
Aqueous Extracts from Hemp Seeds as a New Weapon against Staphylococcus epidermidis Biofilms

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
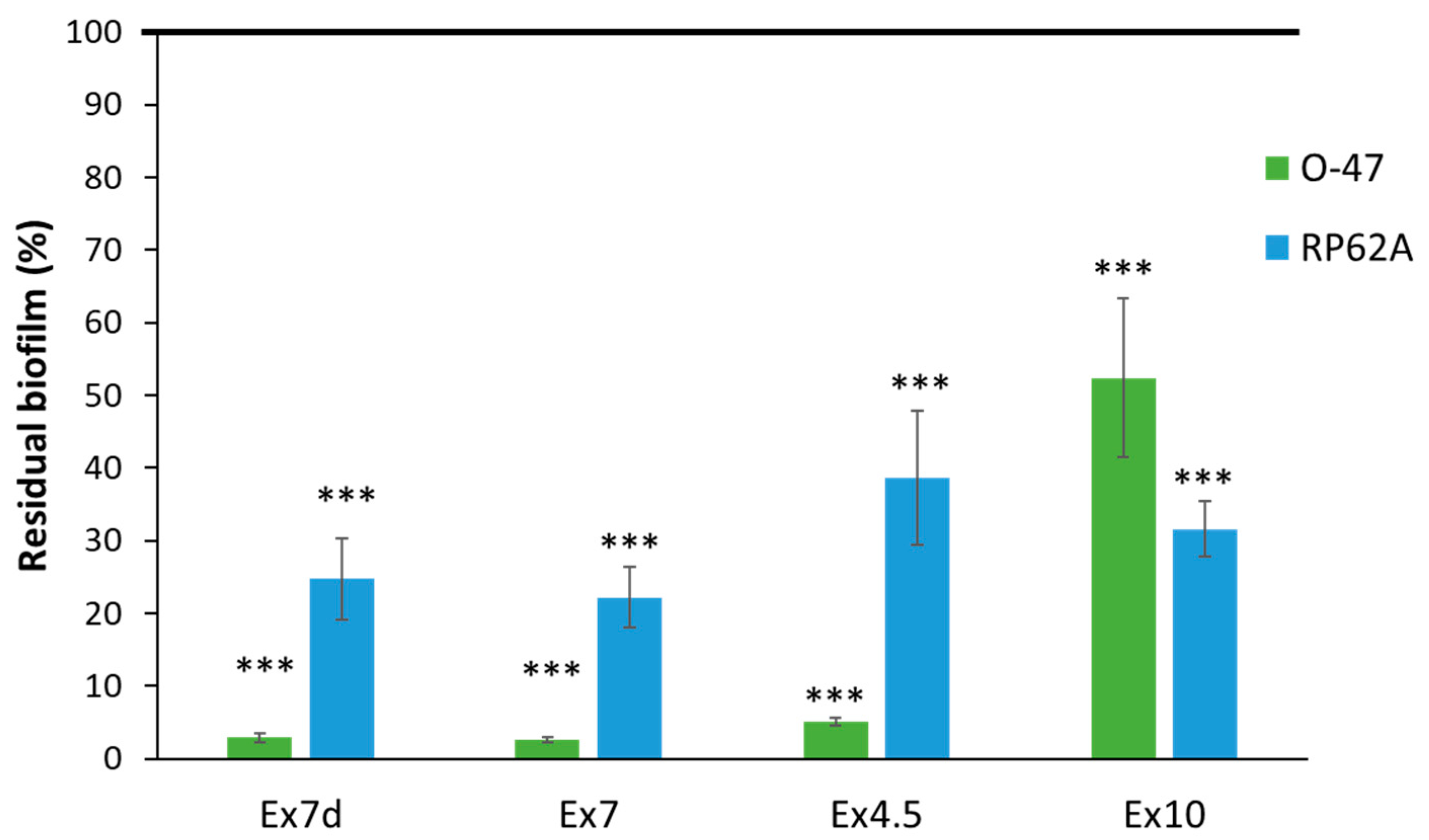
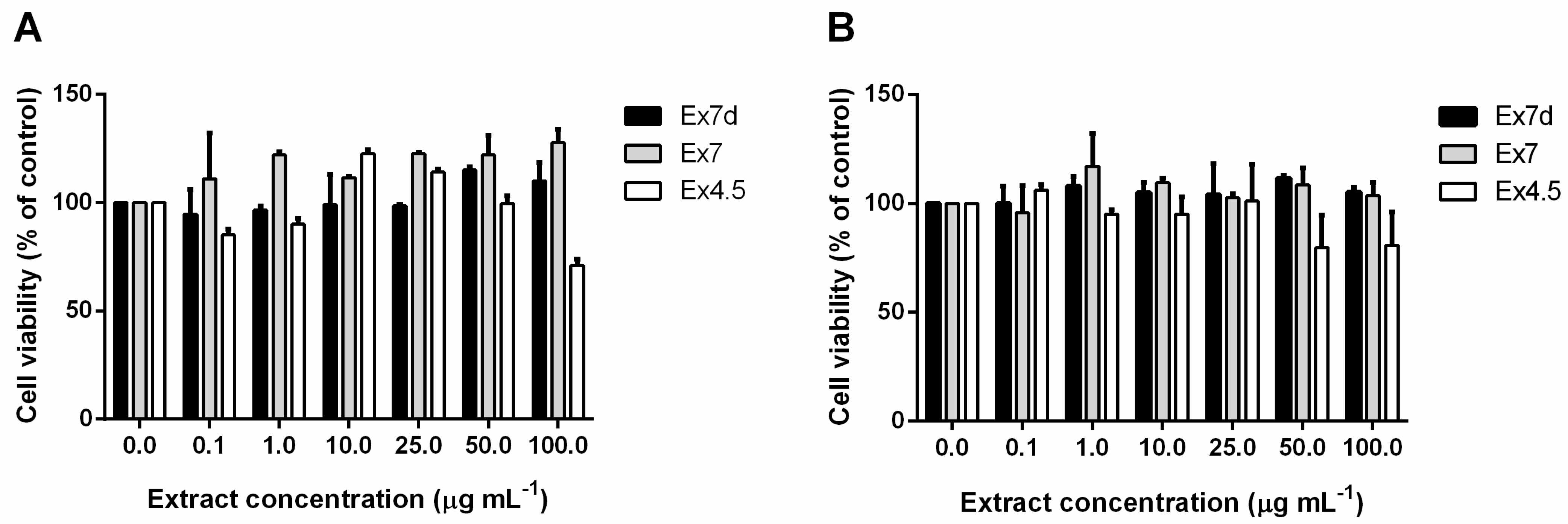
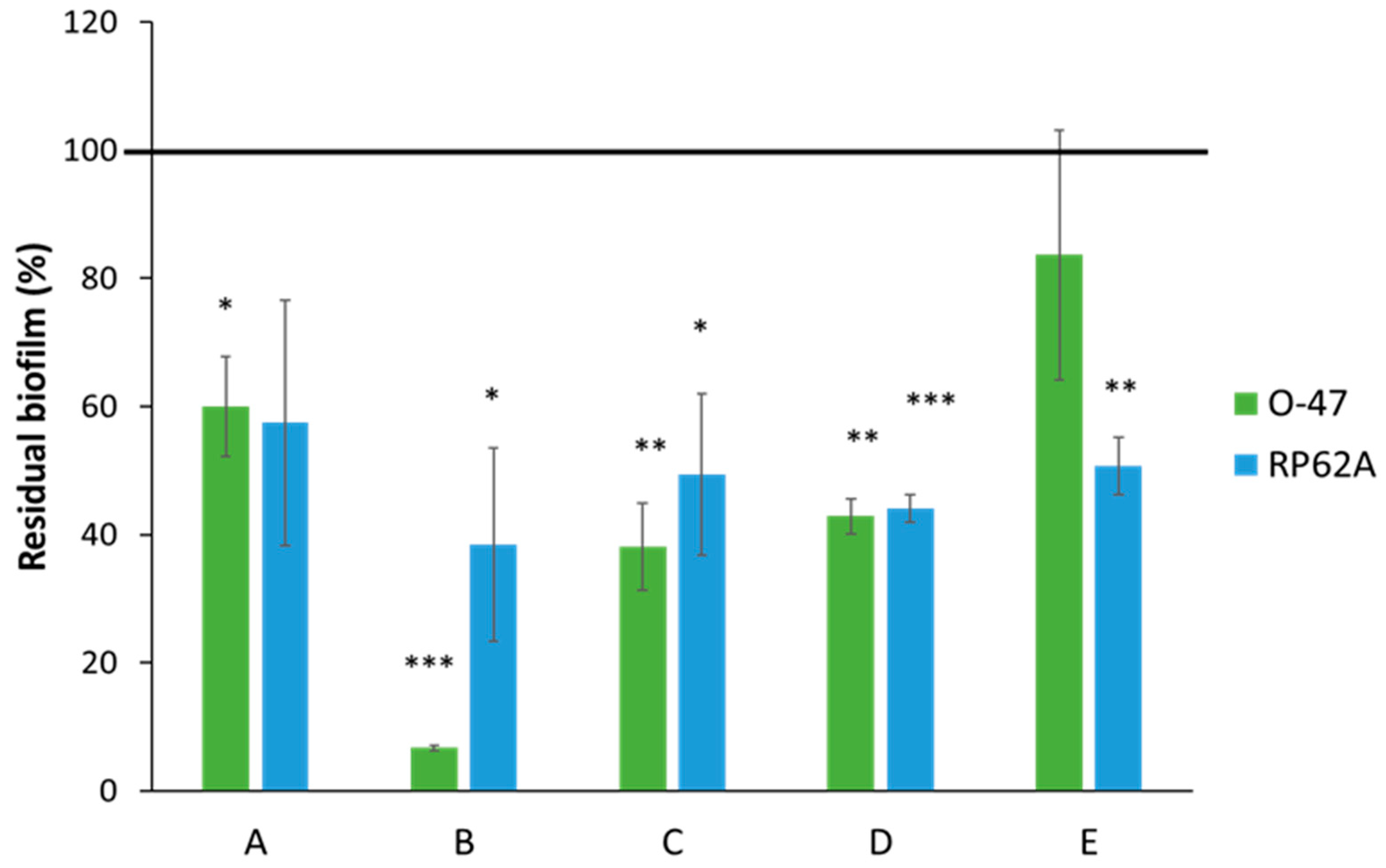
2. Results and Discussion
3. Materials and Methods
3.1. Extraction of Polar Components from Hemp Seeds
3.2. Protein Determination
3.3. GC-MS Analyses
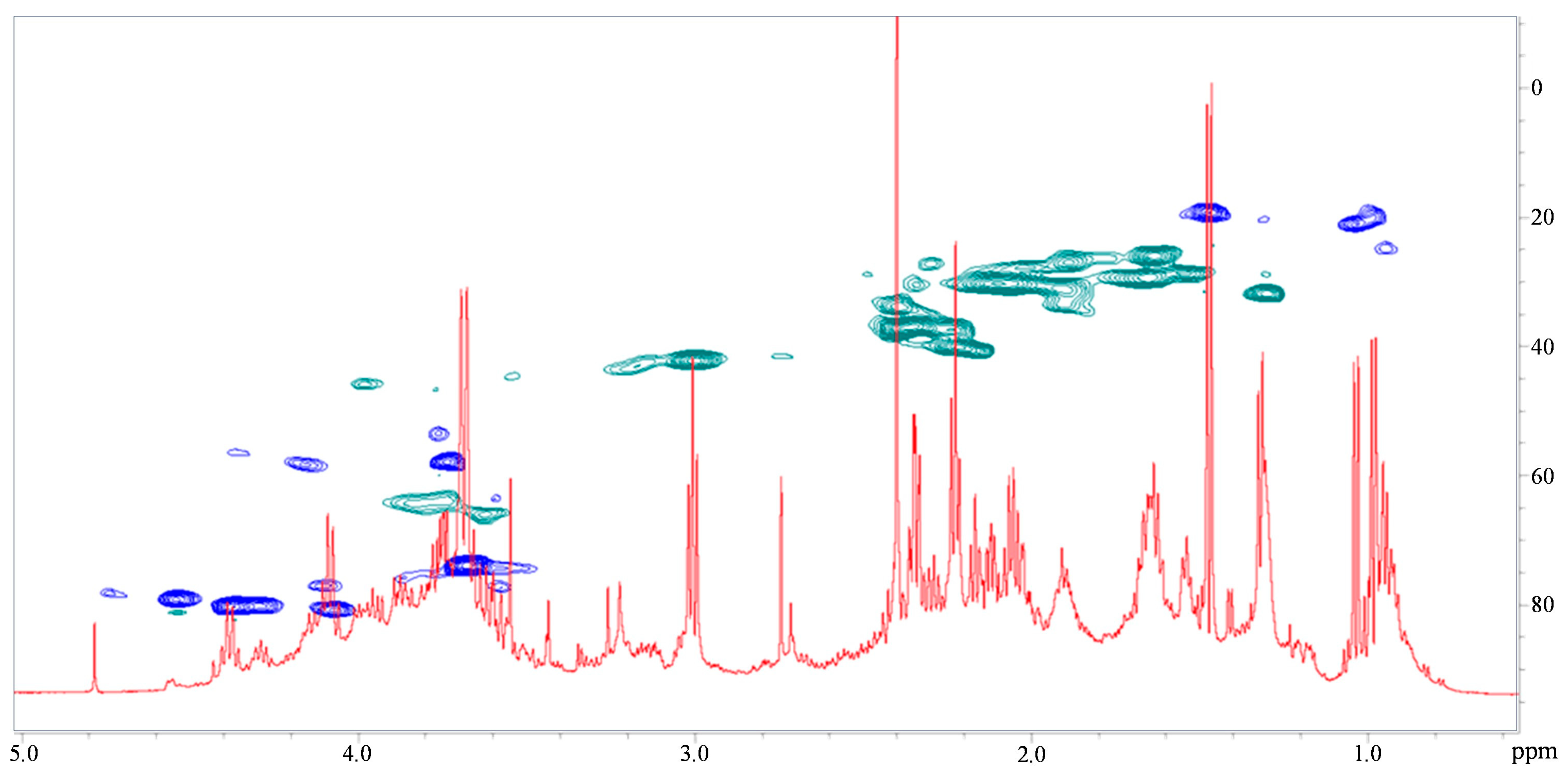
3.4. NMR Spectroscopy
3.5. Gel Filtration Chromatography
3.6. Bacterial Strains and Culture Conditions
3.7. Determination of Minimal Inhibitory Concentrations
3.8. Antibiofilm Assay
3.9. Cytotoxicity Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The Plant of the thousand and One Molecules. Front. Plant. Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Andres, M.; Cole, M.; Cowley, J.M.; Augustin, M.A. Industrial hemp seed: From the feld to value-added food ingredients. J. Cannabis Res. 2022, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Ranalli, P.; Venturi, G. Hemp as a raw material for industrial applications. Euphytica 2004, 140, 1–6. [Google Scholar] [CrossRef]
- Valizadehderakhshan, M.; Shahbazi, A.; Kazem-Rostami, M.; Todd, M.S.; Bhowmik, A.; Wang, L. Extraction of Cannabinoids from Cannabis sativa L. (Hemp)—Review. Agriculture 2021, 11, 384. [Google Scholar] [CrossRef]
- Kaur, S.; Nikita, S.; Arpita, R. Role of cannabinoids in various diseases: A review. Curr. Pharm. Biotechnol. 2022, 23, 1346–1358. [Google Scholar]
- Kogan, N.M.; Mechoulam, R. Cannabinoids in health and disease. Dialogues Clin. Neurosci. 2007, 9, 413–430. [Google Scholar] [CrossRef]
- Rodríguez Mesa, X.M.; Moreno Vergara, A.F.; Contreras Bolaños, L.A.; Guevara Moriones, N.; Mejía Piñeros, A.L.; Santander González, S.P. Therapeutic prospects of cannabinoids in the immunomodulation of prevalent autoimmune diseases. Cannabis Cannabinoid Res. 2021, 6, 196–210. [Google Scholar] [CrossRef]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial Cannabinoids from Cannabis sativa: A Structure−Activity Study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef]
- Nissen, L.; Zatta, A.; Stefanini, I.; Grandi, S.; Sgorbati, B.; Biavati, B.; Monti, A. Characterization and antimicrobial activity of essential oils of industrial hemp varieties (Cannabis sativa L.). Fitoterapia 2010, 81, 413–419. [Google Scholar] [CrossRef]
- Di Onofrio, V.; Gesuele, R.; Maione, A.; Liguori, G.; Liguori, R.; Guida, M.; Nigro, R.; Galdiero, E. Prevention of Pseudomonas aeruginosa Biofilm Formation on Soft Contact Lenses by Allium Sativum Fermented Extract (BGE) and Cannabinol Oil Extract (CBD). Antibiotics 2019, 8, 258. [Google Scholar] [CrossRef]
- Marini, E.; Magi, G.; Ferretti, G.; Bacchetti, T.; Giuliani, A.; Pugnaloni, A.; Rippo, M.R.; Facinelli, M. Attenuation of Listeria monocytogenes Virulence by Cannabis sativa L. Essential Oil. Front. Cell. Infect. Microbiol. 2018, 8, 293. [Google Scholar] [CrossRef] [PubMed]
