
Polynucleotides Suppress Inflammation and Stimulate Matrix Synthesis in an In Vitro Cell-Based Osteoarthritis Model

 , , and
, , and
Abstract
:1. Introduction
2. Results
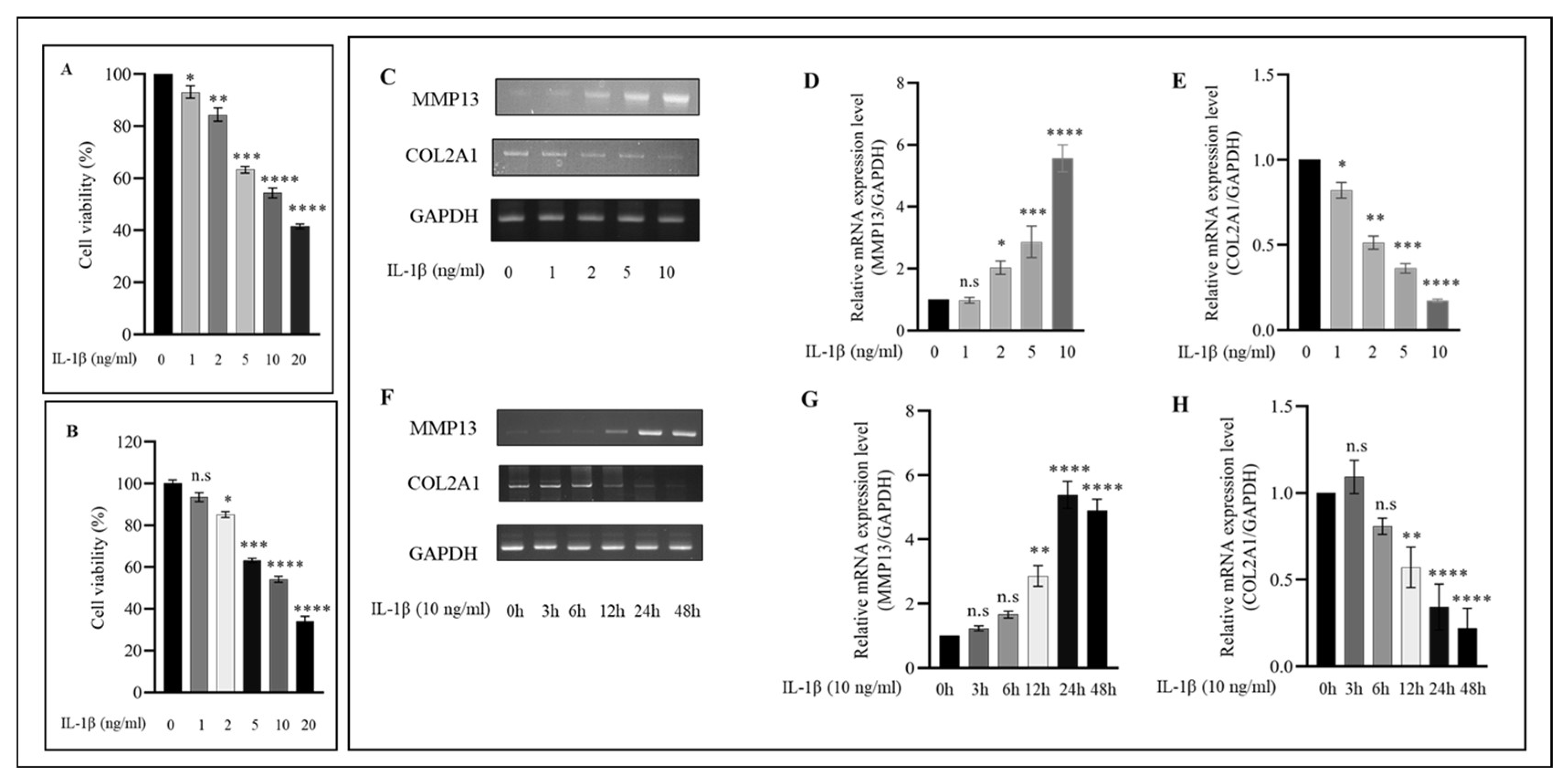
2.1. IL−1β Induces the Expression of Proinflammatory Markers in HC-a in a Concentration and Time-Dependent Manner
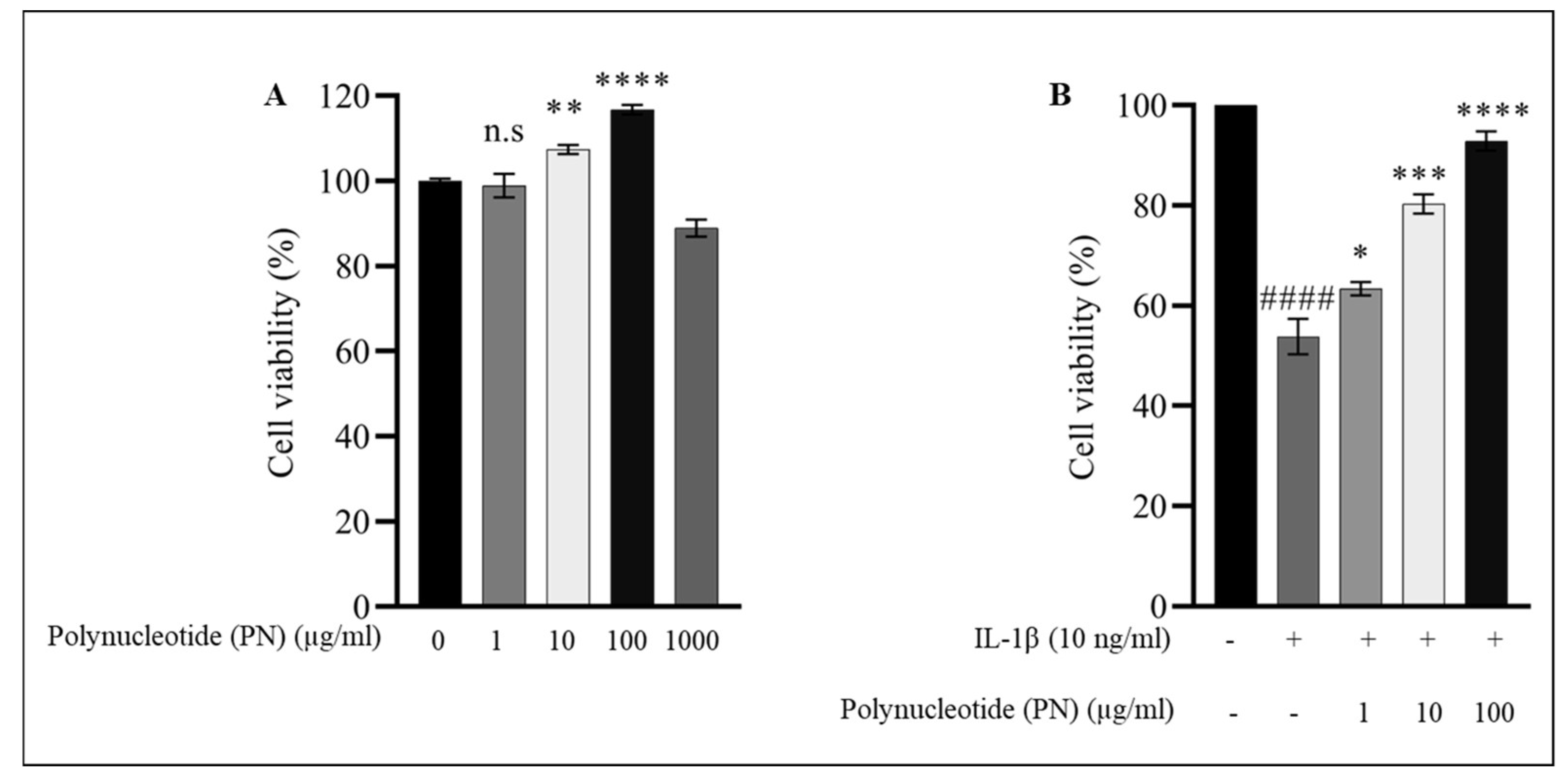
2.2. Effect of PN on HC-a Viability
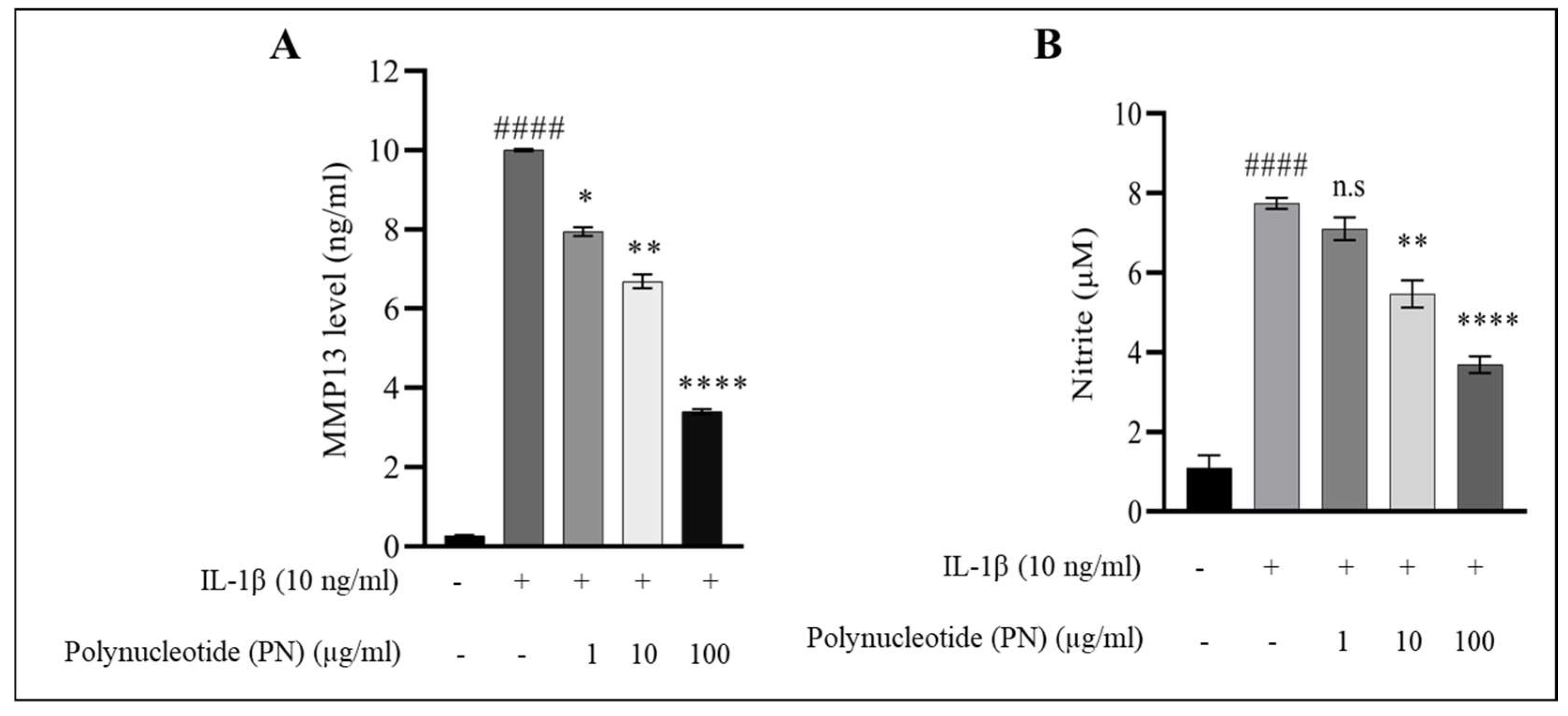
2.3. MMP13 and Nitric Oxide (NO) Assay
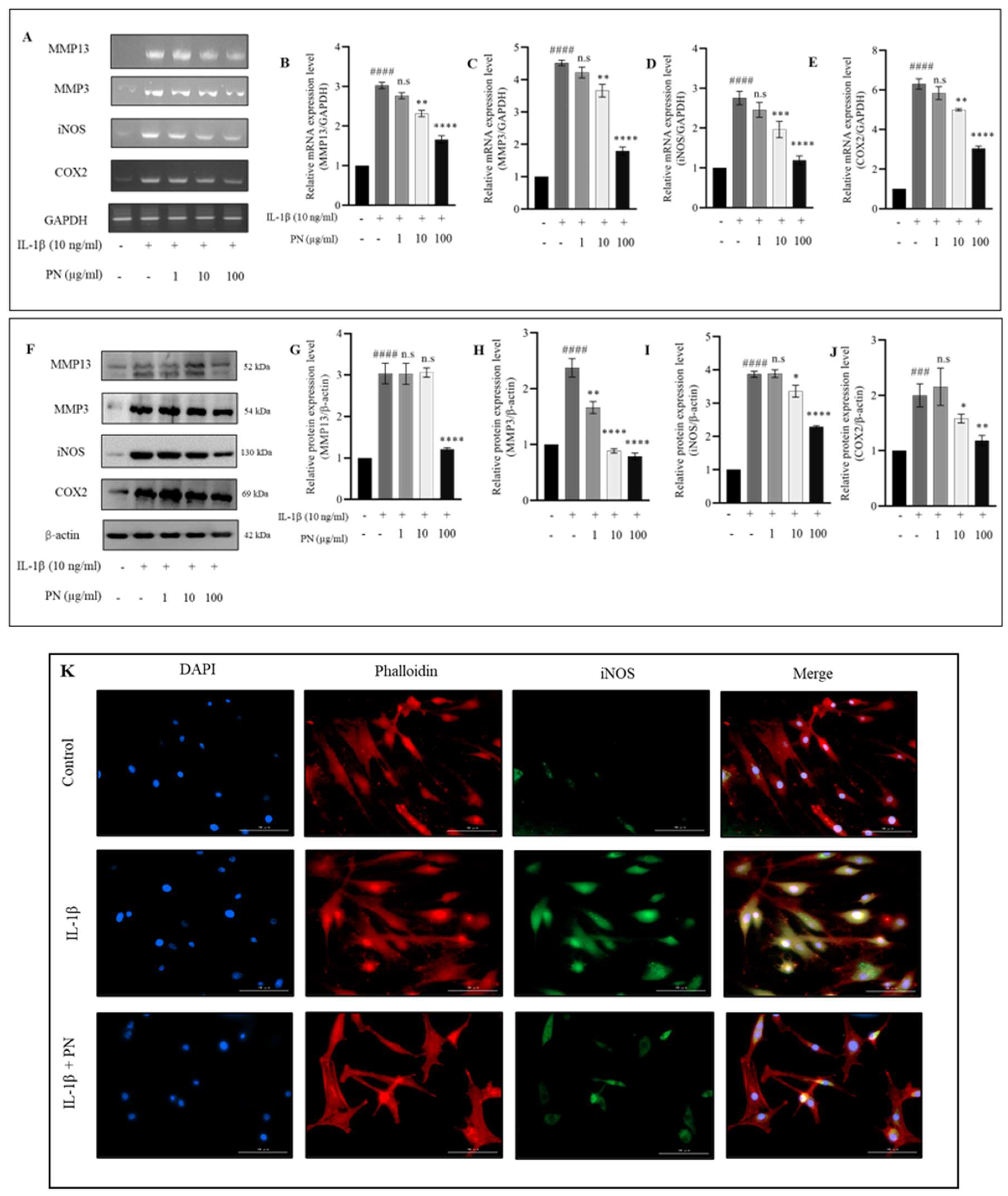
2.4. Effects of PN on Inflammation and Hypertrophy in IL−1β-Induced HC-a
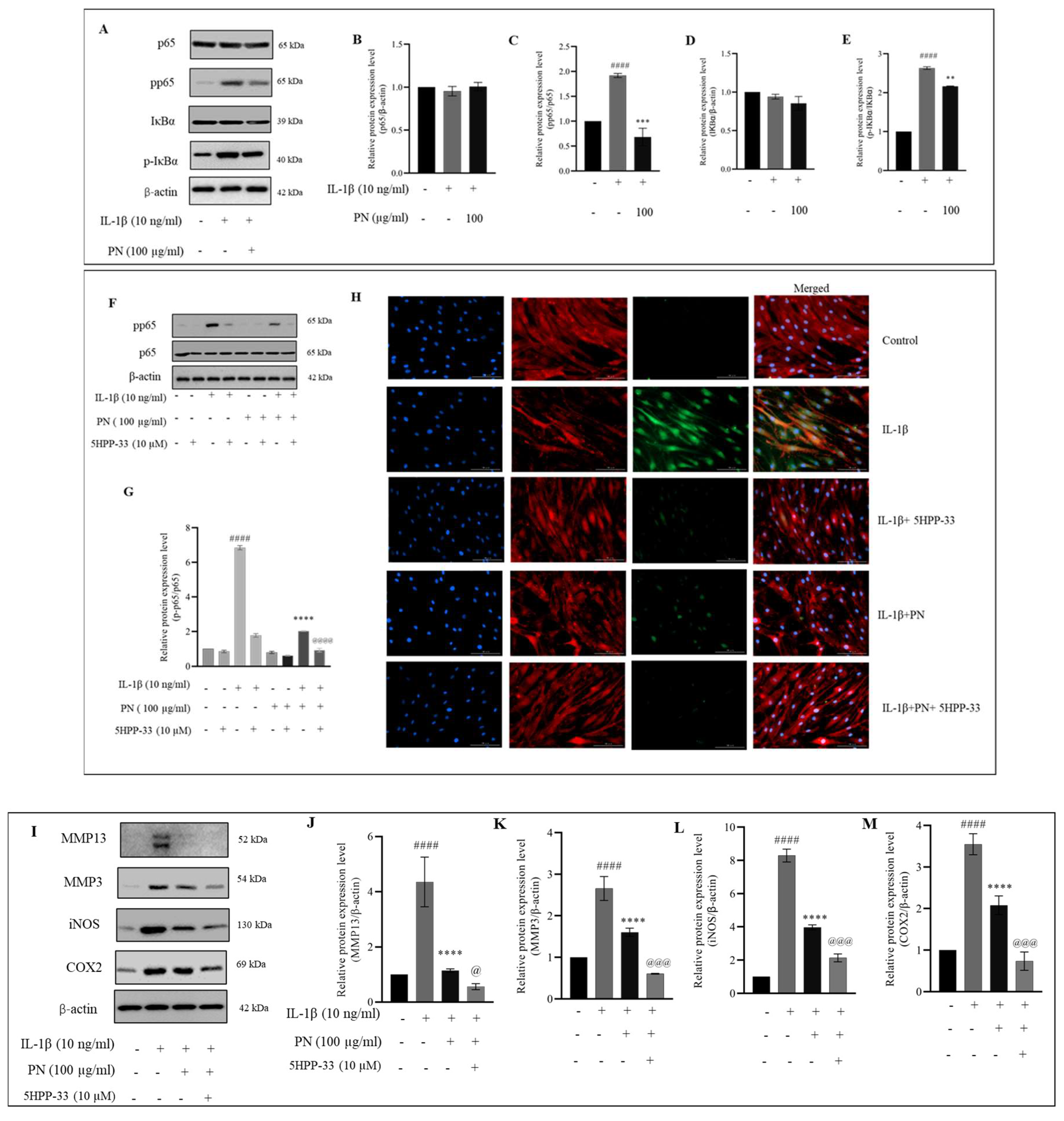
2.5. PN Relieves IL−1β-Induced Inflammation of HC-a through the NF-κB Signaling Pathway
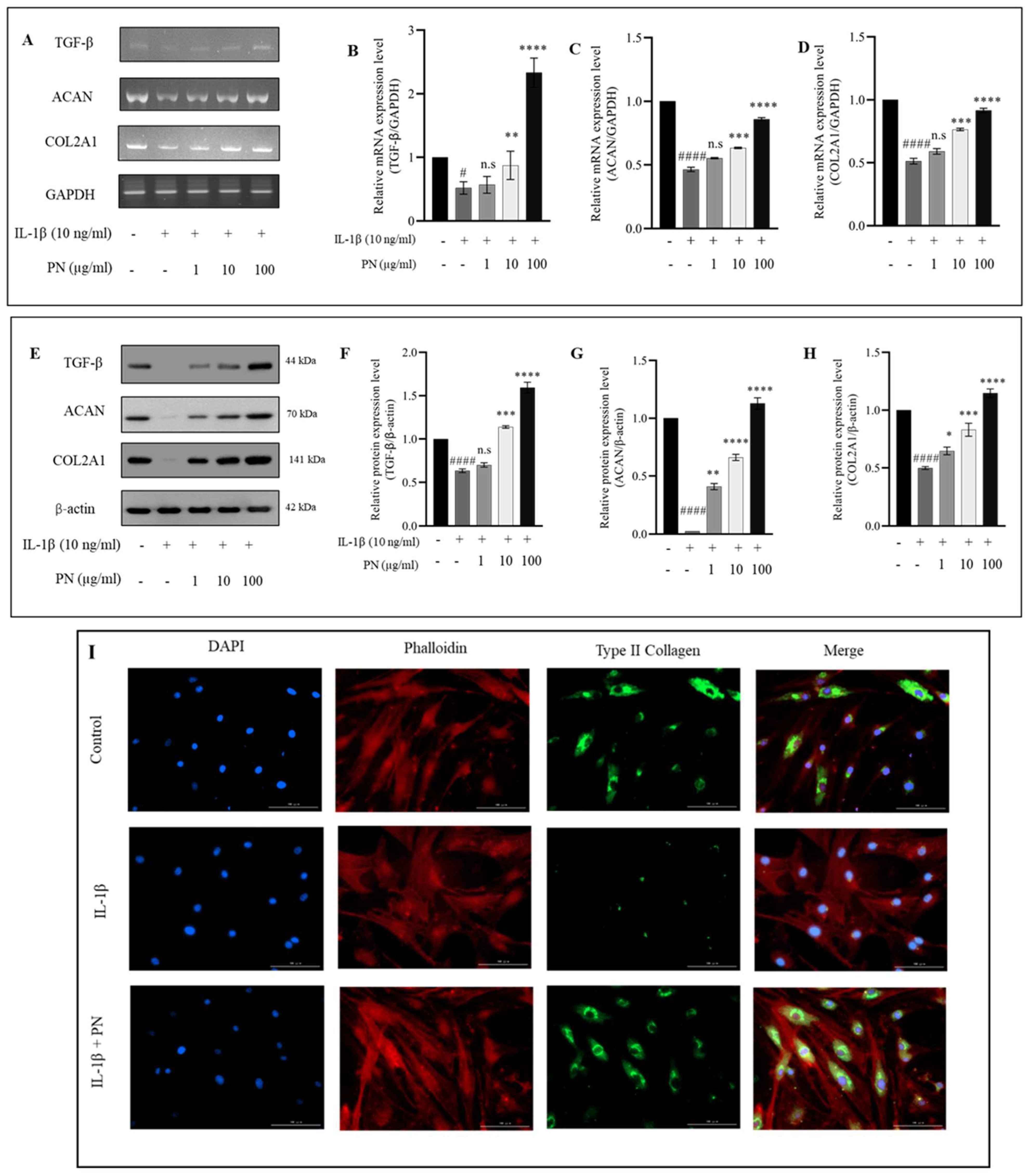
2.6. Effects of PN on ECM Synthesis in IL1β-Induced HC-a
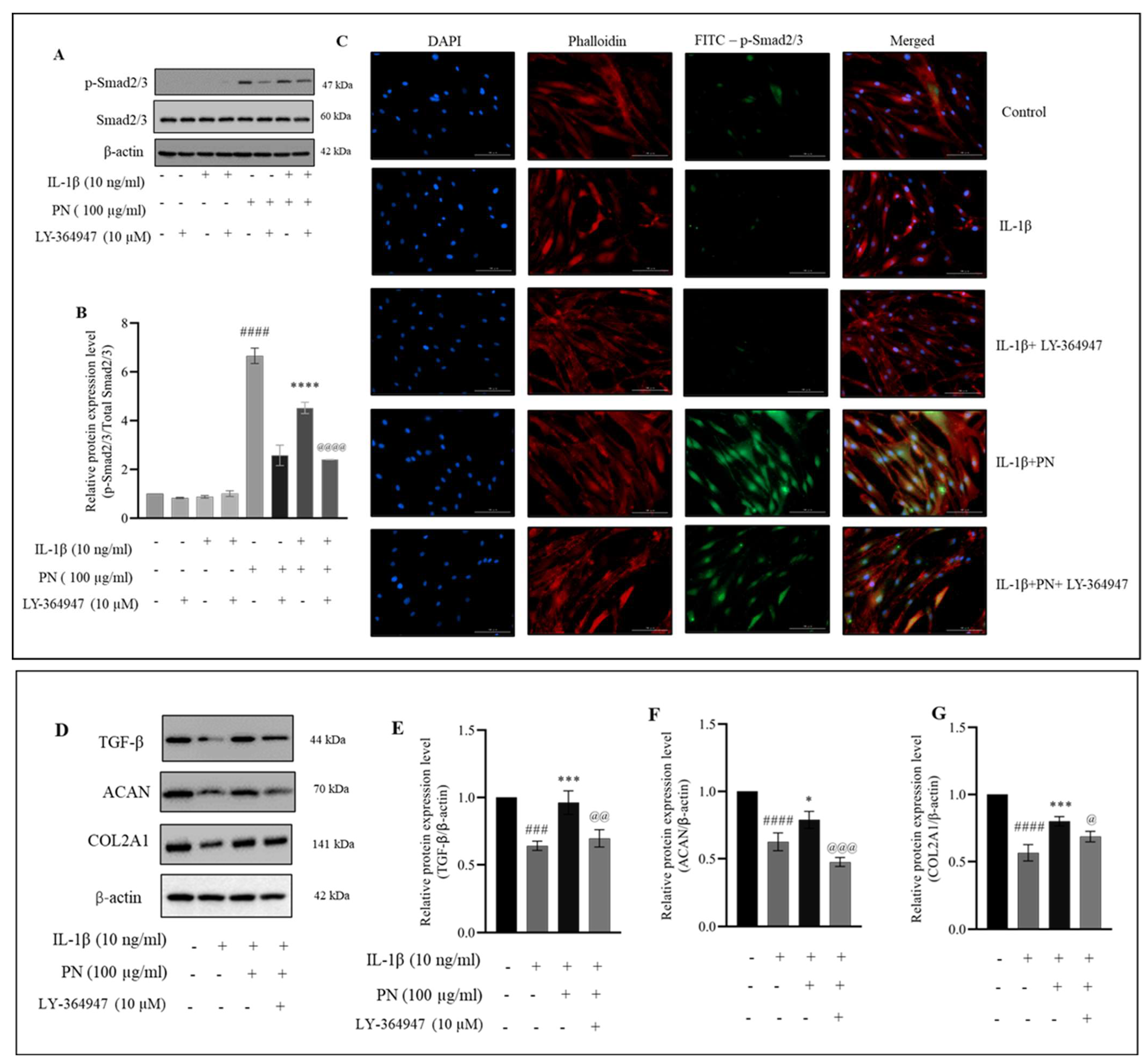
2.7. PN Increases the Phosphorylation of Smad2/3 in HC-a
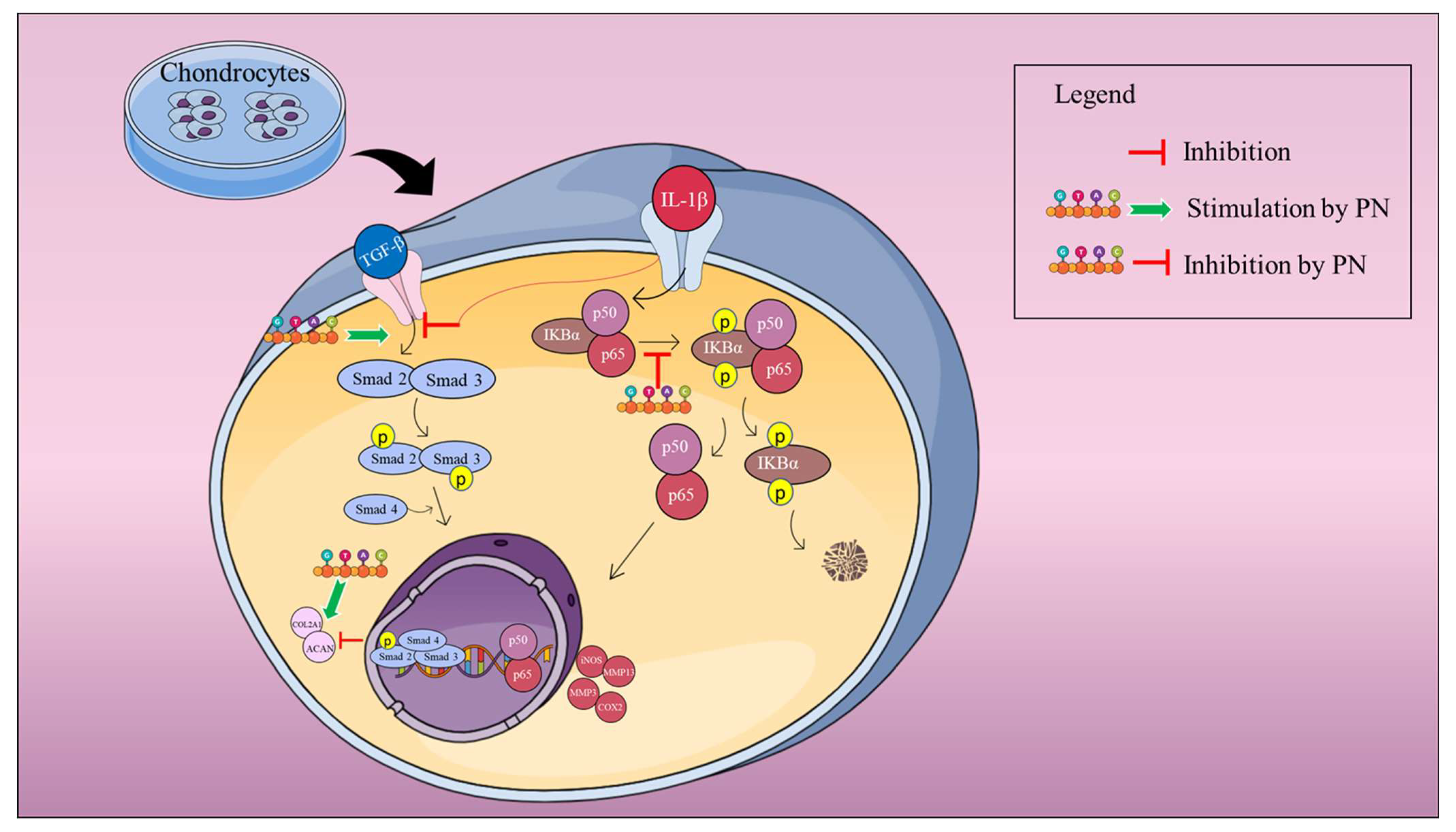
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Chondrocyte Induction In Vitro
4.4. Measurement of Cell Viability
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Nitric Oxide (NO) Assay
4.7. PCR
4.8. Whole-Cell Lysate Preparation and Western Blot Analysis
4.9. Inhibitor Treatment
4.10. Immunofluorescence
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Terkawi, M.A.; Ebata, T.; Yokota, S.; Takahashi, D.; Endo, T.; Matsumae, G.; Shimizu, T.; Kadoya, K.; Iwasaki, N. Low-Grade Inflammation in the Pathogenesis of Osteoarthritis: Cellular and Molecular Mechanisms and Strategies for Future Therapeutic Intervention. Biomedicines 2022, 10, 1109. [Google Scholar] [CrossRef] [PubMed]
- Cieza, A.; Causey, K.; Kamenov, K.; Hanson, S.W.; Chatterji, S.; Vos, T. Global estimates of the need for rehabilitation based on the Global Burden of Disease study 2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 2006–2017. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.-J. Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef]
- Selig, M.; Azizi, S.; Walz, K.; Lauer, J.C.; Rolauffs, B.; Hart, M.L. Cell morphology as a biological fingerprint of chondrocyte phenotype in control and inflammatory conditions. Front. Immunol. 2023, 14, 1102912. