
Impact of Endurance Training on Regeneration of Axons, Glial Cells, and Inhibitory Neurons after Spinal Cord Injury: A Link between Functional Outcome and Regeneration Potential within the Lesion Site and in Adjacent Spinal Cord Tissue
 , ,
, ,
Abstract
:1. Introduction
2. Results
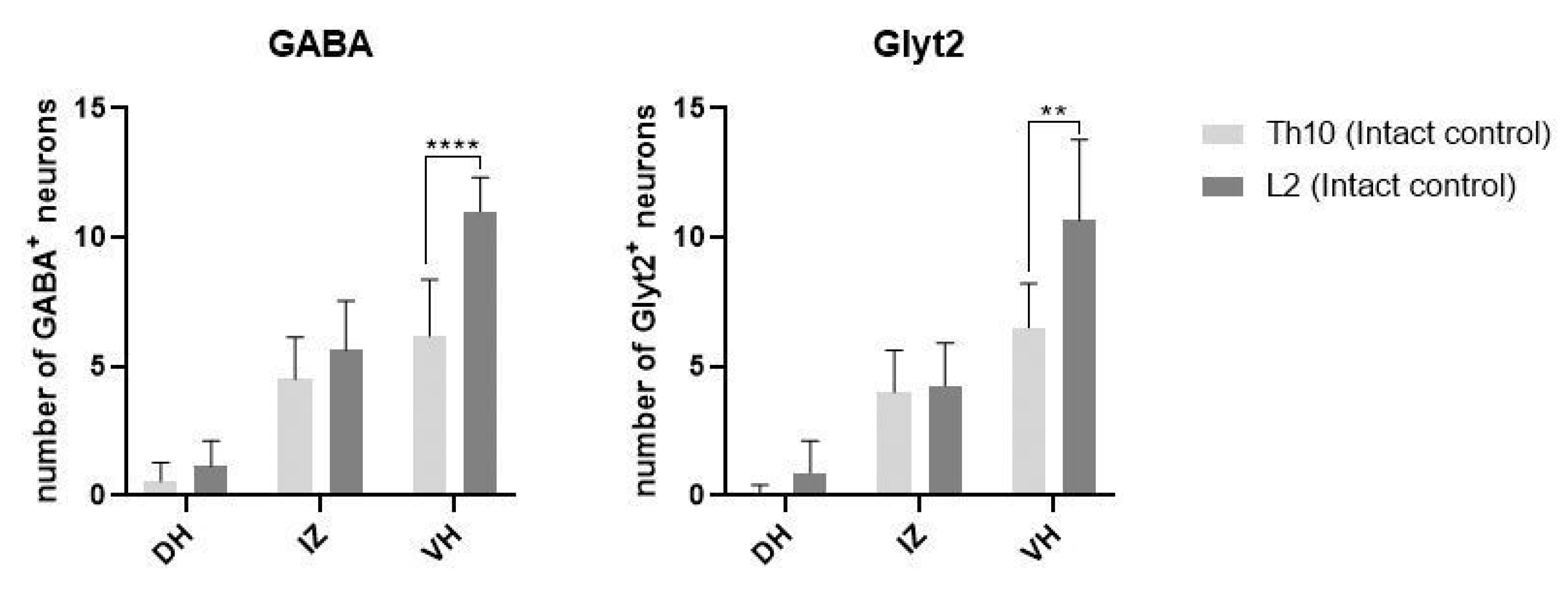
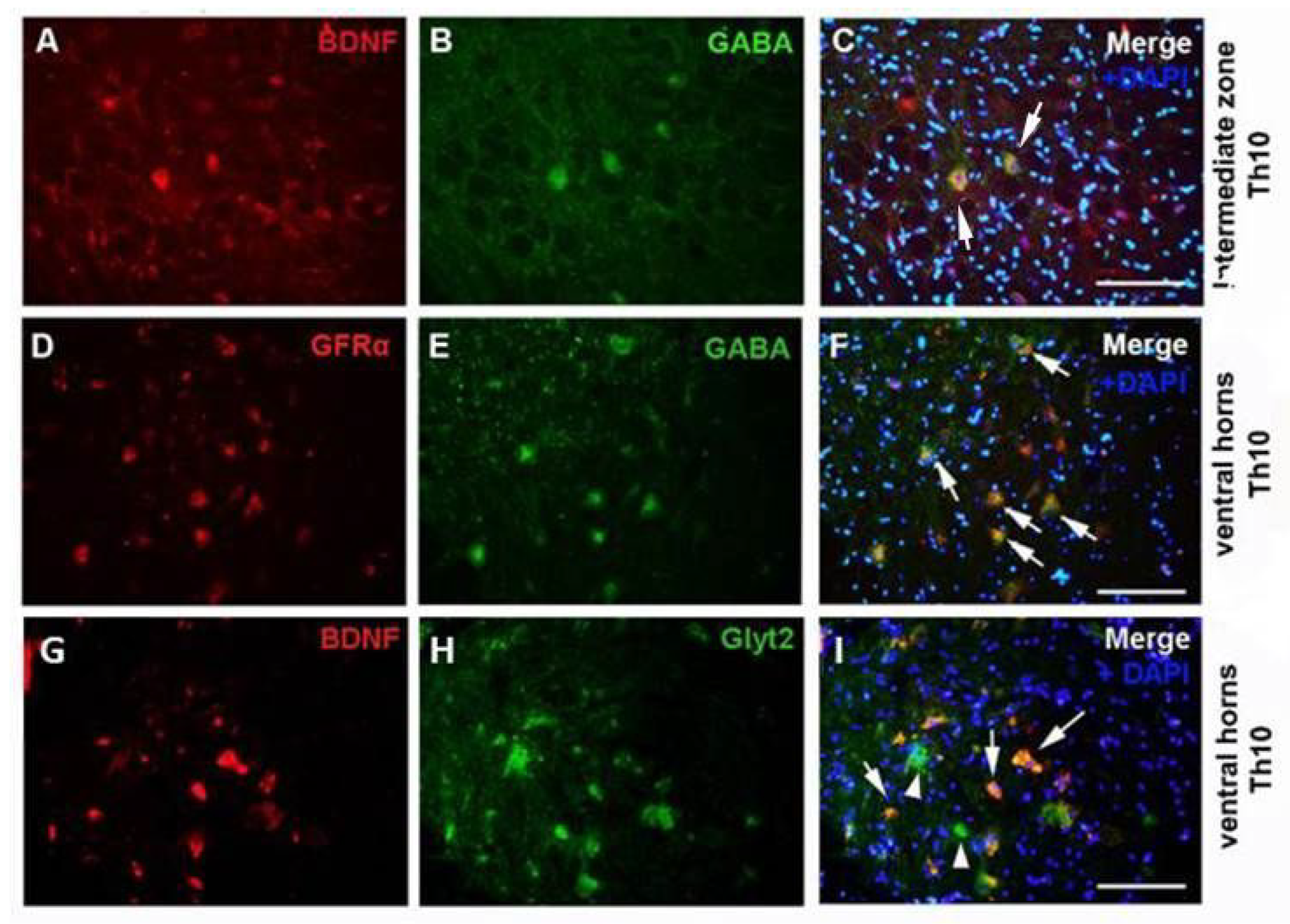
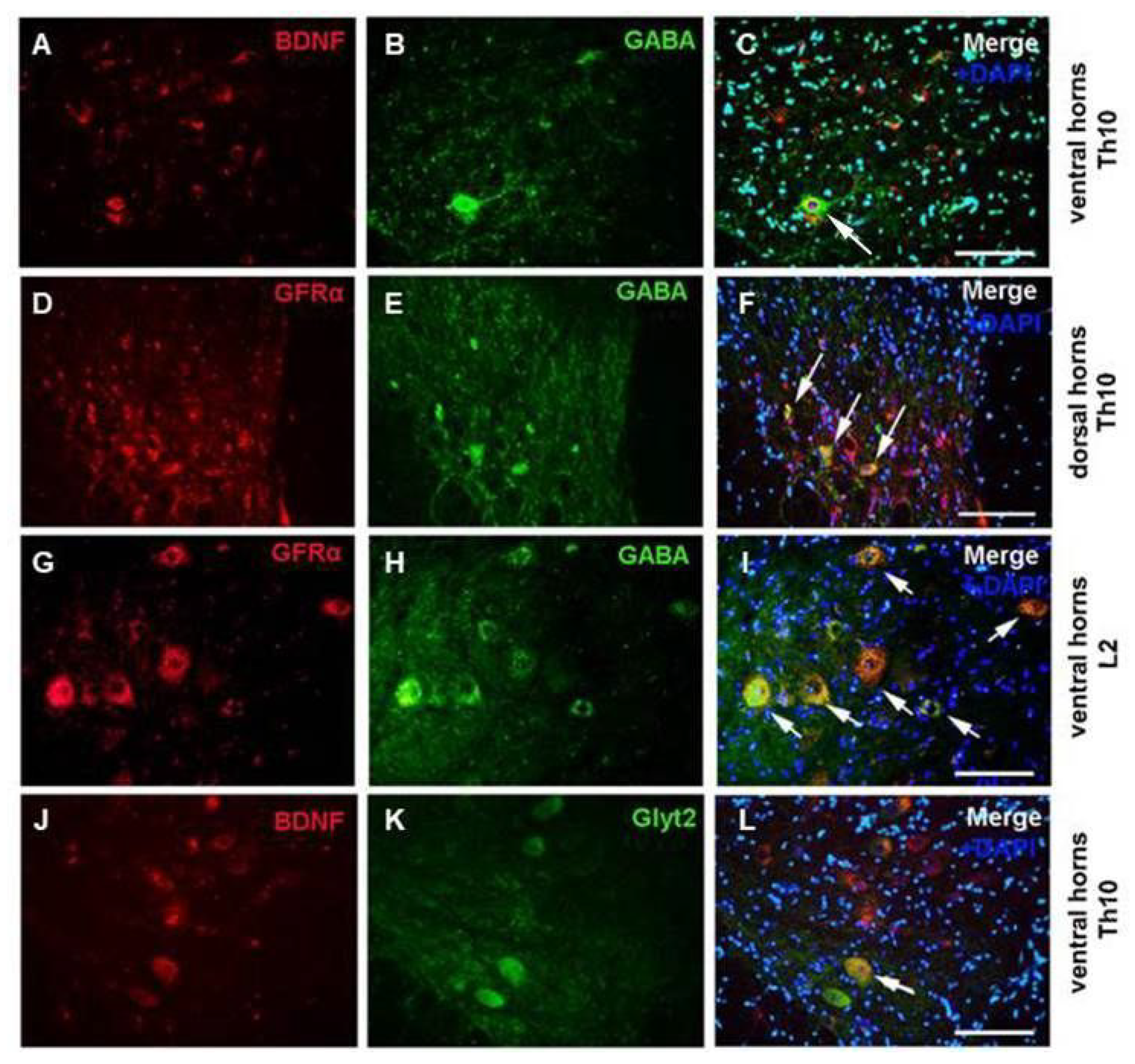
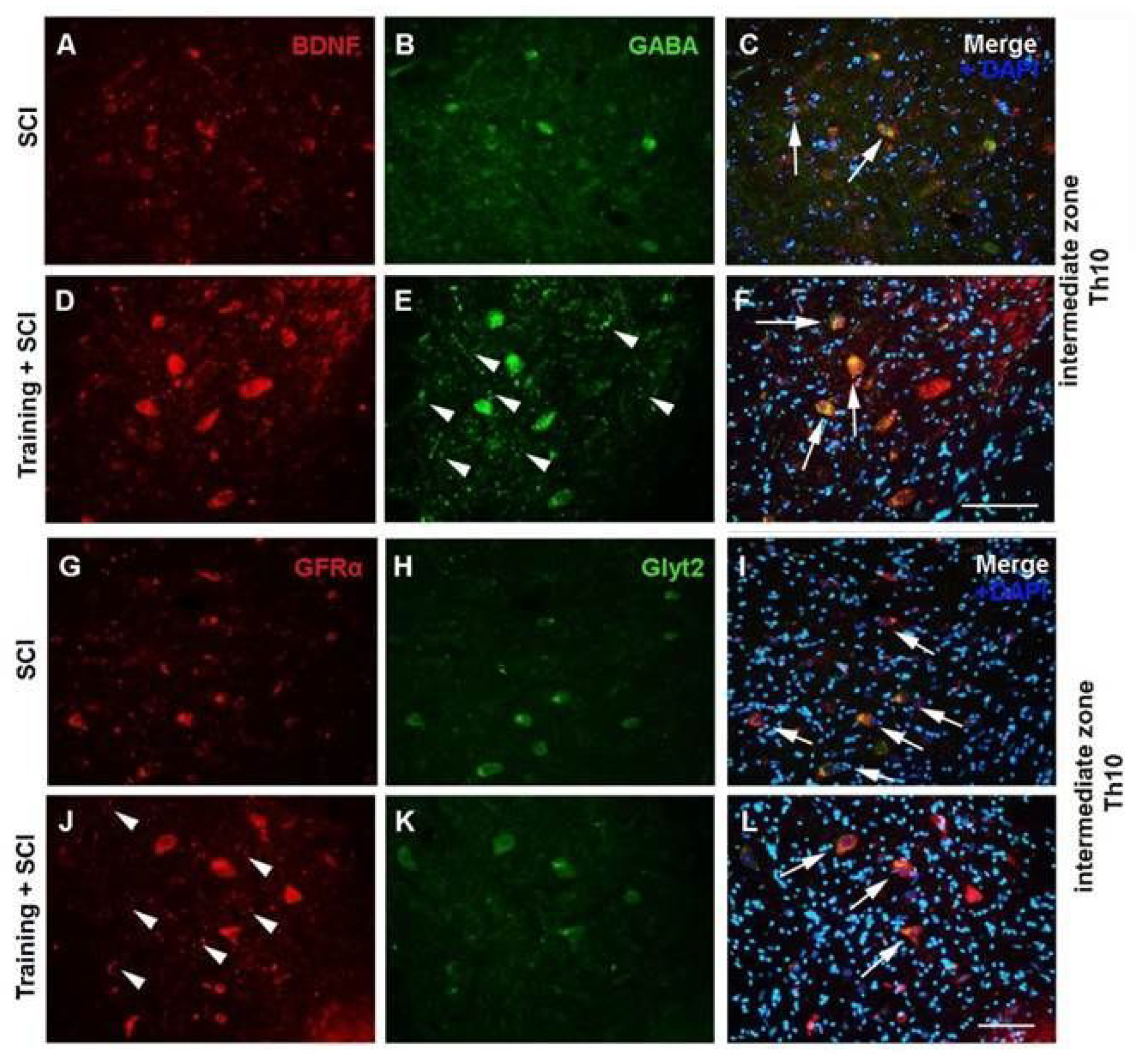
2.1. Immunoexpression of Growth Factors in Population of GABA/Glycinergic Neurons Affected by Training and Th9 Compression
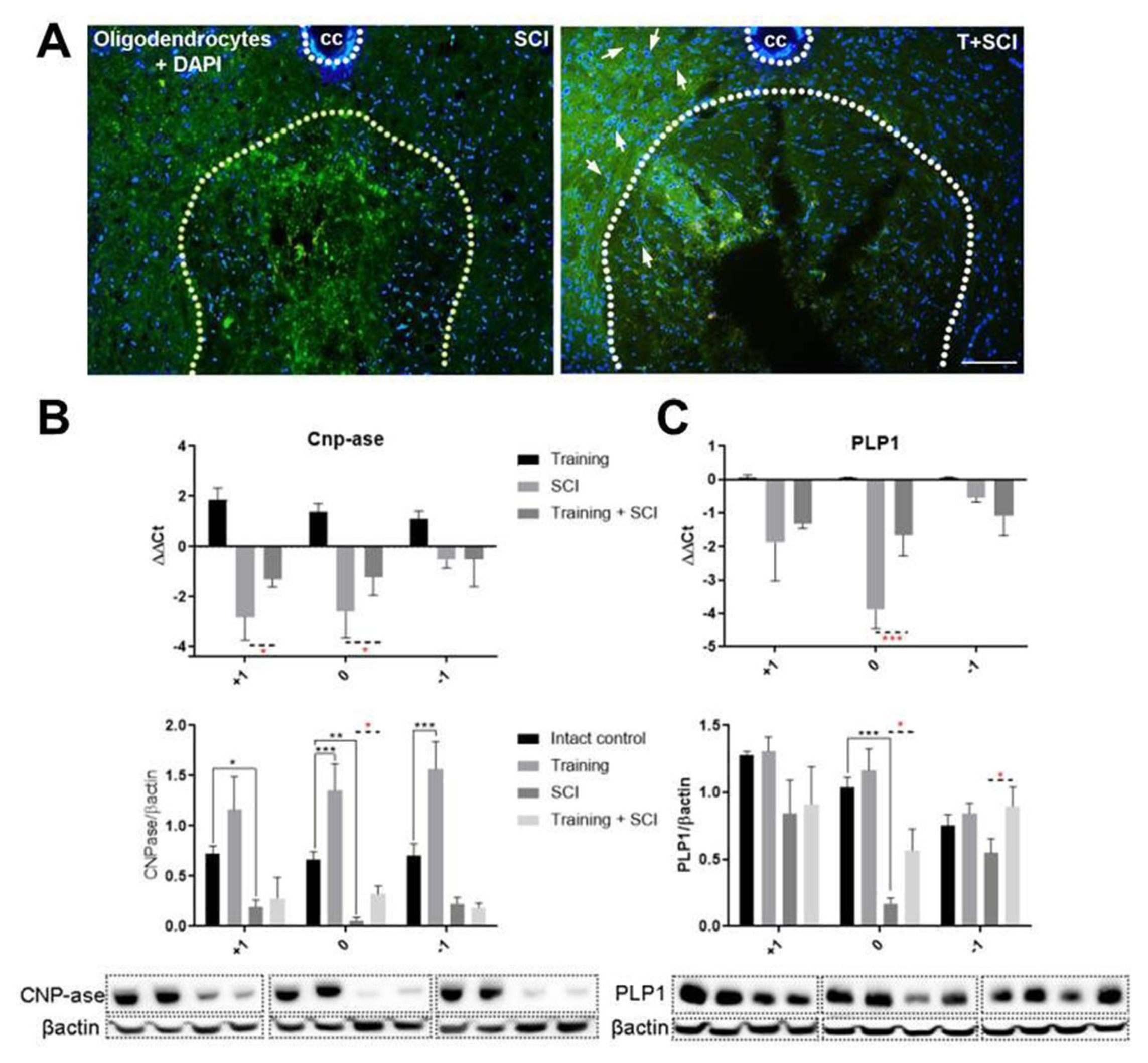
2.2. Endurance Training Applied before Th9 Compression Upregulates Mature and Immature Oligodendrocytes at Lesion Site and in Its Surrounding
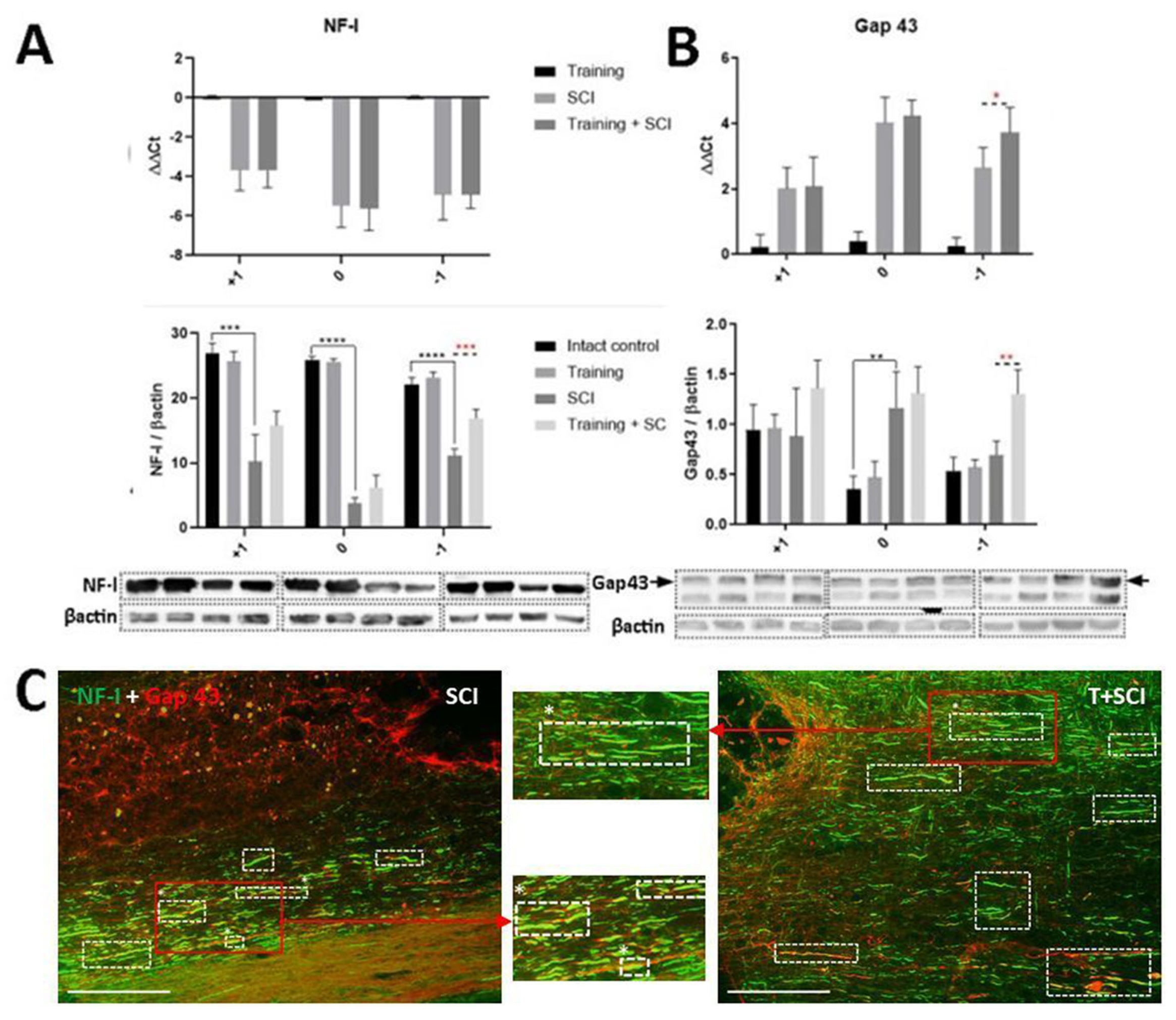
2.3. The Upregulation of Neurofilaments and Outgrowing Axons in Spinal Cord after Endurance Training + Th9 Compression and 6 Weeks Survival
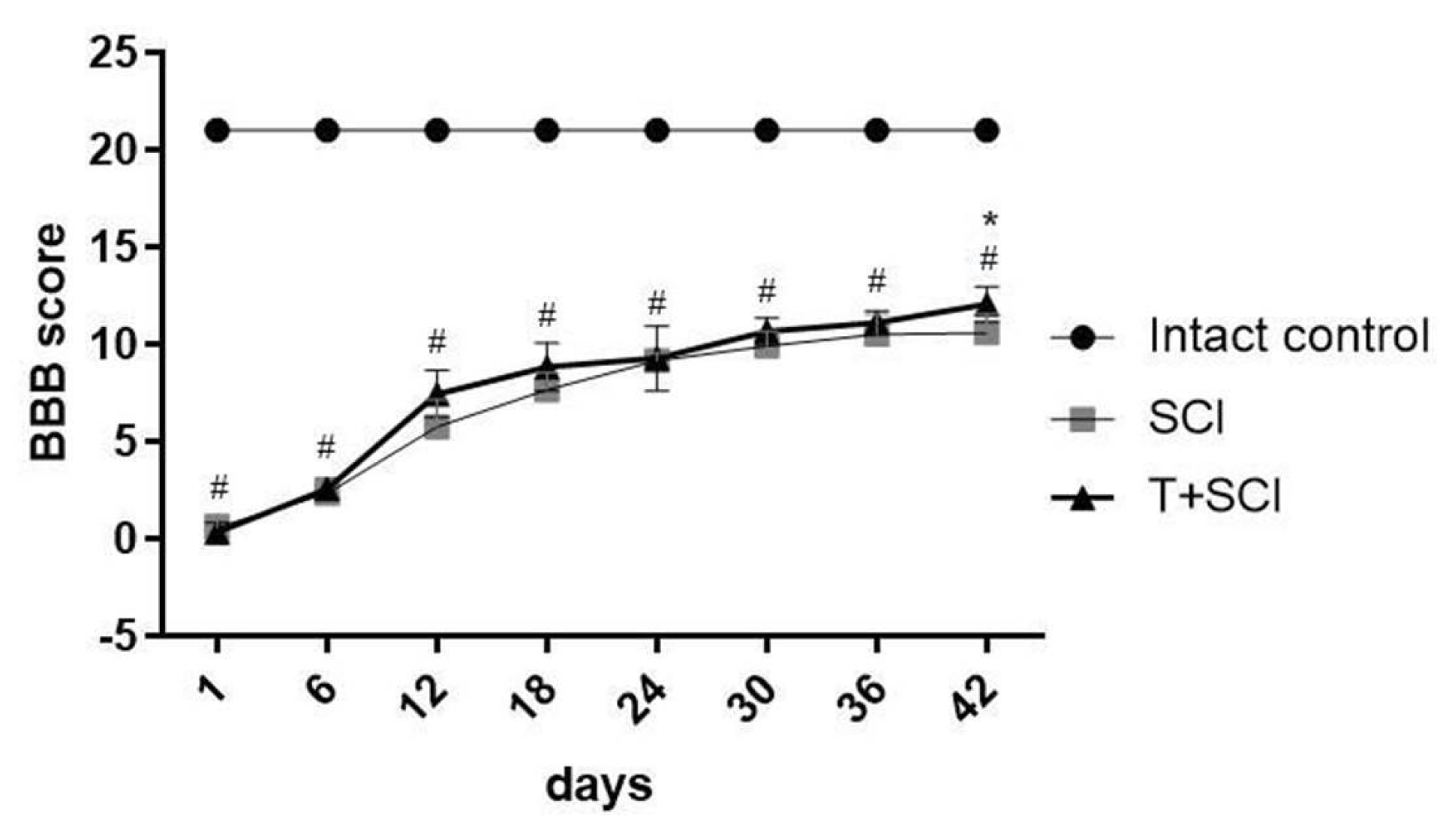
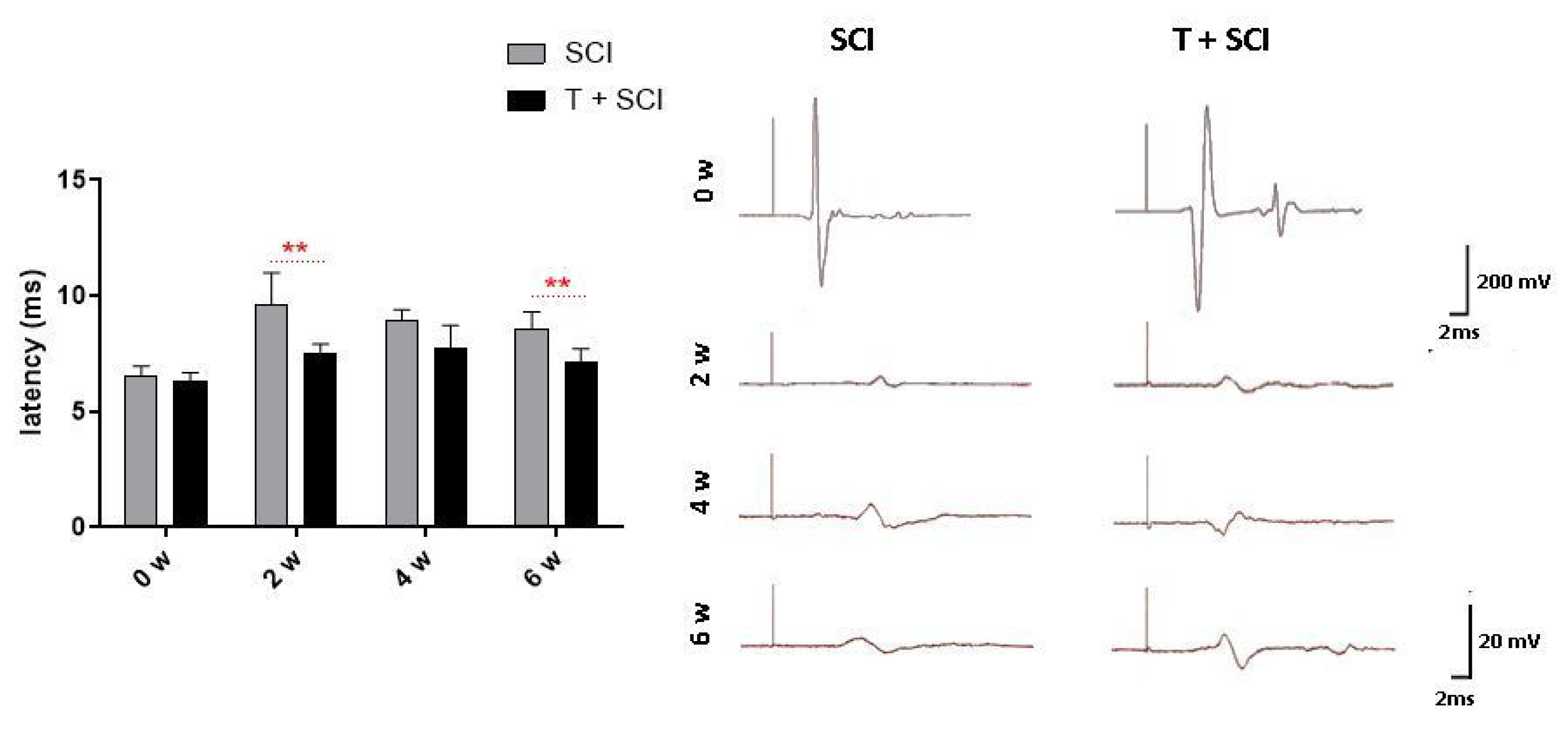
2.4. Endurance Training before SCI Improves Micturition, BBB Score, and Transcranial Motor Evoked Potentials
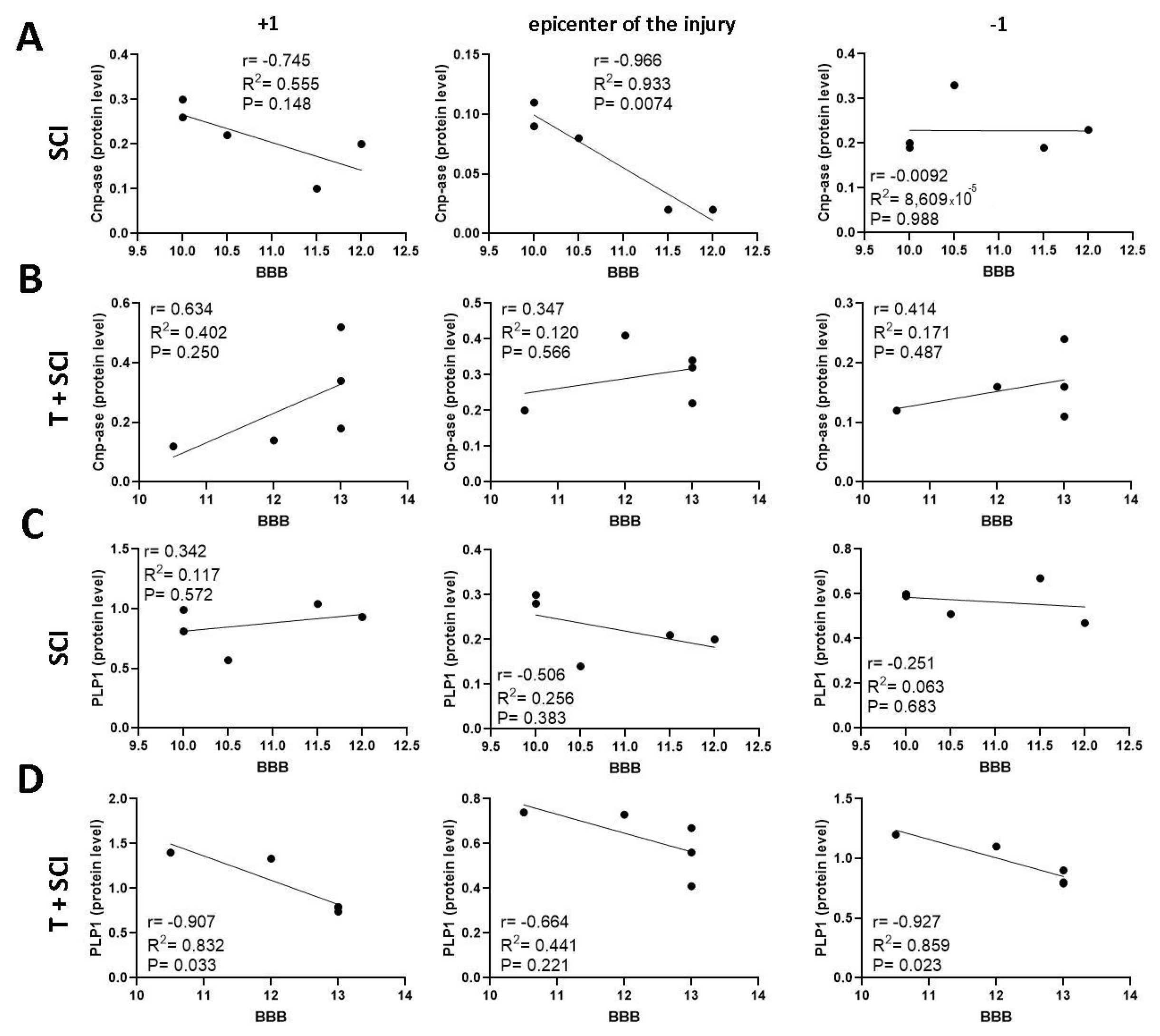
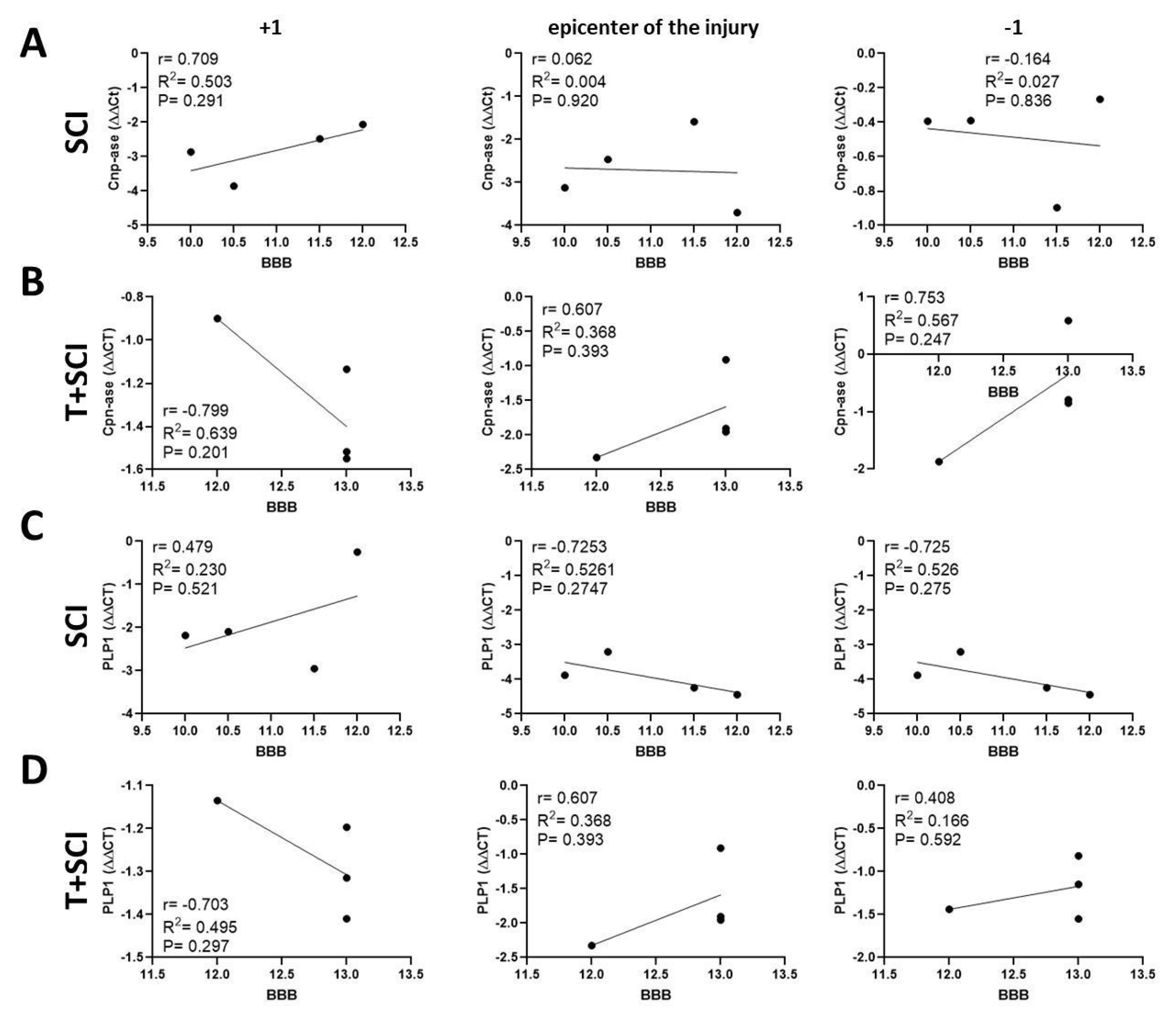
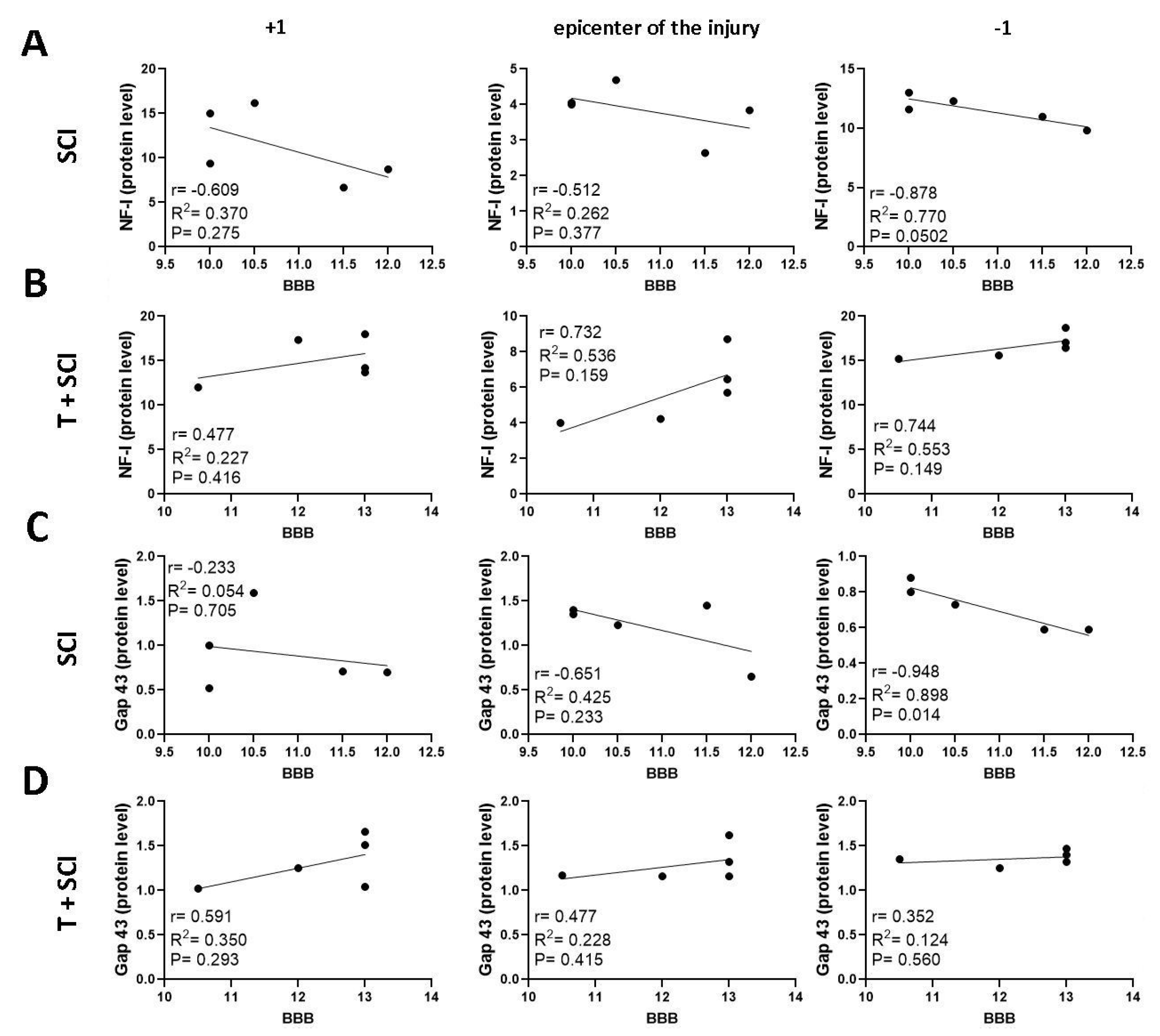
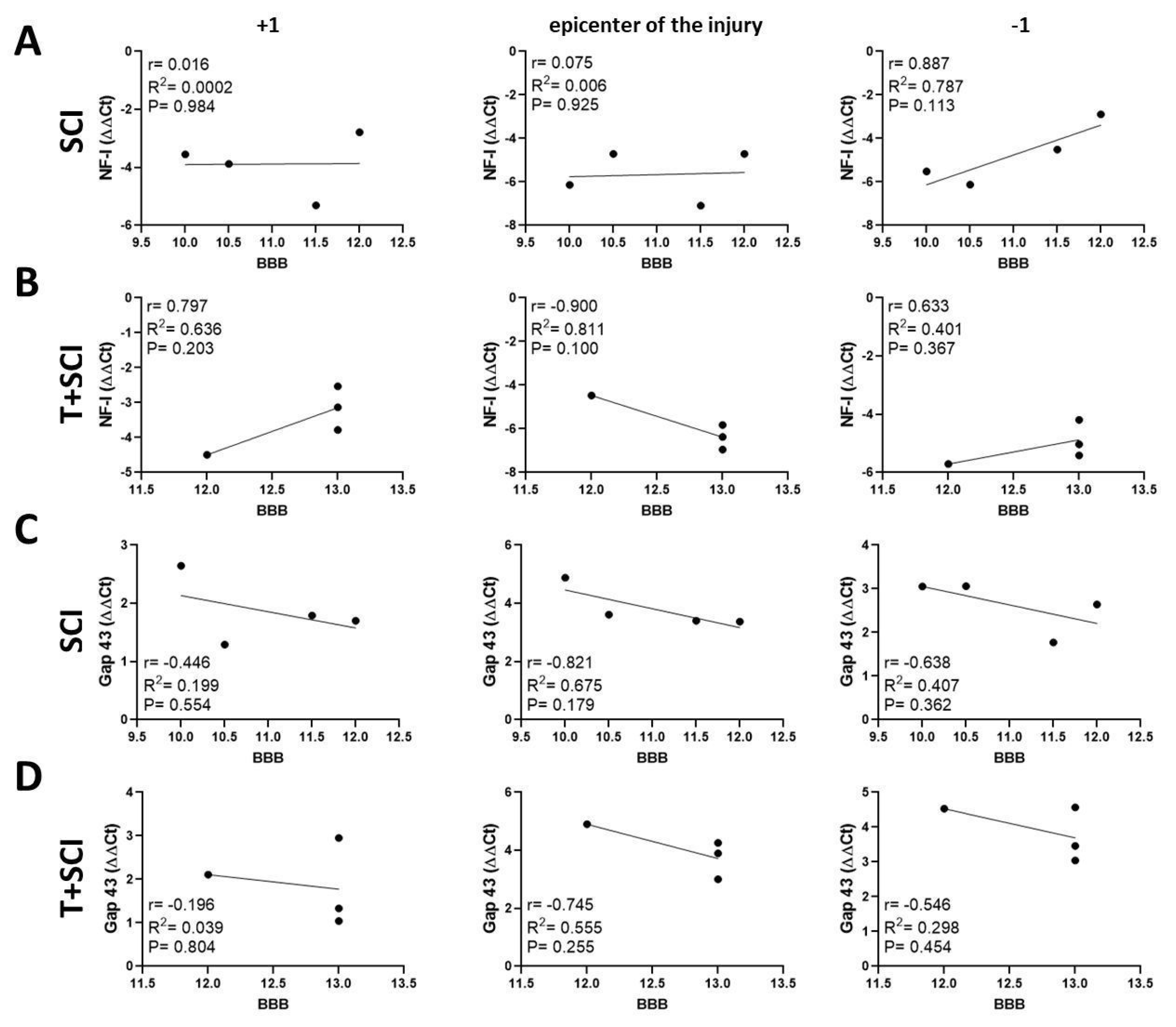
2.5. Correlation between BBB Neurological Score, Protein Levels, and Gene Expression for Axonal and Glial Markers
3. Discussion
3.1. Modulation of Neurotrophic Support in Inhibition Neurons after Endurance Training, SCI, and T + SCI
3.2. Physical Activity Applied before SCI Induced Glial and Axonal Restoration
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Endurance Training
4.3. Surgical Procedure
4.4. Gene Expression Analysis (RT-PCR)
4.5. Western Blot Analysis
4.6. Immunohistochemical Analysis
4.7. GABA/Glycinergic Cell Quantification
4.8. BBB Locomotor Testing
4.9. Transcranial Motor Evoked Potentials (tcMEPs)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arrieta, H.; Rezola-Pardo, C.; Echeverria, I.; Iturburu, M.; Gil, S.M.; Yanguas, J.J.; Irazusta, J.; Rodriguez-Larrad, A. Physical activity and fitness are associated with verbal memory, quality of life and depression among nursing home residents: Preliminary data of a randomized controlled trial. BMC Geriatr. 2018, 18, 80. [Google Scholar] [CrossRef] [PubMed]
- Małczyńska-Sims, P.; Chalimoniuk, M.; Sułek, A. The Effect of Endurance Training on Brain-Derived Neurotrophic Factor and Inflammatory Markers in Healthy People and Parkinson’s Disease. A Narrative Review. Front. Physiol. 2020, 11, 578981. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K. The diseasome of physical inactivity–and the role of myokines in muscle–fat cross talk. J. Physiol. 2009, 587, 5559–5568. [Google Scholar] [CrossRef]