- Krejčí, Z.; Horak, M.; Santavy, F. Hemp (Cannabis sativa)—An antibiotic drug. 3. Isolation and constitution of two acids from Cannabis sativa. Pharmazie 1959, 14, 349–355. [Google Scholar]
- Radoševič, A.; Kupinič, M.; Grlč, L. Antibiotic activity of various types of Cannabis resin. Nature 1962, 195, 1007–1009. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, M.S.; Hossain, M.S.; Ahmed, A.; Islam, M.Z.; Sarker, M.E.; Islam, M.R. Antimicrobial and antiviral (SARS-CoV-2) potential of cannabinoids and Cannabis sativa: A comprehensive review. Molecules 2021, 26, 7216. [Google Scholar] [CrossRef] [PubMed]
- Martinenghi, L.D.; Jønsson, R.; Lund, T.; Jenssen, H. Isolation, purification, and antimicrobial characterization of cannabidiolic acid and cannabidiol from Cannabis sativa L. Biomolecules 2020, 10, 900. [Google Scholar] [CrossRef] [PubMed]
- Fathordoobady, F.; Singh, A.; Kitts, D.D.; Singh, A.P. Hemp (Cannabis sativa L.) Extract: Anti-Microbial Properties, Methods of Extraction, and Potential Oral Delivery. Food Rev. Int. 2019, 35, 664–684. [Google Scholar] [CrossRef]
- Mirpoor, S.F.; Giosafatto, C.V.L.; Di Girolamo, R.; Famiglietti, M.; Porta, R. Hemp (Cannabis sativa) seed oilcake as a promising by-product for developing protein-based films: Effect of transglutaminase-induced crosslinking. Food Packag. Shelf Life 2022, 31, 100779. [Google Scholar] [CrossRef]
- Russo, R.; Reggiani, R. Evaluation of protein concentration, amino acid profile and antinutritional compounds in hemp seed meal from dioecious and monoecious varieties. Am. J. Plant Sci. 2015, 6, 14–22. [Google Scholar] [CrossRef]
- Wen, Z.-S.; Xue, R.; Du, M.; Tanga, Z.; Xiang, X.-W.; Zheng, B.; Qu, J.-L. Hemp seed polysaccharides protect intestinal epithelial cells from hydrogen peroxide-induced oxidative stress. Int. J. Biol. Macromol. 2019, 135, 203–211. [Google Scholar] [CrossRef]
- Yu, L.L.; Zhou, K.K.; Parry, J. Antioxidant properties of cold-pressed black caraway, carrot, cranberry, and hemp seed oils. Food Chem. 2005, 91, 723–729. [Google Scholar] [CrossRef]
- Zengin, G.; Menghini, L.; Di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A.; Angiolella, L.; et al. Chromatographic Analyses, In Vitro Biological Activities, and Cytotoxicity of Cannabis sativa L. Essential Oil: A Multidisciplinary Study. Molecules 2018, 23, 3266. [Google Scholar] [CrossRef]
- Ostapczuk, K.; Apori, S.O.; Estrada, G.; Tian, F. Hemp Growth Factors and Extraction Methods Effect on Antimicrobial Activity of Hemp Seed Oil: A Systematic Review. Separations 2021, 8, 183. [Google Scholar] [CrossRef]
- Aluko, R.E. Hemp seed (Cannabis sativa L.) proteins: Composition, structure, enzymatic modification, and functional or bioactive properties. In Sustainable Protein Sources; Academic Press: Cambridge, MA, USA, 2017; pp. 121–132. [Google Scholar]