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.; Nam, J. The Role of Changes in Extracellular Matrix of Cartilage in the Presence of Inflammation on the Pathology of Osteoarthritis. BioMed. Res. Int. 2013, 2013, 284873. [Google Scholar] [CrossRef] [Green Version]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Attur, M.; Belitskaya-Lévy, I.; Oh, C.; Krasnokutsky, S.; Greenberg, J.; Samuels, J.; Smiles, S.; Lee, S.; Patel, J.; Al-Mussawir, H.; et al. Increased interleukin-1β gene expression in peripheral blood leukocytes is associated with increased pain and predicts risk for progression of symptomatic knee osteoarthritis. Arthritis Rheum. 2011, 63, 1908–1917. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Söder, S.; Oehler, S.; Fundel, K.; Aigner, T. Activation of Interleukin-1 Signaling Cascades in Normal and Osteoarthritic Articular Cartilage. Am. J. Pathol. 2007, 171, 938–946. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhong, L.; Hendriks, J.; Post, J.; Karperien, M. Different response of human chondrocytes from healthy looking areas and damaged regions to IL1β stimulation under different oxygen tension. J. Orthop. Res. 2018, 37, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Qu, R.; Chen, X.; Wang, W.; Qiu, C.; Ban, M.; Guo, L.; Vasilev, K.; Chen, J.; Li, W.; Zhao, Y. Ghrelin protects against osteoarthritis through interplay with Akt and NF-κB signaling pathways. FASEB J. 2018, 32, 1044–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, X.; Chen, X.; Cheng, S.; Shen, Y.; Peng, L.; Xu, H.Z. Piperine inhibits IL-β induced expression of inflammatory mediators in human osteoarthritis chondrocyte. Int. Immunopharmacol. 2013, 17, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Kuppa, S.S.; Kim, H.K.; Kang, J.Y.; Lee, S.C.; Seon, J.K. Role of Mesenchymal Stem Cells and Their Paracrine Mediators in Macrophage Polarization: An Approach to Reduce Inflammation in Osteoarthritis. Int. J. Mol. Sci. 2022, 23, 13016. [Google Scholar] [CrossRef] [PubMed]
- Olivotto, E.; Otero, M.; Marcu, K.B.; Goldring, M.B. Pathophysiology of osteoarthritis: Canonical NF-κB/IKKβ-dependent and kinase-independent effects of IKKα in cartilage degradation and chondrocyte differentiation. RMD Open 2015, 1, e000061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.-C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-B Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thielen, N.; Neefjes, M.; Wiegertjes, R.; Akker, G.v.D.; Vitters, E.; van Beuningen, H.; Davidson, E.B.; Koenders, M.; van Lent, P.; van de Loo, F.; et al. Osteoarthritis-Related Inflammation Blocks TGF-β’s Protective Effect on Chondrocyte Hypertrophy via (de)Phosphorylation of the SMAD2/3 Linker Region. Int. J. Mol. Sci. 2021, 22, 8124. [Google Scholar] [CrossRef]