- Moon, H.Y.; Becke, A.; Berron, D.; Becker, B.; Sah, N.; Benoni, G.; Janke, E.; Lubejko, S.; Greig, N.H.; Mattison, J.A.; et al. Running induced systemic cathepsin b secretion is associated with memory function. Cell Metab. 2016, 24, 332–340. [Google Scholar] [CrossRef]
- Schön, M.; Malenovská, K.M.; Nemec, M.; Laiferová, N.A.; Straka, I.; Košutzká, Z.; Matejička, P.; Valkovič, P.; Ukropec, J.; Ukropcová, B. Acute endurance exercise modulates growth differentiation factor 11 in cerebrospinal fluid of healthy young adults. Front. Endocrinol. 2023, 14, 1137048. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Action Plan on Physical Activity 2018–2030: More Active People for a Healthier World. World Health Organization. 2018. Available online: https://apps.who.int/iris/handle/10665/272722 (accessed on 28 March 2023).
- Quintanar, J.L.; del Carmen Díaz-Galindo, M.; Calderón-Vallejo, D.; Jaramillo-González, F. Trophic factors in patients with spinal cord injury. In Diagnosis and Treatment of Spinal Cord Injury; Rajendram, R., Preedy, V.R., Martin, C.R., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 475–487. [Google Scholar] [CrossRef]
- Boyce, V.S.; Mendell, L.M. Neurotrophic Factors in Spinal Cord Injury. In Neurotrophic Factors; Lewin, G.R., Carter, B.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 443–460. [Google Scholar]
- Dravid, A.; O’Carroll, S.J.; Svirskis, D. Neurotrophins and their role in axonal outgrowth following spinal cord injury. In Cellular, Molecular, Physiological, and Behavioral Aspects of Spinal Cord Injury; Rajendram, R., Preedy, V.R., Martin, C.R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2022; pp. 215–227. [Google Scholar] [CrossRef]
- Bothwell, M. NGF, BDNF, NT3, and NT4. In Neurotrophic Factors; Lewin, G.R., Carter, B.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3–16. [Google Scholar] [CrossRef]
- Smith, P.A. BDNF: No gain without pain? Neuroscience 2014, 283, 107–123. [Google Scholar] [CrossRef]
- Silva, A.C.; Lobo, J.M.S. Cytokines and Growth Factors. Adv. Biochem. Eng. Biotechnol. 2020, 171, 87–113. [Google Scholar] [CrossRef]
- Keefe, K.M.; Sheikh, I.S.; Smith, G.M. Targeting Neurotrophins to Specific Populations of Neurons: NGF, BDNF, and NT-3 and Their Relevance for Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef]