- Chen, T.; He, J.; Zhang, J.; Li, X.; Zhang, H.; Hao, J.; Li, L. The isolation and identification of two compounds with predominant radical scavenging activity in hempseed (seed of Cannabis sativa L.). Food Chem. 2012, 134, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Malomo, S.A.; Aluko, R.E. A comparative study of the structural and functional properties of isolated hemp seed (Cannabis sativa L.) albumin and globulin fractions. Food Hydrocoll. 2015, 43, 743–752. [Google Scholar] [CrossRef]
- Julakanti, S.; Raja Charles, A.P.; Syed, R.; Bullock, F.; Wu, Y. Hempseed polysaccharide (Cannabis sativa L.): Physicochemical characterization and comparison with flaxseed polysaccharide. Food Hydrocoll. 2023, 143, 108900. [Google Scholar] [CrossRef]
- De Blanco, E.J.C.; Addo, E.M.; Rakotondraibe, H.L.; Soejarto, D.D.; Kinghorn, A.D. Strategies for the discovery of potential anticancer agents from plants collected from Southeast Asian tropical rainforests as a case study. Nat. Prod. Rep. 2023, 40, 1181–1197. [Google Scholar] [CrossRef]
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern Approaches in the Discovery and Development of Plant-Based Natural Products and Their Analogues as Potential Therapeutic Agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef]
- Caesar, L.K.; Montaser, R.; Keller, N.P.; Kelleher, N.L. Metabolomics and genomics in natural products research: Complementary tools for targeting new chemical entities. Nat. Prod. Rep. 2021, 38, 2041–2065. [Google Scholar] [CrossRef]
- Edison, A.S.; Clendinen, C.S.; Ajredini, R.; Beecher, C.; Ponce, F.V.; Stupp, G.S. Metabolomics and natural-products strategies to study chemical ecology in nematodes. Integr. Comp. Biol. 2015, 55, 478–485. [Google Scholar] [CrossRef]
- Zhang, L.; Jin, M.; Shi, X.; Jin, L.; Hou, X.; Yu, Y.; Liu, B.; Cao, J.; Quan, C. Macrolactin Metabolite Production by Bacillus sp. ZJ318 Isolated from Marine Sediment. Appl. Biochem. Biotechnol. 2022, 194, 2581–2593. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, S.L.; Kim, J.H.; Deng, H.Y.; Yun, B.S.; Lee, D.S. Triterpene acid (3-O-p-coumaroyltormentic acid) isolated from aronia extracts inhibits breast cancer stem cell formation through downregulation of c-Myc protein. Int. J. Mol. Sci. 2018, 19, 2528. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, K.; Kizhakkekalam, V.K.; Joy, M.; Chakraborty, R.D. Bacillibactin class of siderophore antibiotics from a marine symbiotic Bacillus as promising antibacterial agents. Appl. Microbiol. Biotechnol. 2022, 106, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Radi, M.H.; El-Shiekh, R.A.; El-Halawany, A.M.; Al-Abd, A.M.; Abdel-Sattar, E. In Vitro Cytotoxic Study of Euphorbia grantii Oliv. Aerial Parts against MCF-7 and MCF-7ADRBreast Cancer Cell Lines: A Bioactivity-Guided Isolation. ACS Omega 2023, 8, 18299–18305. [Google Scholar] [CrossRef]
