 12, 746795. [Google Scholar] [CrossRef]
- Jobani, B.M.; Najafzadeh, N.; Mazani, M.; Arzanlou, M.; Vardin, M.M. Molecular Mechanism and Cytotoxicity of Allicin and All-Trans Retinoic Acid against CD44+ versus CD117+ Melanoma Cells. Phytomedicine 2018, 48, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Li, Y.; Chi, Y. Role of P38 MAPK Activation and Mitochondrial Cytochrome-c Release in Allicin-Induced Apoptosis in SK-N-SH Cells. Anticancer Drugs 2016, 27, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Miron, T.; Wilchek, M.; Sharp, A.; Nakagawa, Y.; Naoi, M.; Nozawa, Y.; Akao, Y. Allicin Inhibits Cell Growth and Induces Apoptosis through the Mitochondrial Pathway in HL60 and U937 Cells. J. Nutr. Biochem. 2008, 19, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Leontiev, R.; Hohaus, N.; Jacob, C.; Gruhlke, M.C.H.; Slusarenko, A.J. A Comparison of the Antibacterial and Antifungal Activities of Thiosulfinate Analogues of Allicin. Sci. Rep. 2018, 8, 6763. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, K.S.; Han, I.; Kim, M.H.; Jung, M.H.; Park, H.K. Quantitative and Qualitative Analysis of the Antifungal Activity of Allicin Alone and in Combination with Antifungal Drugs. PLoS ONE 2012, 7, e38242. [Google Scholar] [CrossRef]
- Corral, M.J.; González-Sánchez, E.; Cuquerella, M.; Alunda, J.M. In Vitro Synergistic Effect of Amphotericin B and Allicin on Leishmania Donovani and L. Infantum. Antimicrob. Agents Chemother. 2014, 58, 1596–1602. [Google Scholar] [CrossRef]
- Li, Z.; Li, Z.; Yang, J.; Lu, C.; Li, Y.; Luo, Y.; Cong, F.; Shi, R.; Wang, Z.; Chen, H.; et al. Allicin Shows Antifungal Efficacy against Cryptococcus Neoformans by Blocking the Fungal Cell Membrane. Front. Microbiol. 2022, 13, 1012516. [Google Scholar] [CrossRef]
- Yamada, Y.; Azuma, K. Evaluation of the in Vitro Antifungal Activity of Allicin. Antimicrob. Agents Chemother. 1977, 11, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.D.; Wood, S.G.; Hughes, B.G. HPLC Analysis of Allicin and Other Thiosulfinates in Garlic Clove Homogenates. Planta Med. 1991, 57, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, H.; Suma, K.; Origuchi, K.; Kumagai, H.; Seki, T.; Ariga, T. Biological and Chemical Stability of Garlic-Derived Allicin. J. Agric. Food Chem. 2008, 56, 4229–4235. [Google Scholar] [CrossRef]
- Mengers, H.G.; Schier, C.; Zimmermann, M.; Gruhlke, M.C.H.; Block, E.; Blank, L.M.; Slusarenko, A.J. Seeing the Smell of Garlic: Detection of Gas Phase Volatiles from Crushed Garlic (Allium sativum), Onion (Allium cepa), Ramsons (Allium ursinum) and Human Garlic Breath Using SESI-Orbitrap MS. Food Chem. 2022, 397, 133804. [Google Scholar] [CrossRef] [PubMed]
- Safar, A.; Marsan, J.; Marglani, O.; Al-Sebeih, K.; Al-Harbi, J.; Valvoda, M. Early Identification of Rhinocerebral Mucormycosis. J. Otolaryngol. 2005, 34, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, F.; Leontiev, R.; Jacob, C.; Slusarenko, A.J. An Optimized Facile Procedure to Synthesize and Purify Allicin. Molecules 2017, 22, 770. [Google Scholar] [CrossRef]
- Koch, H.P.; Lawson, L.D. Garlic: The Science and Therapeutic Application of Allium sativum L. and Related Species; Williams & Wilkins: Baltimore, MD, USA, 1996. [Google Scholar]
- Block, E. Garlic and the Other Alliums. The Lore and the Science, 1st ed.; RSC Publishing: Cambridge, UK, 2010. [Google Scholar]
- Minchin, W.D. A Study in Tubercule Virus Polymorphism, and the Treatment of Tuberculosis and Lupus with Oleum Allii, 3rd ed.; Baillière, Tindall and Cox: London, UK, 1927. [Google Scholar]
- Joshi, S.; Telang, R.; Tambe, M.; Havaldar, R.; Sane, M.; Shaikh, A.; Roy, C.; Yathati, K.; Sonawale, S.; Borkar, R.; et al. Outbreak of Mucormycosis in Coronavirus Disease Patients, Pune, India. Emerg. Infect. Dis. 2022, 28, 1. [Google Scholar] [CrossRef]
- Wingard, J.R.; Kubilis, P.; Lee, L.; Yee, G.; White, M.; Walshe, L.; Bowden, R.; Anaissie, E.; Hiemenz, J.; Lister, J. Clinical Significance of Nephrotoxicity in Patients Treated with Amphotericin B for Suspected or Proven Aspergillosis. Clin. Infect. Dis. 1999, 29, 1402–1407. [Google Scholar] [CrossRef]
- Deray, G.; Mercadal, L.; Bagnis, C. Amphotericin B Nephrotoxicity. Nephrologie 2002, 23, 119–122. [Google Scholar] [CrossRef]
