
Novel Antibody–Peptide Binding Assay Indicates Presence of Immunoglobulins against EGFR Phospho-Site S1166 in High-Grade Glioma

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Detection of Anti-DNP-Bound Peptides with Melon Gel Resin
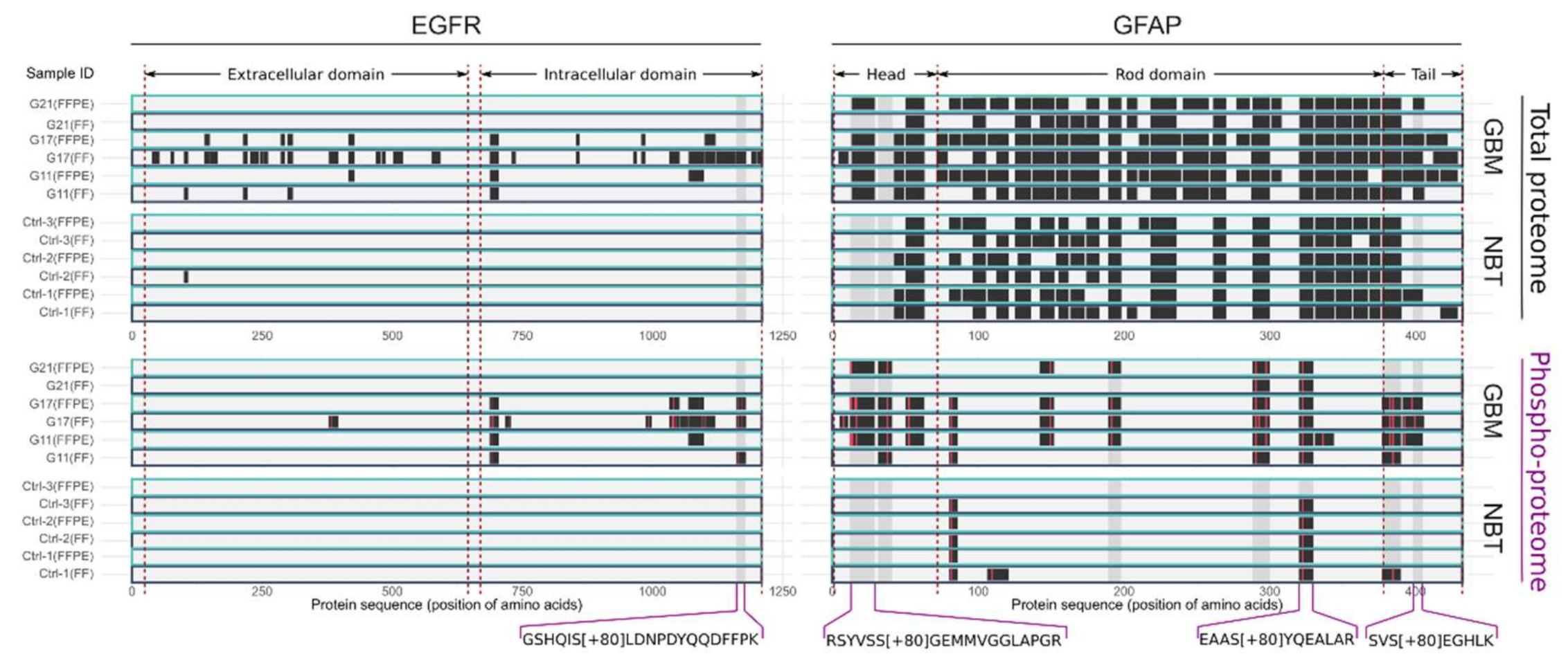
2.2. Selection of GBM-Associated Phospho-Peptide Antigens
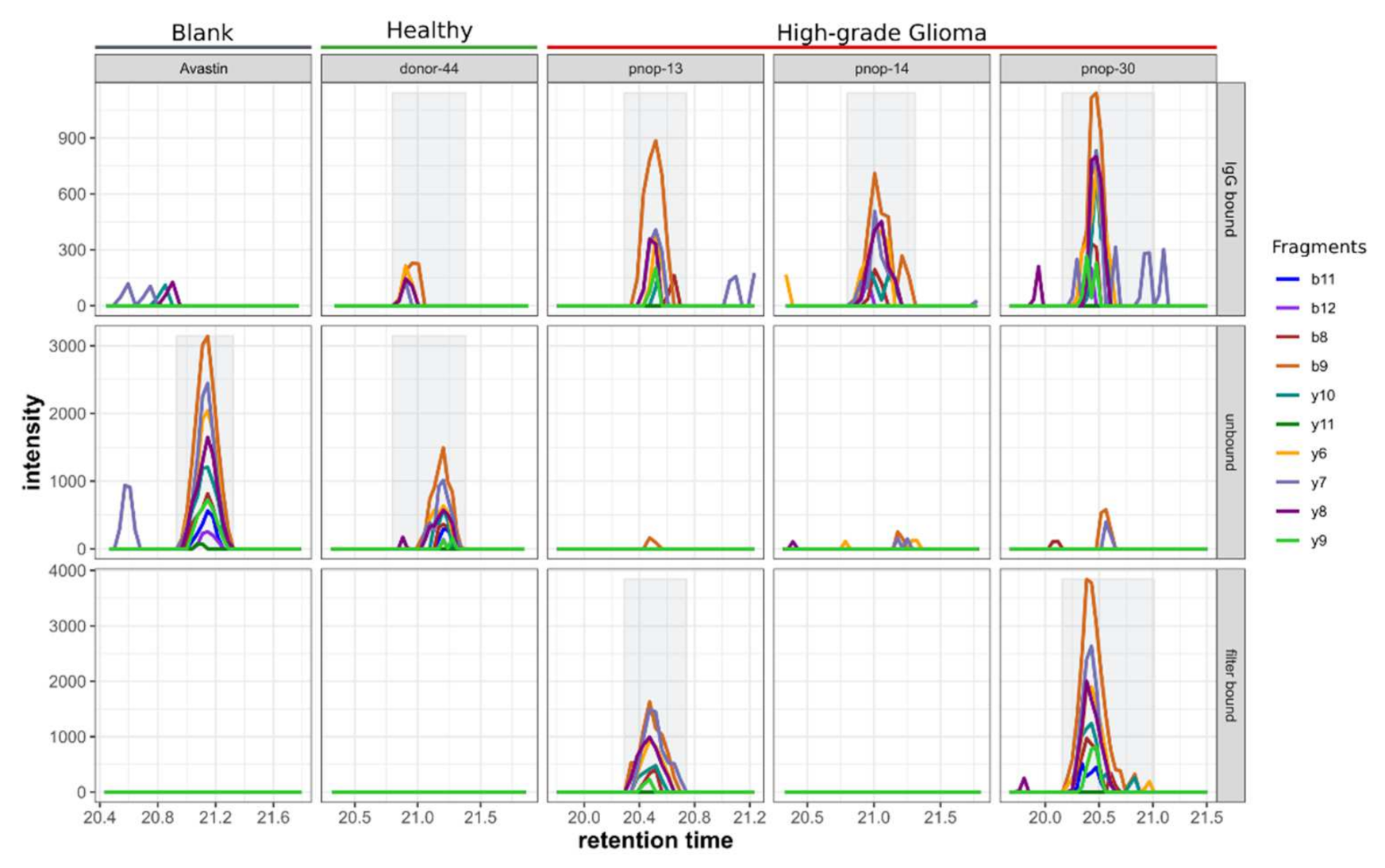
2.3. Ab–Peptide Binding Assay on EGFR and GFAP Phospho-Peptides
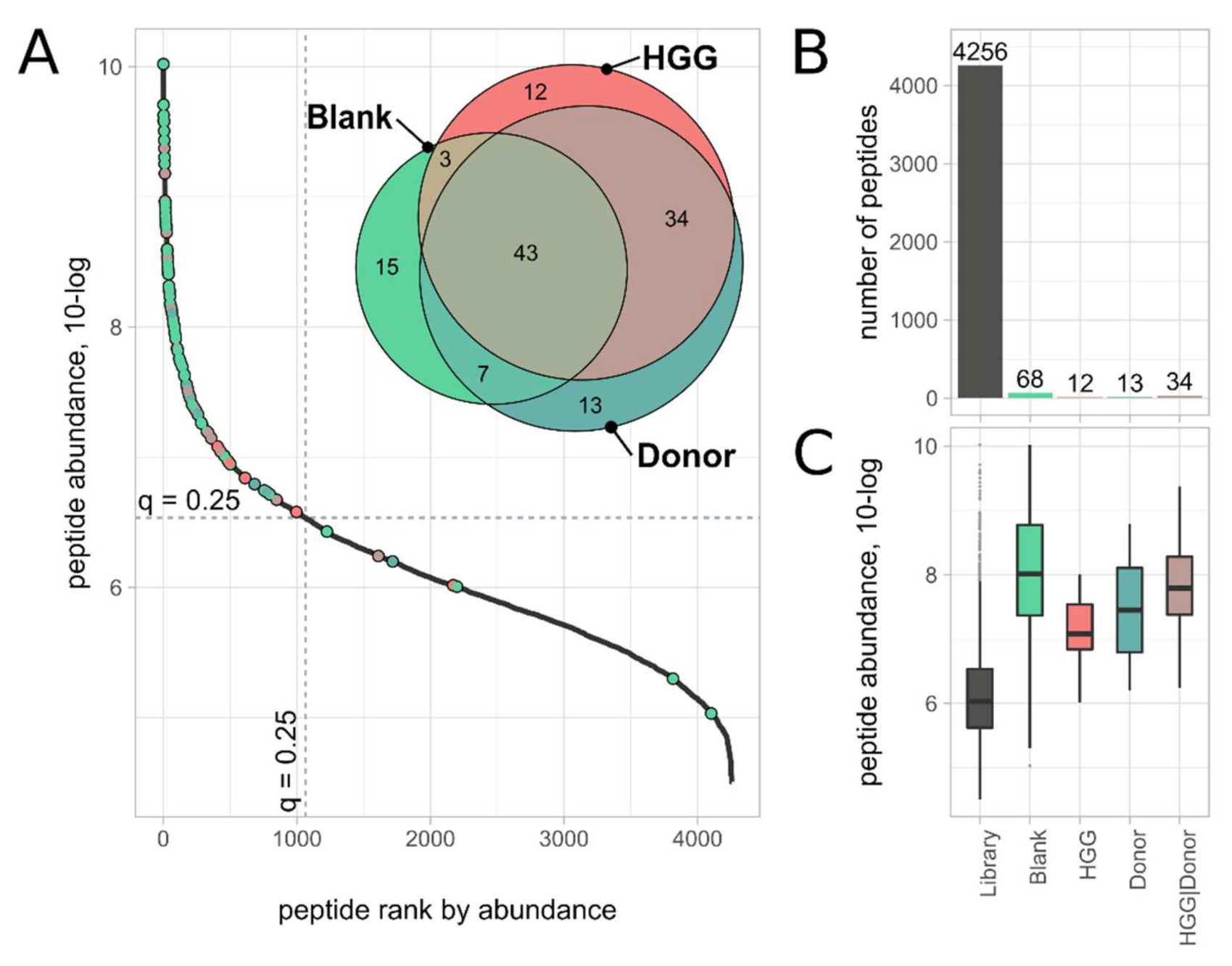
2.4. Mapping of Background and Unspecific Binding Peptides
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Preparation of Tissue Peptide Libraries
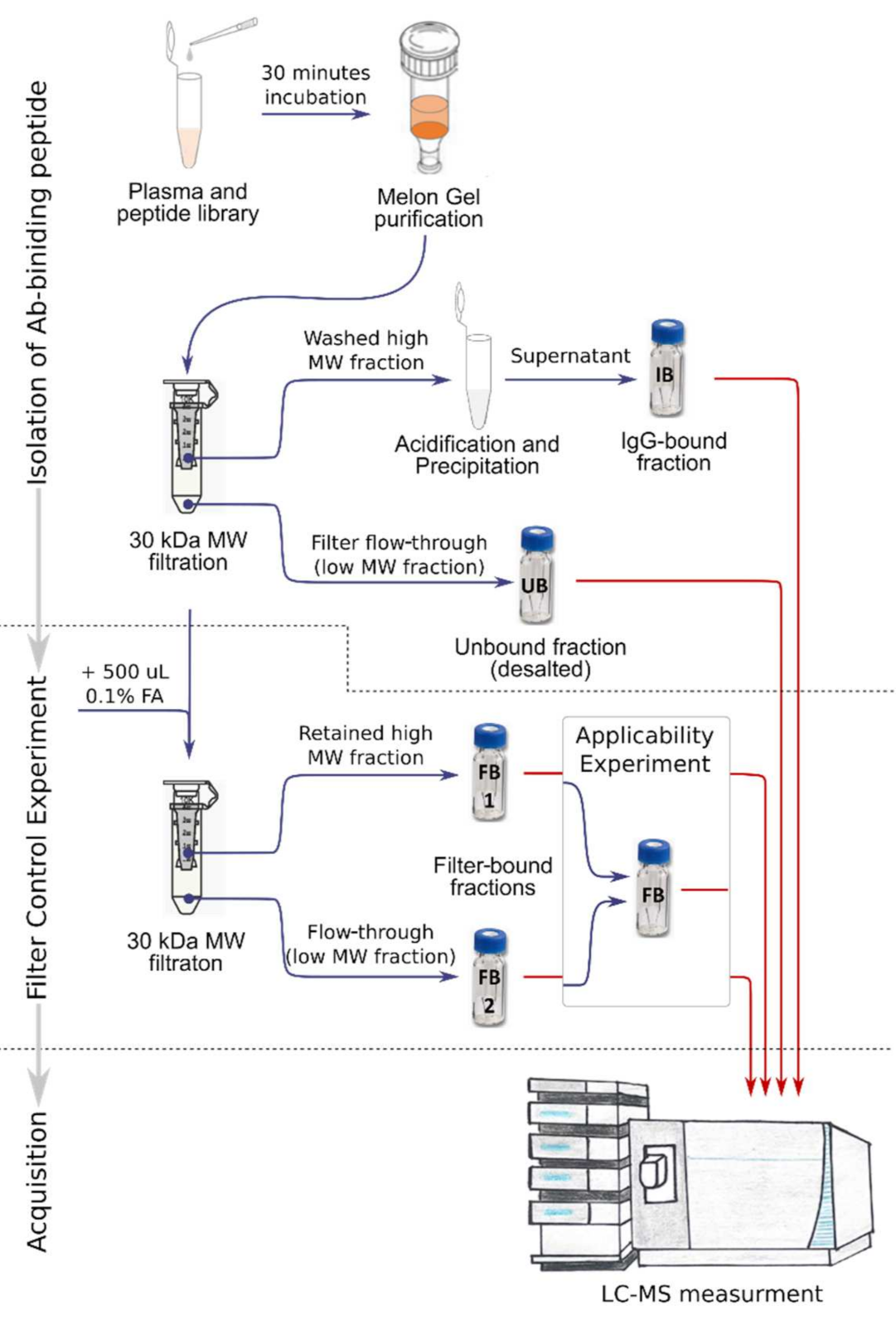
4.3. Isolation of Immunoglobulin-Binding Peptides
4.4. Targeted Parallel Reaction Monitoring (PRM) Measurements
4.5. Data-Dependent Mass Spectrometry Measurements
4.6. Feasibility Experiments
4.7. Selection of GBM-Associated Phospho-Peptide Antigens
4.8. Detection of Peptide–Antigen-Binding IgG in Plasma
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, H.T.; Low, J.; Lim, S.G.; Chung, M.C.M. Serum autoantibodies as biomarkers for early cancer detection. FEBS J. 2009, 276, 6880–6904. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Werner, S.; Tao, S.; Zörnig, I.; Brenner, H. Blood autoantibodies against tumor-associated antigens as biomarkers in early detection of colorectal cancer. Cancer Lett. 2014, 346, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Fan, Y.; Wu, J.; Wan, H.; Wang, J.; Lam, S.; Lam, W.L.; Girard, L.; Gazdar, A.F.; Wu, Z.; et al. Potential application of non-small cell lung cancer-associated autoantibodies to early cancer diagnosis. Biochem. Biophys. Res. Commun. 