- Kranjec, C.; Angeles, D.M.; Mårli, M.T.; Fernández, L.; García, P.; Kjos, M.; Diep, D.B. Staphylococcal Biofilms: Challenges and Novel Therapeutic Perspectives. Antibiotics 2021, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Valour, F.; Trouillet-Assant, S.; Rasigade, J.; Lustig, S.; Chanard, E.; Meugnier, H.; Tigaud, S.; Vandenesch, F.; Etienne, J.; Ferry, T.; et al. Staphylococcus epidermidis in Orthopedic Device Infections: The Role of Bacterial Internalization in Human Osteoblasts and Biofilm Formation. PLoS ONE 2013, 8, e67240. [Google Scholar] [CrossRef] [PubMed]
- Tande, A.J.; Patel, R. Prosthetic joint infection. Clin. Microbiol. Rev. 2014, 27, 302–345. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, M.; Post, V.; Erichsen, C.; Hungerer, S.; Bühren, V.; Militz, M.; Richards, R.G.; Moriarty, T.F. Biofilm formation increases treatment failure in Staphylococcus epidermidis device-related osteomyelitis of the lower extremity in human patients. J. Orthop. Res. 2016, 34, 1905–1913. [Google Scholar] [CrossRef]
- França, A.; Gaio, V.; Lopes, N.; Melo, D.R. Virulence Factors in Coagulase-Negative Staphylococci. Pathogens 2021, 10, 170. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef]
- Patil, A.; Banerji, R.; Kanojiya, P.; Saroj, S.D. Foodborne ESKAPE biofilms and antimicrobial resistance: Lessons learned from clinical isolates. Pathog. Glob. Health 2021, 15, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Xia, G.; Shi, C.; Wan, J.; Liu, L.; Chen, Y.; Wu, Y.; Zhang, W.; Zhou, M.; He, H.; et al. Therapeutic Strategies against Bacterial Biofilms. Fundam. Res. 2021, 1, 193–212. [Google Scholar] [CrossRef]
- Frassinetti, S.; Gabriele, M.; Moccia, E.; Longo, V.; Di Gioia, D. Antimicrobial and antibiofilm activity of Cannabis sativa L. seeds extract against Staphylococcus aureus and growth effects on probiotic Lactobacillus spp. LWT Food Sci. Technol. 2020, 124, 109149. [Google Scholar] [CrossRef]
- Menjivar, J.L.; Bendaoud, M. Anti-Biofilm Properties of Flax, Chia, and Hemp Seed Oil Extracts. FASEB J. 2020, 34 (Suppl. S1), 1. [Google Scholar] [CrossRef]
- Artini, M.; Papa, R.; Vrenna, G.; Lauro, C.; Ricciardelli, A.; Casillo, A.; Corsaro, M.M.; Tutino, L.; Parrilli, E.; Selan, L. Cold-adapted bacterial extracts as a source of anti-infective and antimicrobial compounds against Staphylococcus aureus. Future Microbiol. 2019, 14, 1369–1382. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, J.; Avula, B.; Wang, Y.H.; Avonto, C.; Chittiboyina, A.G.; Khan, I.A. High-resolution gas chromatography/mass spectrometry method for characterization and quantitative analysis of ginkgolic acids in Ginkgo biloba plants, extracts, and dietary supplements. J. Agric. Food Chem. 2014, 62, 12103–12111. [Google Scholar] [CrossRef]