- Madej, W.; Buma, P.; van der Kraan, P. Inflammatory conditions partly impair the mechanically mediated activation of Smad2/3 signaling in articular cartilage. Thromb. Haemost. 2016, 18, 146. [Google Scholar] [CrossRef] [Green Version]
- Burch, M.L.; Zheng, W.; Little, P.J. Smad linker region phosphorylation in the regulation of extracellular matrix synthesis. Cell. Mol. Life Sci. 2011, 68, 97–107. [Google Scholar] [CrossRef]
- Latourte, A.; Kloppenburg, M.; Richette, P. Emerging pharmaceutical therapies for osteoarthritis. Nat. Rev. Rheumatol. 2020, 16, 673–688. [Google Scholar] [CrossRef]
- Chen, T.; Weng, W.; Liu, Y.; Aspera-Werz, R.H.; Nüssler, A.K.; Xu, J. Update on Novel Non-Operative Treatment for Osteoarthritis: Current Status and Future Trends. Front. Pharmacol. 2021, 12, 755230. [Google Scholar] [CrossRef]
- Pontes-Quero, G.M.; García-Fernández, L.; Aguilar, M.R.; Román, J.S.; Cano, J.P.; Vázquez-Lasa, B. Active viscosupplements for osteoarthritis treatment. Semin. Arthritis Rheum. 2019, 49, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Abate, M.; Pulcini, D.; Di Iorio, A.; Schiavone, C. Viscosupplementation with Intra-Articular Hyaluronic Acid for Treatment of Osteoarthritis in the Elderly. Curr. Pharm. Des. 2010, 16, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, M.; De Luca, C.; Prussia, G.; Raichi, M. PN-HPT ® (Polynucleotides Highly Purified Technology) in facial middle third rejuvenation. Exploring the potential. J. Cosmet. Dermatol. 2022, 21, 615–624. [Google Scholar] [CrossRef]
- Vanelli, R.; Costa, P.; Rossi, S.M.P.; Benazzo, F. Efficacy of intra-articular polynucleotides in the treatment of knee osteoarthritis: A randomized, double-blind clinical trial. Knee Surg. Sports Traumatol. Arthrosc. 2010, 18, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Chang, M.J.; Shin, C.Y.; Chang, C.B.; Kang, S.-B. A randomized controlled trial for comparing efficacy and safety between intraarticular polynucleotide and hyaluronic acid for knee osteoarthritis treatment. Sci. Rep. 2023, 13, 9419. [Google Scholar] [CrossRef] [PubMed]
- Rathbone, M.; Christjanson, L.; Deforge, S.; Deluca, B.; Gysbers, J.; Hindley, S.; Jovetich, M.; Middlemiss, P.; Takhal, S. Extracellular purine nucleosides stimulate cell division and morphogenesis: Pathological and physiological implications. Med. Hypotheses 1992, 37, 232–240. [Google Scholar] [CrossRef]