- Kiss Bimbova, K.; Bacova, M.; Kisucka, A.; Galik, J.; Zavacky, P.; Lukacova, N. Activation of Three Major Signaling Pathways After Endurance Training and Spinal Cord Injury. Mol. Neurobiol. 2022, 59, 950–967. [Google Scholar] [CrossRef]
- Li, X.; Wu, Q.; Xie, C.; Wang, C.; Wang, Q.; Dong, C.; Fang, L.; Ding, J.; Wang, T. Blocking of BDNF-TrkB signaling inhibits the promotion effect of neurological function recovery after treadmill training in rats with spinal cord injury. Spinal Cord 2018, 57, 65–74. [Google Scholar] [CrossRef]
- Mekki, M.; Delgado, A.D.; Fry, A.; Putrino, D.; Huang, V. Robotic Rehabilitation and Spinal Cord Injury: A Narrative Review. Neurotherapeutics 2018, 15, 604–617. [Google Scholar] [CrossRef] [PubMed]
- Idrisova, K.F.; Rizvanov, A.A.; Zeinalova, A.K.; Masgutova, G.A.; Bogov, A.A.; Allegrucci, C.; Syromiatnikova, V.Y.; Salafutdinov, I.I.; Garanina, E.E.; Andreeva, D.I. Application of neurotrophic and proangiogenic factors as therapy after peripheral nervous system injury. Neural. Regen. Res. 2022, 17, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, E. Interneuronal relay in spinal pathways from proprioceptors. Prog. Neurobiol. 1992, 38, 335–378. [Google Scholar] [CrossRef] [PubMed]
- Kucharíková, A.; Schreiberová, A.; Závodská, M.; Gedrová, Š.; Hricová, Ľ.; Pavel, J.; Gálik, J.; Maršala, M.; Lukáčová, N. Repeated Baclofen treatment ameliorates motor dysfunction, suppresses reflex activity and decreases the expression of signaling proteins in reticular nuclei and lumbar motoneurons after spinal trauma in rats. Acta Histochem. 2014, 116, 344–353. [Google Scholar] [CrossRef]
- Sartori, A.M.; Hofer, A.S.; Scheuber, M.I.; Rust, R.; Kessler, T.M.; Schwab, M.E. Slow development of bladder malfunction parallels spinal cord fiber sprouting and interneurons’ loss after spinal cord transection. Exp. Neurol. 2022, 348, 113937. [Google Scholar] [CrossRef]
- Kjaerulff, O.; Kiehn, O. Distribution of networks generating and coordinating locomotor activity in the neonatal rat spinal cord in vitro: A lesion study. J. Neurosci. 1996, 16, 5777–5794. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A sensitive and reliable locomotor rating scale for open field testing in rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef]
- Kuhn, S.; Gritti, L.; Crooks, D.; Dombrowski, Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells 2019, 8, 1424. [Google Scholar] [CrossRef]