- Al-Rubaye, A.F.; Hameed, I.H.; Kadhim, M.J. A review: Uses of gas chromatography-mass spectrometry (GC-MS) technique for analysis of bioactive natural compounds of some plants. Int. J. Toxicol. Pharmacol. Res. 2017, 9, 81–85. [Google Scholar] [CrossRef]
- Al-Tameme, H.J.; Hameed, I.H.; Idan, S.A.; Hadi, M.Y. Biochemical analysis of Origanum vulgare seeds by fourier-transform infrared (FT-IR) spectroscopy and gas chromatography-mass spectrometry (GC-MS). J. Pharmacogn. Phytother. 2015, 7, 221–237. [Google Scholar]
- Delgado-Povedano, M.M.; Callado, C.S.C.; Priego-Capote, F.; Ferreiro-Vera, C. Untargeted characterization of extracts from Cannabis sativa L. cultivars by gas and liquid chromatography coupled to mass spectrometry in high resolution mode. Talanta 2020, 208, 120384. [Google Scholar] [CrossRef]
- Gade, A.; Pinapati, K.K.; Verma, V.; Akula, S.; Sharma, A.; Pullapanthula, R.; Srivastava, N. Evaluation of Antibiofilm Activity of Alkaloids Extracted from Capsicum annuum Stalk: A Preliminary Study of Phytochemical Screening in Vegetable Waste. Waste Biomass Valorization 2023, 1–18. [Google Scholar] [CrossRef]
- Peschel, W.; Politi, M. 1H NMR and HPLC/DAD for Cannabis sativa L. chemotype distinction, extract profiling and specification. Talanta 2015, 140, 150–165. [Google Scholar] [CrossRef]
- Hamerly, T.; Tripet, B.; Wurch, L.; Hettich, R.L.; Podar, M.; Bothner, B.; Copié, V. Characterization of Fatty Acids in Crenarchaeota by GC-MS and NMR. Archaea 2015, 2015, 472726. [Google Scholar] [CrossRef] [PubMed]
- Exarchou, V.; Kanetis, L.; Charalambous, Z.; Apers, S.; Pieters, L.; Gekas, V.; Goulas, V. HPLC-SPE-NMR characterization of major metabolites in Salvia fruticosa Mill. extract with antifungal potential: Relevance of carnosic acid, carnosol, and hispidulin. J. Agric. Food Chem. 2015, 63, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, M.J.; Soengas, R.; Probert, I.; Guilloud, E.; Gourvil, P.; Mehiri, M.; Lopez, Y.; Cepas, V.; Gutierrez-del-Rio, I.; Redondo-Blanco, S.; et al. NMR characterization and evaluation of antibacterial and antiobiofilm activity of organic extracts from stationary phase batch cultures of five marine microalgae (Dunaliella sp., D. salina, Chaetoceros calcitrans, C. gracilis and Tisochrysis lutea). Phytochemistry 2019, 164, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthi, R.; Srinivash, M.; Mahalingam, P.U.; Malaikozhundan, B.; Suganya, P.; Gurushankar, K. Antimicrobial, anti-biofilm, antioxidant and cytotoxic effects of bacteriocin by Lactococcus lactis strain CH3 isolated from fermented dairy products—An in vitro and in silico approach. Int. J. Biol. Macromol. 2022, 220, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Mahomoodally, M.F.; Zengin, G.; Aladag, M.O.; Ozparlak, H.; Diuzheva, A.; Jekő, J.; Cziaky, Z.; Aumeeruddy, M.Z. HPLC-MS/MS chemical characterization and biological properties of Origanum onites extracts: A recent insight. Int. J. Environ. Health Res. 2019, 29, 607–621. [Google Scholar] [CrossRef]