2012, 423, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.-Q.; Weng, X.-F.; Huang, X.-C.; Chu, L.-Y.; Wei, L.-F.; Lin, Y.-W.; Chen, L.-Y.; Liu, C.-T.; Xu, Y.-W.; Peng, Y.-H. A Panel of Tumor-associated Autoantibodies for the Detection of Early-stage Breast Cancer. J. Cancer 2021, 12, 2747–2755. [Google Scholar] [CrossRef] [PubMed]
- Rauf, F.; Anderson, K.S.; LaBaer, J. Autoantibodies in Early Detection of Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2020, 29, 2475–2485. [Google Scholar] [CrossRef] [PubMed]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Lo Muzio, L. The crucial role of protein phosphorylation in cell signaling and its use as targeted therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Mahoney, K.E.; Shabanowitz, J.; Hunt, D.F. MHC Phosphopeptides: Promising Targets for Immunotherapy of Cancer and Other Chronic Diseases. Mol. Cell. Proteom. 2021, 20, 100112. [Google Scholar] [CrossRef]
- Zeneyedpour, L.; Sten-van, T.H.J.; Luider, T. Using phosphoproteomics and next generation sequencing to discover novel therapeutic targets in patient antibodies. Expert Rev. Proteom. 2020, 17, 675–684. [Google Scholar] [CrossRef]
- Depontieu, F.R.; Qian, J.; Zarling, A.L.; McMiller, T.L.; Salay, T.M.; Norris, A.; English, A.M.; Shabanowitz, J.; Engelhard, V.H.; Hunt, D.F.; et al. Identification of tumor-associated, MHC class II-restricted phosphopeptides as targets for immunotherapy. Proc. Natl. Acad. Sci. USA 2009, 106, 12073–12078. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, F.; Stones, D.H.; Zarling, A.L.; Willcox, C.R.; Shabanowitz, J.; Cummings, K.L.; Hunt, D.F.; Cobbold, M.; Engelhard, V.H.; Willcox, B.E. The antigenic identity of human class I MHC phosphopeptides is critically dependent upon phosphorylation status. Oncotarget 2017, 8, 54160–54172. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.C.; Russell, J.D.; Bailey, D.J.; Westphall, M.S.; Coon, J.J. Parallel Reaction Monitoring for High Resolution and High Mass Accuracy Quantitative, Targeted Proteomics. Mol. Cell. Proteom. 2012, 11, 1475–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osinalde, N.; Aloria, K.; Omaetxebarria, M.J.; Kratchmarova, I. Targeted mass spectrometry: An emerging powerful approach to unblock the bottleneck in phosphoproteomics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1055–1056, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.T.; Searle, B.C.; Llovet, A.; Villén, J. Plug-and-play analysis of the human phosphoproteome by targeted high-resolution mass spectrometry. Nat. Methods 2016, 13, 431–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.L.; Dionne, U.; Lambert, J.-P.; Bisson, N. Targeted proteomics analyses of phosphorylation-dependent signalling networks. J. Proteom. 2018, 189, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Zeneyedpour, L.; Stingl, C.; Dekker, L.J.M.; Mustafa, D.A.M.; Kros, J.M.; Luider, T.M. Phosphorylation Ratio Determination in Fresh-Frozen and Formalin-Fixed Paraffin-Embedded Tissue with Targeted Mass Spectrometry. J. Proteome Res. 2020, 19, 4179–4190. [Google Scholar] [CrossRef]