- Hwang, J.-T.; Lee, S.-S.; Han, S.H.; Sherchan, B.; Panakkal, J.J. Polydeoxyribonucleotide and Polynucleotide Improve Tendon Healing and Decrease Fatty Degeneration in a Rat Cuff Repair Model. Tissue Eng. Regen. Med. 2021, 18, 1009–1020. [Google Scholar] [CrossRef]
- Jang, J.Y.; Kim, J.H.; Kim, M.W.; Kim, S.H.; Yong, S.Y. Study of the Efficacy of Artificial Intelligence Algorithm-Based Analysis of the Functional and Anatomical Improvement in Polynucleotide Treatment in Knee Osteoarthritis Patients: A Prospective Case Series. J. Clin. Med. 2022, 11, 2845. [Google Scholar] [CrossRef]
- Giarratana, L.S.; Marelli, B.M.; Crapanzano, C.; De Martinis, S.E.; Gala, L.; Ferraro, M.; Marelli, N.; Albisetti, W. A randomized double-blind clinical trial on the treatment of knee osteoarthritis: The efficacy of polynucleotides compared to standard hyaluronian viscosupplementation. Knee 2014, 21, 661–668. [Google Scholar] [CrossRef]
- Cavallini, M.; Bartoletti, E.; Maioli, L.; Massirone, A.; Palmieri, I.P.; Papagni, M.; Priori, M.; Trocchi, G. Consensus report on the use of PN-HPT™ (polynucleotides highly purified technology) in aesthetic medicine. J. Cosmet. Dermatol. 2021, 20, 922–928. [Google Scholar] [CrossRef]
- Moon, J.Y.; Kim, J.; Lee, J.Y.; Ko, Y.; Park, H.J.; Jeon, Y.H. Comparison of Polynucleotide, Sodium Hyaluronate, and Crosslinked Sodium Hyaluronate for the Management of Painful Knee Osteoarthritis: A Multi-Center, Randomized, Double-Blind, Parallel-Group Study. Pain Med. 2022, 24, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.; Thoma, L.; Golightly, Y. Epidemiology of osteoarthritis. Osteoarthr. Cartil. 2022, 30, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.-P. Osteoarthritis. Nat. Rev. Dis. Prim. 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magni, A.; Agostoni, P.; Bonezzi, C.; Massazza, G.; Menè, P.; Savarino, V.; Fornasari, D. Management of Osteoarthritis: Expert Opinion on NSAIDs. Pain Ther. 2021, 10, 783–808. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.V.; Jüni, P.; Saadat, P.; Xing, D.; Yao, L.; Bobos, P.; Agarwal, A.; A Hincapié, C.; da Costa, B.R. Viscosupplementation for knee osteoarthritis: Systematic review and meta-analysis. BMJ 2022, 378, e069722. [Google Scholar] [CrossRef]