- Mulinacci, N.; Romani, A.; Pinelli, P.; Vincieri, F.F.; Prucher, D. Characterization of Matricaria recutita L. flower extracts by HPLC-MS and HPLC-DAD analysis. Chromatographia 2000, 51, 301–307. [Google Scholar] [CrossRef]
- Benkirane, C.; Moumen, A.B.; Fauconnier, M.-L.; Belhaj, K.; Abid, M.; Serghini Caid, H.; Elamrani, A.; Mansouri, F. Bioactive compounds from hemp (Cannabis sativa L.) seeds: Optimization of phenolic antioxidant extraction using simplex lattice mixture design and HPLC-DAD/ESI-MS2 analysis. RSC Adv. 2022, 12, 25764–25777. [Google Scholar] [CrossRef]
- Raue, S.; Fan, S.-H.; Rosenstein, R.; Zabel, S.; Luqman, A.; Nieselt, K.; Götz, F. The Genome of Staphylococcus epidermidis O47. Front. Microbiol. 2020, 11, 2061. [Google Scholar] [CrossRef]
- Vuong, C.; Gerke, C.; Somerville, G.A.; Fischer, E.R.; Otto, M. Quorum-Sensing Control of Biofilm Factors in Staphylococcus epidermidis. J. Infect. Dis. 2003, 188, 706–718. [Google Scholar] [CrossRef]
- Gowrishankar, S.; Pandian, S.K. Modulation of Staphylococcus epidermidis (RP62A) Extracellular Polymeric Layer by Marine Cyclic Dipeptide-Cyclo(L-Leucyl-L-Prolyl) Thwarts Biofilm Formation. Biochim. Biophys. Acta BBA Biomembr. 2017, 1859, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C.; Schweitzer, O.; Gerke, C.; Vanittanakom, N.; Mack, D.; Götz, F. Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. Mol. Microbiol. 1996, 20, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Martí, M.; Trotonda, M.P.; Tormo-Más, M.Á.; Vergara-Irigaray, M.; Cheung, A.L.; Lasa, I.; Penadés, J.R. Extracellular proteases inhibit protein-dependent biofilm formation in Staphylococcus aureus. Microbes Infect. 2010, 12, 55–64. [Google Scholar] [CrossRef]
- Büttner, H.; Mack, D.; Rohde, H. Structural basis of Staphylococcus epidermidis biofilm formation: Mechanisms and molecular interactions. Front. Cell. Infect. Microbiol. 2015, 5, 14. [Google Scholar]
- Gill, S.R.; Fouts, D.E.; Archer, G.L.; Mongodin, E.F.; DeBoy, R.T.; Ravel, J.; Paulsen, I.T.; Kolonay, J.F.; Brinkac, L.; Beanan, M.; et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J. Bacteriol. 2005, 187, 2426–2438. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Conlan, S.; Mijares, L.A.; NISC Comparative Sequencing Program; Becker, J.; Blackeslet, R.W.; Bouffard, G.G.; Brooks, S.; Coleman, H.; Gupta, J.; Gurson, N.; et al. Staphylococcus epidermidis pan-genome sequence analysis reveals diversity of skin commensal and hospital infection-associated isolates. Genome Biol. 2012, 13, R64. [Google Scholar] [CrossRef]
- Cabrera-Contreras, R.; Santamaría, R.I.; Bustos, P.; Martínez-Flores, I.; Meléndez-Herrada, E.; Morelos-Ramírez, R.; Barbosa-Amezcua, M.; González-Covarrubias, V.; Silva-Herzog, E.; Soberón, X.; et al. Genomic diversity of prevalent Staphylococcus epidermidis multidrug-resistant strains isolated from a Children’s Hospital in México City in an eight-years survey. PeerJ 2019, 7, e8068. [Google Scholar] [CrossRef]