- Dekker, L.J.M.; Zeneyedpour, L.; Snoeijers, S.; Joore, J.; Leenstra, S.; Luider, T.M. Determination of Site-Specific Phosphorylation Ratios in Proteins with Targeted Mass Spectrometry. J. Proteome Res. 2018, 17, 1654–1663. [Google Scholar] [CrossRef]
- Zetterberg, H. Review: Tau in biofluids—Relation to pathology, imaging and clinical features. Neuropathol. Appl. Neurobiol. 2017, 43, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Bergmann-Leitner, E.S.; Mease, R.M.; Duncan, E.H.; Khan, F.; Waitumbi, J.; Angov, E. Evaluation of immunoglobulin purification methods and their impact on quality and yield of antigen-specific antibodies. Malar. J. 2008, 7, 129. [Google Scholar] [CrossRef] [Green Version]
- Grodzki, A.C.; Berenstein, E. Antibody purification: Ammonium sulfate fractionation or gel filtration. Methods Mol. Biol. 2010, 588, 15–26. [Google Scholar]
- Lopez, E.; Scott, N.E.; Wines, B.D.; Hogarth, P.M.; Wheatley, A.K.; Kent, S.J.; Chung, A.W. Low pH Exposure During Immunoglobulin G Purification Methods Results in Aggregates That Avidly Bind Fcγ Receptors: Implications for Measuring Fc Dependent Antibody Functions. Front. Immunol. 2019, 10, 2415. [Google Scholar] [CrossRef]
- Singh, V.; Stoop, M.P.; Stingl, C.; Luitwieler, R.L.; Dekker, L.J.; van Duijn, M.M.; Kreft, K.L.; Luider, T.M.; Hintzen, R.Q. Cerebrospinal-fluid-derived Immunoglobulin G of Different Multiple Sclerosis Patients Shares Mutated Sequences in Complementarity Determining Regions. Mol. Cell. Proteom. 