- Bimbova, K.; Bacova, M.; Kisucka, A.; Pavel, J.; Galik, J.; Zavacky, P.; Marsala, M.; Stropkovska, A.; Fedorova, J.; Papcunova, S. A Single Dose of Atorvastatin Applied Acutely after Spinal Cord Injury Suppresses Inflammation, Apoptosis, and Promotes Axon Outgrowth, Which Might Be Essential for Favorable Functional Outcome. Int. J. Mol. Sci. 2018, 19, 1106. [Google Scholar] [CrossRef]
- Imai, T.; Katoh, H.; Suyama, K.; Kuroiwa, M.; Yanagisawa, S.; Watanabe, M. Amiloride Promotes Oligodendrocyte Survival and Remyelination after Spinal Cord Injury in Rats. J. Clin. Med. 2018, 7, 46. [Google Scholar] [CrossRef]
- Wu, B.; Sun, L.; Li, P.; Tian, M.; Luo, Y.; Ren, X. Transplantation of oligodendrocyte precursor cells improves myelination and promotes functional recovery after spinal cord injury. Injury 2012, 43, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Stropkovska, A.; Kisucka, A.; Bimbova, K.; Bacova, M.; Galik, J.; Medvecky, L.; Sulla, I.; Karasova, M.; Lukacova, N. Combined therapy (Rho-A-kinase inhibitor and chitosan/collagen porous scaffold) provides a supportive environment for endogenous regenerative processes after spinal cord trauma. Arch. Ital. Biol. 2021, 159, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Yuan, A.; Veeranna, M.V.R.; Nixon, R.A. Neurofilaments and Neurofilament Proteins in Health and Disease. Cold Spring Harb. Perspect. Biol. 2017, 9, a018309. [Google Scholar] [CrossRef]
- Langfort, J.; Barańczuk, E.; Pawlak, D.; Chalimoniuk, M.; Lukacova, N.; Marsala, J.; Górski, J. The Effect of Endurance Training on Regional Serotonin Metabolism in the Brain During Early Stage of Detraining Period in the Female Rat. Cell. Mol. Neurobiol. 2006, 26, 1325–1340. [Google Scholar] [CrossRef]
- Chalimoniuk, M.; Chrapusta, S.J.; Lukacova, N.; Langfort, J. Endurance training upregulates the nitric oxide/soluble guanylyl cyclase/cyclic guanosine 3′,5′-monophosphate pathway in the striatum, midbrain and cerebellum of male rats. Brain Res. 2015, 1618, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Skup, M.; Dwornik, A.; Macias, M.; Sulejczak, D.; Wiater, M.; Czarkowska-Bauch, J. Long-Term Locomotor Training Up-Regulates TrkBFL Receptor-like Proteins, Brain-Derived Neurotrophic Factor, and Neurotrophin 4 with Different Topographies of Expression in Oligodendroglia and Neurons in the Spinal Cord. Exp. Neurol. 2002, 176, 289–307. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Pedersen, M.; Krabbe, K.S.; Bruunsgaard, H.; Matthews, V.B.; Febbraio, M.A. Role of exercise-induced brain-derived neurotrophic factor production in the regulation of energy homeostasis in mammals. Exp. Physiol. 2009, 94, 1153–1160. [Google Scholar] [CrossRef]