- Park, J.; Park, H.J.; Rho, M.C.; Joo, J. Viscosupplementation in the Therapy for Osteoarthritic Knee. Appl. Sci. 2021, 11, 11621. [Google Scholar] [CrossRef]
- Park, K.Y.; Seok, J.; Rho, N.K.; Kim, B.J.; Kim, M.N. Long-chain polynucleotide filler for skin rejuvenation: Efficacy and complications in five patients. Dermatol. Ther. 2016, 29, 37–40. [Google Scholar] [CrossRef]
- Baek, A.; Kim, M.; Kim, S.H.; Cho, S.-R.; Kim, H.J. Anti-inflammatory Effect of DNA Polymeric Molecules in a Cell Model of Osteoarthritis. Inflammation 2018, 41, 677–688. [Google Scholar] [CrossRef]
- Baek, A.; Kim, Y.; Lee, J.W.; Lee, S.C.; Cho, S.-R. Effect of Polydeoxyribonucleotide on Angiogenesis and Wound Healing in an In Vitro Model of Osteoarthritis. Cell Transplant. 2018, 27, 1623–1633. [Google Scholar] [CrossRef] [Green Version]
- Mun, J.-U.; Cho, H.R.; Choi, Y.S.; Kim, Y.U. Effect of multiple intra-articular injections of polynucleotides on treatment of intractable knee osteoarthritis. Medicine 2017, 96, e9127. [Google Scholar] [CrossRef]
- Saggini, R.; Di Stefano, A.; Cavezza, T.; Saladino, G.; Bellomo, R.G. Intrarticular treatment of osteoartropaty knee with polynucleotides: A pilot study with medium-term follow-up. J. Biol. Regul. Homeost. Agents 2013, 27, 543–549. [Google Scholar] [PubMed]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.-P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, S.; Su, H.; Cheng, J. Isoliquiritigenin Inhibits IL-1β-Induced Production of Matrix Metalloproteinase in Articular Chondrocytes. Mol. Ther. Methods Clin. Dev. 2018, 9, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a Promising Target for the Treatment of Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 1742. [Google Scholar] [CrossRef]
- Xu, F.; Zhao, L.-J.; Liao, T.; Li, Z.-C.; Wang, L.-L.; Lin, P.-Y.; Jiang, R.; Wei, Q.-J. Ononin ameliorates inflammation and cartilage degradation in rat chondrocytes with IL-1β-induced osteoarthritis by downregulating the MAPK and NF-κB pathways. BMC Complement. Med. Ther. 2022, 22, 25. [Google Scholar] [CrossRef]
- Van der Kraan, P.M.; van den Berg, W.B. Chondrocyte hypertrophy and osteoarthritis: Role in initiation and progression of cartilage degeneration? Osteoarthr. Cartil. 2012, 20, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Oeckinghaus, A.; Ghosh, S. The NF- B Family of Transcription Factors and Its Regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Liao, S.; Zhou, K.; Li, D.; Xie, X.; Jun, F.; Wang, J. Schisantherin A suppresses interleukin-1β-induced inflammation in human chondrocytes via inhibition of NF-κB and MAPKs activation. Eur. J. Pharmacol. 2016, 780, 65–70. [Google Scholar] [CrossRef]