2013, 12, 3924–3934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broodman, I.; De Costa, D.; Stingl, C.; Dekker, L.J.M.; van Duijn, M.M.; Lindemans, J.; Van Klaveren, R.J.; Luider, T.M. Mass spectrometry analyses of κ and λ fractions result in increased number of complementarity-determining region identifications. Proteomics 2012, 12, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Barnidge, D.R.; Dasari, S.; Botz, C.M.; Murray, D.H.; Snyder, M.R.; Katzmann, J.A.; Dispenzieri, A.; Murray, D.L. Using Mass Spectrometry to Monitor Monoclonal Immunoglobulins in Patients with a Monoclonal Gammopathy. J. Proteome Res. 2014, 13, 1419–1427. [Google Scholar] [CrossRef]
- Zarling, A.L.; Polefrone, J.M.; Evans, A.M.; Mikesh, L.M.; Shabanowitz, J.; Lewis, S.T.; Engelhard, V.H.; Hunt, D.F. Identification of class I MHC-associated phosphopeptides as targets for cancer immunotherapy. Proc. Natl. Acad. Sci. USA 2006, 103, 14889–14894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhard, V.H.; Obeng, R.C.; Cummings, K.L.; Petroni, G.R.; Ambakhutwala, A.L.; Chianese-Bullock, K.A.; Smith, K.T.; Lulu, A.; Varhegyi, N.; Smolkin, M.E.; et al. MHC-restricted phosphopeptide antigens: Preclinical validation and first-in-humans clinical trial in participants with high-risk melanoma. J. Immunother. Cancer 2020, 8, e000262. [Google Scholar] [CrossRef] [PubMed]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef] [Green Version]
- Assiddiq, B.F.; Tan, K.Y.; Toy, W.; Chan, S.P.; Chong, P.K.; Lim, Y.P. EGFR S1166 Phosphorylation Induced by a Combination of EGF and Gefitinib Has a Potentially Negative Impact on Lung Cancer Cell Growth. J. Proteome Res. 2012, 11, 4110–4119. [Google Scholar] [CrossRef]