- Kiehn, O. Locomotor circuits in the mammalian spinal cord. Annu. Rev. Neurosci. 2006, 29, 279–306. [Google Scholar] [CrossRef]
- Kiehn, O. Decoding the organization of spinal circuits that control locomotion. Nat. Rev. Neurosci. 2016, 17, 224–238. [Google Scholar] [CrossRef]
- Brown, A.G. The spinocervical tract. Prog. Neurobiol. 1981, 17, 59–96. [Google Scholar] [CrossRef]
- Gosgnach, S.; Lanuza, G.; Butt, S.; Saueressig, H.; Zhang, Y.; Velasquez, T.; Riethmacher, D.; Callaway, E.M.; Kiehn, O.; Goulfing, M. V1 spinal neurons regulate the speed of vertebrate locomotor outputs. Nature 2006, 440, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Callahan, R.A.; Roberts, R.; Sengupta, M.; Kimura, Y.; Higashijima, S.; Bagnall, M.W. Spinal V2b neurons reveal a role for ipsilateral inhibition in speed control. eLife 2019, 8, e47837. [Google Scholar] [CrossRef] [PubMed]
- Britz, O.; Zhang, J.; Grossmann, K.S.; Dyck, J.; Kim, J.C.; Dymecki, S.; Gosgnach, S.; Goulding, M. A genetically defined asymmetry underlies the inhibitory control of flexor–extensor locomotor movements. eLife 2015, 4, e04718. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Song, X.; Fang, L.; Ding, J.; Qi, W.; Dong, C.H.; Wang, S.; Wu, J.; Wang, T.; Wu, Q. Body Weight-Supported Treadmill Training Ameliorates Motoneuronal Hyperexcitability by Increasing GAD-65/67 and KCC2 Expression via TrkB Signaling in Rats with Incomplete Spinal Cord Injury. Neurochem. Res. 2022, 47, 1679–1691. [Google Scholar] [CrossRef]
- Steeves, J.D.; Jordan, L.M. Localization of a descending pathway in the spinal cord which is necessary for controlled treadmill locomotion. Neurosci. Lett. 1980, 20, 283–288. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Skinner, R.D. The mesencephalic locomotor region: I Activation of a medullary projection site. Brain Res. 1987, 411, 1–12. [Google Scholar] [CrossRef]
- Matsuyama, K.; Mori, F.; Nakajima, K.; Drew, T.; Aoki, M.; Mori, S. Locomotor role of the corticoreticular-reticulospinal-spinal interneuronal system. Prog. Brain Res. 2004, 143, 239–249. [Google Scholar] [CrossRef]
- Lee, S.; Nam, H.; Joo, K.M.; Lee, S.H. Advances in Neural Stem Cell Therapy for Spinal Cord Injury: Safety, Efficacy, and Future Perspectives. Neurospine 2022, 19, 946–960. [Google Scholar] [CrossRef]
- Su, Z.; Niu, W.; Liu, M.L.; Zou, Y.; Zhang, C.L. In vivo conversion of astrocytes to neurons in the injured adult spinal cord. Nat. Commun. 2014, 5, 3338. [Google Scholar] [CrossRef]
- Fawcett, J. Repair of spinal cord injuries: Where are we, where are we going? Spinal Cord 2002, 40, 615–623. [Google Scholar] [CrossRef]