- Vuolteenaho, K.; Moilanen, T.; Knowles, R.G.; Moilanen, E. The role of nitric oxide in osteoarthritis. Scand. J. Rheumatol. 2007, 36, 247–258. [Google Scholar] [CrossRef]
- Ulivi, V.; Giannoni, P.; Gentili, C.; Cancedda, R.; Descalzi, F. p38/NF-kB-dependent expression of COX-2 during differentiation and inflammatory response of chondrocytes. J. Cell. Biochem. 2008, 104, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- De-Blanco, E.J.C.; Pandit, B.; Hu, Z.; Shi, J.; Lewis, A.; Li, P.-K. Inhibitors of NF-κB derived from thalidomide. Bioorganic Med. Chem. Lett. 2007, 17, 6031–6035. [Google Scholar] [CrossRef] [PubMed]
- Keifer, J.A.; Guttridge, D.C.; Ashburner, B.P.; Baldwin, A.S., Jr. Inhibition of NF-κB Activity by Thalidomide through Suppression of IκB Kinase Activity. J. Biol. Chem. 2001, 276, 22382–22387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Xie, H.; Chen, Z.; Xu, S.; Wang, B.; Peng, Q.; Sha, K.; Xiao, W.; Liu, T.; Zhang, Y.; et al. Thalidomide ameliorates rosacea-like skin inflammation and suppresses NF-κB activation in keratinocytes. Biomed. Pharmacother. 2019, 116, 109011. [Google Scholar] [CrossRef]
- Amirshahrokhi, K. Thalidomide reduces glycerol-induced acute kidney injury by inhibition of NF-κB, NLRP3 inflammasome, COX-2 and inflammatory cytokines. Cytokine 2021, 144, 155574. [Google Scholar] [CrossRef]
- Picciolo, G.; Mannino, F.; Irrera, N.; Altavilla, D.; Minutoli, L.; Vaccaro, M.; Arcoraci, V.; Squadrito, V.; Picciolo, G.; Squadrito, F.; et al. PDRN, a natural bioactive compound, blunts inflammation and positively reprograms healing genes in an “in vitro” model of oral mucositis. Biomed. Pharmacother. 2021, 138, 111538. [Google Scholar] [CrossRef]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef]
- Vincent, T.L.; McClurg, O.; Troeberg, L. The Extracellular Matrix of Articular Cartilage Controls the Bioavailability of Pericellular Matrix-Bound Growth Factors to Drive Tissue Homeostasis and Repair. Int. J. Mol. Sci. 2022, 23, 6003. [Google Scholar] [CrossRef]