- Doll, S.; Urisman, A.; Oses-Prieto, J.A.; Arnott, D.; Burlingame, A.L. Quantitative Proteomics Reveals Fundamental Regulatory Differences in Oncogenic HRAS and Isocitrate Dehydrogenase (IDH1) Driven Astrocytoma. Mol. Cell. Proteom. MCP 2017, 16, 39–56. [Google Scholar] [CrossRef] [Green Version]
- Uceda-Castro, R.; van Asperen, J.V.; Vennin, C.; Sluijs, J.A.; van Bodegraven, E.J.; Margarido, A.S.; Robe, P.A.J.; van Rheenen, J.; Hol, E.M. GFAP splice variants fine-tune glioma cell invasion and tumour dynamics by modulating migration persistence. Sci. Rep. 2022, 12, 424. [Google Scholar] [CrossRef]
- Cirkel, A.; Wandinger, K.-P.; Ditz, C.; Leppert, J.; Hanker, L.; Cirkel, C.; Neumann, A.; Brocke, J.; Höftberger, R.; Komorowski, L.; et al. Paraneoplastic encephalomyeloradiculits with multiple autoantibodies against ITPR-1, GFAP and MOG: Case report and literature review. Neurol. Res. Pract. 2021, 3, 48. [Google Scholar] [CrossRef]
- Ahmadipour, Y.; Gembruch, O.; Pierscianek, D.; Sure, U.; Jabbarli, R. Does the expression of glial fibrillary acid protein (GFAP) stain in glioblastoma tissue have a prognostic impact on survival? Neurochirurgie 2020, 66, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Imakamura, Y.; Takeda, M.; Nishimura, T.; Inagaki, N. Glial Fibrillary Acidic Protein: Dynamic Property and Regulation by Phosphorylation. Brain Pathol. 1994, 4, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Agilent. Workflow Automation for LC/MS: In-Solution Protein Digestion, Peptide Cleanup, and Strong Cation-Exchange Fractionation of Peptides Enabled by AssayMAP Technology. Available online: https://www.agilent.com/cs/library/applications/5991-3602EN.pdf (accessed on 16 March 2016).