- Mah, K.M.; Wu, W.; Al-Ali, H.; Sun, Y.; Han, Q.; Ding, Y.; Munoz, M.; Xu, X.M.; Lemmon, V.P.; Bixby, J.L. Compounds co-targeting kinases in axon regulatory pathways promote regeneration and behavioral recovery after spinal cord injury in mice. Exp. Neurol. 2022, 355, 114117. [Google Scholar] [CrossRef] [PubMed]
- Borghi, S.M.; Fattori, V.; Hohmann, M.S.N.; Verri, W.A. Contribution of Spinal Cord Oligodendrocytes to Neuroinflammatory Diseases and Pain. Curr. Med. Chem. 2019, 26, 5781–5810. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Karimi-Abdolrezaee, S. Microenvironmental regulation of oligodendrocyte replacement and remyelination in spinal cord injury. J. Physiol. 2016, 594, 3539–3552. [Google Scholar] [CrossRef]
- Kisucká, A.; Bimbová, K.; Bačová, M.; Gálik, J.; Lukáčová, N. Activation of Neuroprotective Microglia and Astrocytes at the Lesion Site and in the Adjacent Segments Is Crucial for Spontaneous Locomotor Recovery after Spinal Cord Injury. Cells 2021, 10, 1943. [Google Scholar] [CrossRef] [PubMed]
- Siebert, J.R.; Osterhout, D.J. Select neurotrophins promote oligodendrocyte progenitor cell process outgrowth in the presence of chondroitin sulfate proteoglycans. J. Neurosci. Res. 2021, 99, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Goebbels, S.; Oltrogge, J.H.; Kemper, R.; Heilmann, I.; Bormuth, I.; Wolfer, S.; Wichert, S.P.; Möbius, W.; Liu, X.; Lappe-Siefke, C. Elevated phosphatidylinositol 3,4,5-trisphosphate in glia triggers cell-autonomous membrane wrapping and myelination. J. Neurosci. 2010, 30, 8953–8964. [Google Scholar] [CrossRef]
- Ding, Z.; Dai, C.; Zhong, L.; Liu, R.; Gao, W.; Zhang, H.; Yin, Z. Neuregulin-1 converts reactive astrocytes toward oligodendrocyte lineage cells via upregulating the PI3K-AKT-mTOR pathway to repair spinal cord injury. Biomed. Pharmacother. 2021, 134, 111168. [Google Scholar] [CrossRef]
- Balik, V.; Šulla, I. Autonomic Dysreflexia following Spinal Cord Injury. Asian J. Neurosurg. 2022, 17, 165–172. [Google Scholar] [CrossRef]
- Lukacova, N.; Kisucka, A.; Kiss Bimbova, K.; Bacova, M.; Ileninova, M.; Kuruc, T.; Galik, J. Glial-Neuronal Interactions in Pathogenesis and Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 13577. [Google Scholar] [CrossRef]
- Cai, L.; Finkel, Z. Transcription factors promote neural regeneration after spinal cord injury. Neural Reg. Res. 2022, 17, 2439–2440. [Google Scholar] [CrossRef]