- Begović, J.; Jovčić, B.; Papić-Obradović, M.; Veljović, K.; Lukić, J.; Kojić, M.; Topisirović, L. Genotypic diversity and virulent factors of Staphylococcus epidermidis isolated from human breast milk. Microbiol. Res. 2013, 168, 77–83. [Google Scholar] [CrossRef]
- Chen, H.; Xu, B.; Wang, Y.; Li, W.; He, D.; Zhang, Y.; Zhang, X.; Xing, X. Emerging natural hemp seed proteins and their functions for nutraceutical applications. Food Sci. Hum. Wellness 2023, 12, 929–941. [Google Scholar] [CrossRef]
- Frassinetti, S.; Moccia, E.; Caltavuturo, L.; Gabriele, M.; Longo, V.; Bellani, L.; Giorgi, G.; Giorgetti, L. Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food Chem. 2018, 262, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, H.A.A.; Gomez d’Ayala, G.; Santagata, G.; Bosco, F.; Mollea, C.; Larsen, N.; Duraccio, D. Bioactive films based on barley β-glucans and ZnO for wound healing applications. Carbohydr. Polym. 2021, 272, 118442. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Zhu, P.; Wang, W.; Wang, M. The influence of extraction pH on the chemical compositions, macromolecular characteristics, and rheological properties of polysaccharide: The case of okra polysaccharide. Food Hydrocoll. 2020, 102, 105586. [Google Scholar] [CrossRef]
- Chen, C.; Liu, C.-H.; Cai, J.; Zhang, W.; Qi, W.-L.; Wang, Z.; Liu, Z.-B.; Yang, Y. Broad-spectrum antimicrobial activity, chemical composition and mechanism of action of garlic (Allium sativum) extracts. Food Control 2018, 86, 117–125. [Google Scholar] [CrossRef]
- Carillo, S.; Pieretti, G.; Parrilli, E.; Tutino, M.L.; Gemma, S.; Molteni, M.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Structural investigation and biological activity of the lipooligosaccharide from the psychrophilic bacterium Pseudoalteromonas haloplanktis TAB 23. Chem. Eur. J. 2011, 17, 7053–7060. [Google Scholar] [CrossRef]
- Corsaro, M.M.; Pieretti, G.; Lindner, B.; Lanzetta, R.; Parrilli, E.; Tutino, M.L.; Parrilli, M. Highly phosphorylated core oligosaccharide structures from cold-adapted Psychromonas arctica. Chem. A Eur. J. 2008, 14, 9368–9376. [Google Scholar] [CrossRef]
- Casillo, A.; Parrilli, E.; Sannino, F.; Buko, L.; Lanzetta, R.; Parrilli, M.; Tutino, M.L.; Corsaro, M.M. Structural Investigation of the Oligosaccharide Portion Isolated from the Lipooligosaccharide of the Permafrost Psychrophile Psychrobacter arcticus 273-4. Mar. Drugs 2015, 13, 4539–4555. [Google Scholar] [CrossRef]
- De Pinto, M.C.; Lavermicocca, P.; Evidente, A.; Corsaro, M.M.; Lazzaroni, S.; De Gara, L. Exopolysaccharides Produced by Plant Pathogenic Bacteria Affect Ascorbate Metabolism in Nicotiana tabacum. Plant Cell Physiol. 2003, 44, 803–810. [Google Scholar] [CrossRef]