- Roughley, P.J.; Mort, J.S. The role of aggrecan in normal and osteoarthritic cartilage. J. Exp. Orthop. 2014, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Timur, U.T.; Caron, M.; Akker, G.v.D.; van der Windt, A.; Visser, J.; van Rhijn, L.; Weinans, H.; Welting, T.; Emans, P.; Jahr, H. Increased TGF-β and BMP Levels and Improved Chondrocyte-Specific Marker Expression In Vitro under Cartilage-Specific Physiological Osmolarity. Int. J. Mol. Sci. 2019, 20, 795. [Google Scholar] [CrossRef] [Green Version]
- van der Kraan, P.M.; Davidson, E.N.B.; Berg, W.B.v.D. A role for age-related changes in TGFβ signaling in aberrant chondrocyte differentiation and osteoarthritis. Thromb. Haemost. 2010, 12, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Kraan, P.M.; Blaney Davidson, E.N.; Blom, A.; van den Berg, W.B. TGF-beta signaling in chondrocyte terminal differentiation and osteoarthritis. Osteoarthr. Cartil. 2009, 17, 1539–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, C.; Wang, X.; Qiu, X.; Wu, Z.; Gao, B.; Liu, L.; Liang, G.; Zhou, H.; Yang, X.; Peng, Y.; et al. Collagen type II suppresses articular chondrocyte hypertrophy and osteoarthritis progression by promoting integrin β1−SMAD1 interaction. Bone Res. 2019, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagni, C.; Rocchi, M.; Mazzotta, A.; Del Piccolo, N.; Rani, N.; Govoni, M.; Vivarelli, L.; Veronesi, F.; Fini, M.; Dallari, D. Randomised, double-blind comparison of a fixed co-formulation of intra-articular polynucleotides and hyaluronic acid versus hyaluronic acid alone in the treatment of knee osteoarthritis: Two-year follow-up. BMC Musculoskelet. Disord. 2021, 22, 773. [Google Scholar] [CrossRef]
- Kwon, D.R.; Kim, K.L.; Moon, Y.S. Regeneration of Chronic Rotator Cuff Tear in a Rabbit Model: Synergetic Benefits of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells, Polydeoxyribonucleotides, and Microcurrent Therapy. BioMed Res. Int. 2022, 2022, 6496773. [Google Scholar] [CrossRef]
- Johnson, C.I.; Argyle, D.J.; Clements, D.N. In vitro models for the study of osteoarthritis. Veter. J. 2016, 209, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Samvelyan, H.J.; Hughes, D.; Stevens, C.; Staines, K.A. Models of Osteoarthritis: Relevance and New Insights. Calcif. Tissue Int. 2021, 109, 243–256. [Google Scholar] [CrossRef]
- Cancedda, R.; Cancedda, F.D.; Castagnola, P. Chondrocyte Differentiation. Int. Rev. Cytol. 1995, 159, 265–358. [Google Scholar]
- Gosset, M.; Berenbaum, F.; Thirion, S.; Jacques, C. Primary culture and phenotyping of murine chondrocytes. Nat. Protoc. 2008, 3, 1253–1260. [Google Scholar] [CrossRef]