- Alibardi, L. Growth associated protein 43 and neurofilament immunolabeling in the transected lumbar spinal cord of lizard indicates limited axonal regeneration. Neural Reg. Res. 2022, 17, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- Bacova, M.; Bimbova, K.; Kisucka, A.; Lukacova, N.; Galik, J. Epidural oscillating field stimulation increases axonal regenerative capacity and myelination after spinal cord trauma. Neural Reg. Res. 2022, 17, 2730–2736. [Google Scholar] [CrossRef]
- Jing, J.-H.; Qian, J.; Zhu, N.; Chou, W.B.; Huang, X.-J. Improved differentiation of oligodendrocyte precursor cells and neurological function after spinal cord injury in rats by oscillating field stimulation. Neuroscience 2015, 303, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Weishaupt, N.; Blesch, A.; Fouad, K. BDNF: The career of a multifaceted neurotrophin in spinal cord injury. Exp. Neurol. 2012, 238, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Yazicioglu, K.; Yavuz, F.; Goktepe, A.S.; Tan, A.K. Influence of adapted sports on quality of life and life satisfaction in sport participants and non-sport participants with physical disabilities. Disabil. Health J. 2012, 5, 249–253. [Google Scholar] [CrossRef]
- Goraczko, A.; Zurek, G.; Lachowicz, M.; Kujawa, K.; Blach, W.; Zurek, A. Quality of Life after Spinal Cord Injury: A Multiple Case Study Examination of Elite Athletes. Int. J. Environ. Res. Public Health 2020, 17, 7437. [Google Scholar] [CrossRef]
- Tator, C.H. Update on the pathophysiology and pathology of acute spinal cord injury. Brain Pathol. 1995, 5, 407–413. [Google Scholar] [CrossRef]
- Min, K.J.; Jeong, H.K.; Kim, B.; Hwang, D.H.; Shin, H.Y.; Nguyen, A.T.; Kim, J.H.; Jou, I.; Kim, B.G.; Joe, E.H. Spatial and temporal correlation in progressive degeneration of neurons and astrocytes in contusion-induced spinal cord injury. J. Neuroinflamm. 2012, 9, 100. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
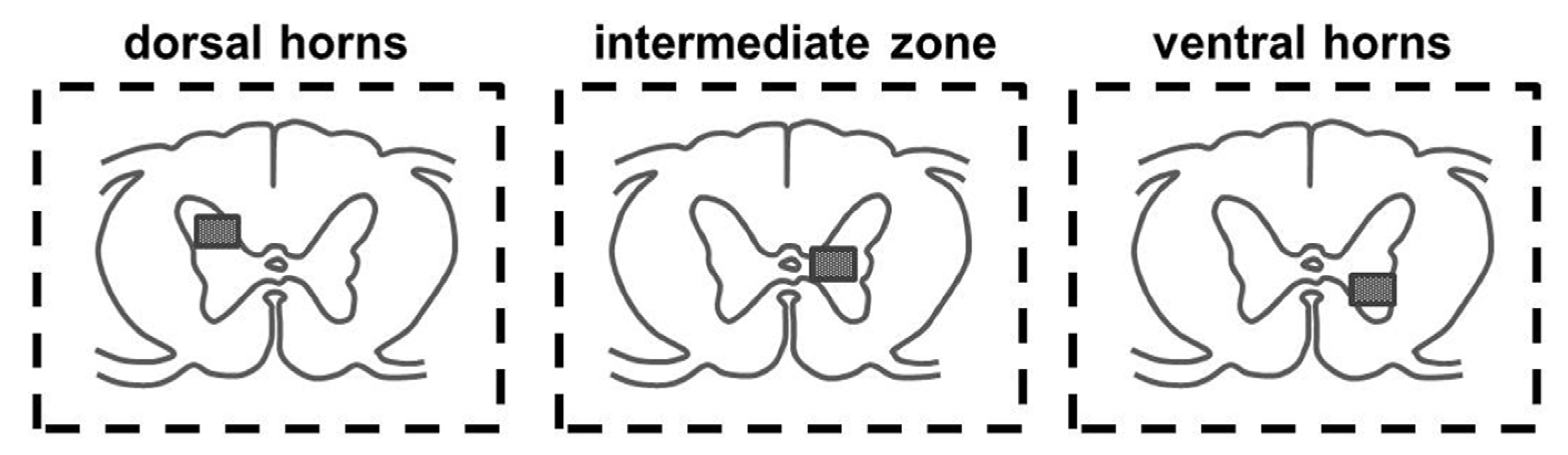{kind=link}
| Significance | ||||
|---|---|---|---|---|
| IC vs. T | IC vs. SCI | SCI vs. T + SCI | ||
| GABA (Th10 segment) | IC | T | SCI | T + SCI |
| dorsal horns | 0.70 ± 0.83 | 0.36 ± 0.67 | 0 ± 0 | 0.58 ± 0.99 |
| ns | ** | * | ||
| intermediate zone | 4.50 ± 1.65 | 4.10 ± 1.66 | 1.84 ± 1.28 | 2.10 ± 1.66 |
| ns | *** | ns | ||
| ventral horns | 6.25 ± 2.09 | 6.00 ± 1.83 | 4.35 ± 1.08 | 4.41 ± 1.51 |
| ns | ** | ns | ||
| Glyt2 (Th10 segment) | IC | T | SCI | T + SCI |
| dorsal horns | 0.10 ± 0.31 | 1.81 ± 1.60 | 0.17 ± 0.38 | 0.45 ± 0.68 |
| ** | ns | ns | ||
| intermediate zone | 4.00 ± 1.63 | 4.63 ± 1.43 | 2.91 ± 1.22 | 3.00 ± 1.10 |
| ns | ns | ns | ||