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.R-project.org/ (accessed on 22 April 2022).
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- van Duijn, M.M.; Dekker, L.J.; Zeneyedpour, L.; Smitt, P.A.; Luider, T.M. Immune Responses Are Characterized by Specific Shared Immunoglobulin Peptides That Can Be Detected by Proteomic Techniques. J. Biol. Chem. 2010, 285, 29247–29253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, F.C.; Westwood, O.M. Practical Immunology; Blackwell Publishing Company: Oxford, UK, 2008; pp. 115–162. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Individual | Age | Gender | Treatment | Molecular Diagnosis | Tumor Type and WHO Classification | Experiment |
|---|---|---|---|---|---|---|---|
| Donor-44 | D1 | 53 | Male | - | - | Healthy | GFAP and EGFR |
| Donor-45 | D2 | 52 | Male | - | - | Healthy | GFAP |
| Pnop-01 | P1 | 61 | Male | Before | IDH-mutant p1p/19q No EGFR amplification | Oligodendroglioma (grade 2) | GFAP |
| Pnop-02 | P1 | 61 | Male | After surgery and therapy | IDH-mutant p1p/19q No EGFR amplification | Oligodendroglioma (grade 2) | GFAP |
| Pnop-09 | P2 | 47 | Female | Before | IDH-mutant No EGFR amplification | Glioblastoma (grade 4) | GFAP |
| Pnop-13 | P3 | 75 | Male | Before | Not available | Glioblastoma (grade 4) | GFAP and EGFR |
| Pnop-14 | P3 | 75 | Male | After surgery and therapy | Not available | Glioblastoma (grade 4) | EGFR |
| Pnop-17 | P4 | 66 | Male | Before | Not available | Glioblastoma (grade 4) | EGFR |
| Pnop-30 | P5 | 41 | Female | Before | p1p/19q | Oligodendroglioma (grade 3) | GFAP and EGFR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeneyedpour, L.; Stingl, C.; Kros, J.M.; Sillevis Smitt, P.A.E.; Luider, T.M. Novel Antibody–Peptide Binding Assay Indicates Presence of Immunoglobulins against EGFR Phospho-Site S1166 in High-Grade Glioma. Int. J. Mol. Sci. 2022, 23, 5061. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095061
Zeneyedpour L, Stingl C, Kros JM, Sillevis Smitt PAE, Luider TM. Novel Antibody–Peptide Binding Assay Indicates Presence of Immunoglobulins against EGFR Phospho-Site S1166 in High-Grade Glioma. International Journal of Molecular Sciences. 2022; 23(9):5061. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095061
Chicago/Turabian StyleZeneyedpour, Lona, Christoph Stingl, Johan M. Kros, Peter A. E. Sillevis Smitt, and Theo M. Luider. 2022. "Novel Antibody–Peptide Binding Assay Indicates Presence of Immunoglobulins against EGFR Phospho-Site S1166 in High-Grade Glioma" International Journal of Molecular Sciences 23, no. 9: 5061. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095061