- Bao, X.; Ren, T.; Huang, Y.; Sun, K.; Wang, S.; Liu, K.; Zheng, B.; Guo, W. Knockdown of long non-coding RNA HOTAIR increases miR-454-3p by targeting Stat3 and Atg12 to inhibit chondrosarcoma growth. Cell Death Dis. 2017, 8, e2605. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Li, H.; Wu, B.; Jiang, N.; Wang, B.; Wang, D.; Zhong, J.; Chen, Y.; Xu, X.; Lu, H. CircHIPK3 prevents chondrocyte apoptosis and cartilage degradation by sponging miR-30a-3p and promoting PON2. Cell Prolif. 2022, 55, e13285. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S.; Grisham, M.B. Methods to detect nitric oxide and its metabolites in biological samples. Free. Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name. | Forward Primer | Reverse Primer | Melting Temp |
|---|---|---|---|
| hCollagen II | TCTGCAACATGGAGACTGGC | GAAGCAGACCGGCCCTATGT | F 55.7 R 57.6 |
| hAggrecan | ACGAGTGGCAGCGGTGAAT | GCCCTTCTCCTGCCTCTTG | F 57.6 R 56 |
| hCOX2 | TTC AAATGAGATTGTGGGAAA | AGATCATCTCTGCCTGAGTATCTT | F 51.9 R 52.0 |
| hMMP3 | GGCAGTTTGCTCAGCCTATC | GTCACCTCCAATCCAAGGAA | F 53.8 R 53.4 |
| hMMP13 | GATGAAGACCCCAACCCTAAA | CTGGCCAAAATGATTTCGTTA | F 54.3 R 54.1 |
| hiNOS | AGCGGGATGACTTTCCAAGA | CTCCCGTCAGTTGGTAGGTT | F 59.02 R 59.03 |
| hGAPDH | ACCACAGTCCATGCCATCAC | TCCACCACCCTGTTGCTGTA | F 55 R 55.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuppa, S.S.; Kim, H.-K.; Kang, J.-Y.; Lee, S.-C.; Yang, H.-Y.; Sankaranarayanan, J.; Seon, J.-K. Polynucleotides Suppress Inflammation and Stimulate Matrix Synthesis in an In Vitro Cell-Based Osteoarthritis Model. Int. J. Mol. Sci. 2023, 24, 12282. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241512282
Kuppa SS, Kim H-K, Kang J-Y, Lee S-C, Yang H-Y, Sankaranarayanan J, Seon J-K. Polynucleotides Suppress Inflammation and Stimulate Matrix Synthesis in an In Vitro Cell-Based Osteoarthritis Model. International Journal of Molecular Sciences. 2023; 24(15):12282. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241512282
Chicago/Turabian StyleKuppa, Sree Samanvitha, Hyung-Keun Kim, Ju-Yeon Kang, Seok-Cheol Lee, Hong-Yeol Yang, Jaishree Sankaranarayanan, and Jong-Keun Seon. 2023. "Polynucleotides Suppress Inflammation and Stimulate Matrix Synthesis in an In Vitro Cell-Based Osteoarthritis Model" International Journal of Molecular Sciences 24, no. 15: 12282. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241512282