| ventral horns | 6.50 ± 1.72 | 7.27 ± 1.95 | 4.83 ± 1.53 | 5.09 ± 1.14 |
| ns | * | ns |
| Significance | ||||
|---|---|---|---|---|
| IC vs. T | IC vs. SCI | SCI vs. T + SCI | ||
| GABA (L2 segment) | IC | T | SCI | T + SCI |
| dorsal horns | 1.38 ± 0.91 | 0.44 ± 0.72 | 0.63 ± 1.21 | 0.60 ± 0.69 |
| * | ns | ns | ||
| intermediate zone | 5.62 ± 1.92 | 4.37 ± 0.91 | 4.00 ± 2.00 | 5.50 ± 1.43 |
| ns | ns | ns | ||
| ventral horns | 11.13 ± 1.35 | 8.77 ± 1.39 | 9.09 ± 1.57 | 9.70 ± 1.83 |
| ** | ** | ns | ||
| Glyt2 (L2 segment) | IC | T | SCI | T + SCI |
| dorsal horns | 0.87 ± 1.25 | 0.55 ± 0.72 | 0.33 ± 0.71 | 1.13 ± 1.64 |
| ns | ns | ns | ||
| intermediate zone | 4.25 ± 1.66 | 3.77 ± 7.09 | 1.56 ± 1.51 | 5.13 ± 1.46 |
| ns | ** | *** | ||
| ventral horns | 10.67 ± 3.12 | 8.40 ± 2.45 | 7.44 ± 1.23 | 10.00 ± 1.93 |
| ns | * | ** |
| Targets | Forward Primer | Reverse Primer |
|---|---|---|
| CNP-ase | CCTCAGAGCCACCACACATC | CCTTCCTTGGGGCTACAGTG |
| PLP1 | TTGGCGACTACAAGACCACC | AATGACACACCCGCTCCAAA |
| NF-l | AAGGCTAAGACCCTGGAGATCGAAG | GGGATAGTTGGGAATGGGGCTCAA |
| Gap-43 | AGGAAAGGAGAGAAGGCAGG | GCAGGAGAGACAGGGTTCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiss Bimbova, K.; Bacova, M.; Kisucka, A.; Gálik, J.; Ileninova, M.; Kuruc, T.; Magurova, M.; Lukacova, N. Impact of Endurance Training on Regeneration of Axons, Glial Cells, and Inhibitory Neurons after Spinal Cord Injury: A Link between Functional Outcome and Regeneration Potential within the Lesion Site and in Adjacent Spinal Cord Tissue. Int. J. Mol. Sci. 2023, 24, 8616. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24108616
Kiss Bimbova K, Bacova M, Kisucka A, Gálik J, Ileninova M, Kuruc T, Magurova M, Lukacova N. Impact of Endurance Training on Regeneration of Axons, Glial Cells, and Inhibitory Neurons after Spinal Cord Injury: A Link between Functional Outcome and Regeneration Potential within the Lesion Site and in Adjacent Spinal Cord Tissue. International Journal of Molecular Sciences. 2023; 24(10):8616. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24108616
Chicago/Turabian StyleKiss Bimbova, Katarina, Maria Bacova, Alexandra Kisucka, Ján Gálik, Maria Ileninova, Tomas Kuruc, Martina Magurova, and Nadezda Lukacova. 2023. "Impact of Endurance Training on Regeneration of Axons, Glial Cells, and Inhibitory Neurons after Spinal Cord Injury: A Link between Functional Outcome and Regeneration Potential within the Lesion Site and in Adjacent Spinal Cord Tissue" International Journal of Molecular Sciences 24, no. 10: 8616. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24108616