- de Kruijff, B.; Demel, R.A. Polyene Antibiotic-Sterol Interactions in Membranes of Acholeplasma laidlawii Cells and Lecithin Liposomes. 3. Molecular Structure of the Polyene Antibiotic-Cholesterol Complexes. Biochim. Biophys. Acta 1974, 339, 57–70. [Google Scholar] [CrossRef]
- Ermishkin, L.N.; Kasumov, K.M.; Potzeluyev, V.M. Single Ionic Channels Induced in Lipid Bilayers by Polyene Antibiotics Amphotericin B and Nystatine. Nature 1976, 262, 698–699. [Google Scholar] [CrossRef]
- Anderson, T.M.; Clay, M.C.; Cioffi, A.G.; Diaz, K.A.; Hisao, G.S.; Tuttle, M.D.; Nieuwkoop, A.J.; Comellas, G.; Maryum, N.; Wang, S.; et al. Amphotericin Forms an Extramembranous and Fungicidal Sterol Sponge. Nat. Chem. Biol. 2014, 10, 400–406. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Portz, D.; Stitz, M.; Anwar, A.; Schneider, T.; Jacob, C.; Schlaich, N.L.; Slusarenko, A.J. Allicin Disrupts the Cell’s Electrochemical Potential and Induces Apoptosis in Yeast. Free Radic. Biol. Med. 2010, 49, 1916–1924. [Google Scholar] [CrossRef] [PubMed]
- Getti, G.T.M.; Poole, P.L. Allicin Causes Fragmentation of the Peptidoglycan Coat in Staphylococcus aureus by Effecting Synthesis and Aiding Hydrolysis: A Determination by MALDI-TOF Mass Spectrometry on Whole Cells. J. Med. Microbiol. 2019, 68, 667–677. [Google Scholar] [CrossRef]
- Yang, X.; Bai, S.; Wu, J.; Fan, Y.; Zou, Y.; Xia, Z.; Ao, J.; Chen, T.; Zhang, M.; Yang, R. Antifungal Activity and Potential Action Mechanism of Allicin against Trichosporon asahii. Microbiol. Spectr. 2023, 11, e00907-23. [Google Scholar] [CrossRef] [PubMed]
- Adetumbi, M.; Javor, G.T.; Lau, B.H.S. Allium sativum (Garlic) Inhibits Lipid Synthesis by Candida Albicans. Antimicrob. Agents Chemother. 1986, 30, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A. Studies on the Anticandidal Mode of Action of Allium sativum (Garlic). J. Gen. Microbiol. 1988, 134, 2917–2924. [Google Scholar] [CrossRef]
- Yu, L.; Guo, N.; Meng, R.; Liu, B.; Tang, X.; Jin, J.; Cui, Y.; Deng, X. Allicin-Induced Global Gene Expression Profile of Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2010, 88, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Porter, T.D. Garlic and Garlic-Derived Compounds Inhibit Human Squalene Monooxygenase. J. Nutr. 2001, 131, 1662–1667. [Google Scholar] [CrossRef] [PubMed]
- Horn, T.; Bettray, W.; Slusarenko, A.; Gruhlke, M. S-Allylmercaptoglutathione Is a Substrate for Glutathione Reductase (E.C. 1.8.1.7) from Yeast (Saccharomyces cerevisiae). Antioxidants 2018, 7, 86. [Google Scholar] [CrossRef]
- Rahman, I. Inflammation and the Regulation of Glutathione Level in Lung Epithelial Cells. Antioxid. Redox Signal. 1999, 1, 425–447. [Google Scholar] [CrossRef]
- Cantin, A.M.; North, S.L.; Hubbard, R.C.; Crystal, R.G. Normal Alveolar Epithelial Lining Fluid Contains High Levels of Glutathione. J. Appl. Physiol. 1987, 63, 152–157. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of Glutathione and Glutathione Disulfide Using Glutathione Reductase and 2-Vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Determination of Glutathione and Glutathione Disulfide in Biological Samples. Methods Enzym. 1985, 113, 548–555. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mucorales Species | Mean Effective Concentration of Application Types to Inhibit Spore Germination [µM] | |||
|---|---|---|---|---|
| Allicin | Amphotericin B | |||
| Vapour | Diffusion | Direct Contact | Direct Contact | |
| Rhizopus stolonifer | 255 | 142 | 31.3 | >250 |
| Mucor racemosus | 210 | 118 | 31.3 | >250 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schier, C.; Gruhlke, M.C.H.; Reucher, G.; Slusarenko, A.J.; Rink, L. Combating Black Fungus: Using Allicin as a Potent Antifungal Agent against Mucorales. Int. J. Mol. Sci. 2023, 24, 17519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242417519
Schier C, Gruhlke MCH, Reucher G, Slusarenko AJ, Rink L. Combating Black Fungus: Using Allicin as a Potent Antifungal Agent against Mucorales. International Journal of Molecular Sciences. 2023; 24(24):17519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242417519
Chicago/Turabian StyleSchier, Christina, Martin C. H. Gruhlke, Georg Reucher, Alan J. Slusarenko, and Lothar Rink. 2023. "Combating Black Fungus: Using Allicin as a Potent Antifungal Agent against Mucorales" International Journal of Molecular Sciences 24, no. 24: 17519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242417519