- Heilmann, C.; Gerke, C.; Perdreau-Remington, F.; Götz, F. Characterization of Tn917 Insertion Mutants of Staphylococcus epidermidis affected in biofilm formation. Infect. Immun. 1996, 64, 277–282. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Bisno, A.L.; Beachey, E.H. Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect. Immun. 1982, 37, 318–326. [Google Scholar] [CrossRef]
- Papa, R.; Parrilli, E.; Sannino, F.; Barbato, G.; Tutino, M.L.; Artini, M.; Selan, L. Anti-Biofilm Activity of the Antarctic Marine Bacterium Pseudoalteromonas haloplanktis TAC125. Res. Microbiol. 2013, 164, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Di Lorenzo, F.; Imbimbo, P.; Silipo, A.; Bonina, A.; Rizza, L.; Piccoli, R.; Monti, D.M.; Lanzetta, R. Protective effect of Opuntia ficus-indica L. cladodes against UVA-induced oxidative stress in normal human keratinocytes. Bioorg. Med. Chem. Lett. 2017, 27, 5485–5489. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gro | Glu | Rha | Ara | Phe | GalA | Man | Gal | Glc | Ino | GalN/GlcN | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rt (min) | 3.98 | 8.64 | 10.11 | 10.68 | 12.10 | 18.26 | 18.88 | 19.18 | 19.52 | 23.14 | 24.58/24.61 |
| Ex7d | 2.6 | 26.9 | 7.6 | 16.2 | 10.3 | 8.0 | 2.3 | 17.3 | 5.5 | 0.2 | 3.1 |
| Ex4.5 | 1.2 | 7.9 | 10.6 | 15.3 | - | 19.7 | 3.1 | 15.5 | 20.4 | - | 6.3 |
| Ex7 | 24.4 | 4.2 | 0.7 | 6.7 | 4.1 | - | - | 22.9 | 31.3 | 5.7 | - |
| Ex10 | 21.8 | 2.6 | 0.4 | 6.8 | 1.5 | - | - | 22.3 | 39.5 | 5.1 | - |
| C14:0 | C16:0 | C18:2 | C18:1 | C18:0 | |
|---|---|---|---|---|---|
| Rt (min) | 8.75 | 10.96 | 12.64 | 12.69 | 12.94 |
| Ex7d | 2.7 | 31.5 | - | 21.5 | 44.3 |
| Ex4.5 | 3.5 | 30.2 | - | 25.2 | 41.1 |
| Ex7 | - | 13.3 | 47.4 | 31.5 | 7.8 |
| Ex10 | 1.2 | 22.1 | 19.5 | 32.5 | 24.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casillo, A.; D’Angelo, C.; Imbimbo, P.; Monti, D.M.; Parrilli, E.; Lanzetta, R.; Gomez d’Ayala, G.; Mallardo, S.; Corsaro, M.M.; Duraccio, D. Aqueous Extracts from Hemp Seeds as a New Weapon against Staphylococcus epidermidis Biofilms. Int. J. Mol. Sci. 2023, 24, 16026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216026
Casillo A, D’Angelo C, Imbimbo P, Monti DM, Parrilli E, Lanzetta R, Gomez d’Ayala G, Mallardo S, Corsaro MM, Duraccio D. Aqueous Extracts from Hemp Seeds as a New Weapon against Staphylococcus epidermidis Biofilms. International Journal of Molecular Sciences. 2023; 24(22):16026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216026
Chicago/Turabian StyleCasillo, Angela, Caterina D’Angelo, Paola Imbimbo, Daria Maria Monti, Ermenegilda Parrilli, Rosa Lanzetta, Giovanna Gomez d’Ayala, Salvatore Mallardo, Maria Michela Corsaro, and Donatella Duraccio. 2023. "Aqueous Extracts from Hemp Seeds as a New Weapon against Staphylococcus epidermidis Biofilms" International Journal of Molecular Sciences 24, no. 22: 16026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216026