
Data Mining Suggests That CXCL14 Gene Silencing in Colon Cancer Is Due to Promoter Methylation

Abstract
:1. Introduction
2. Result
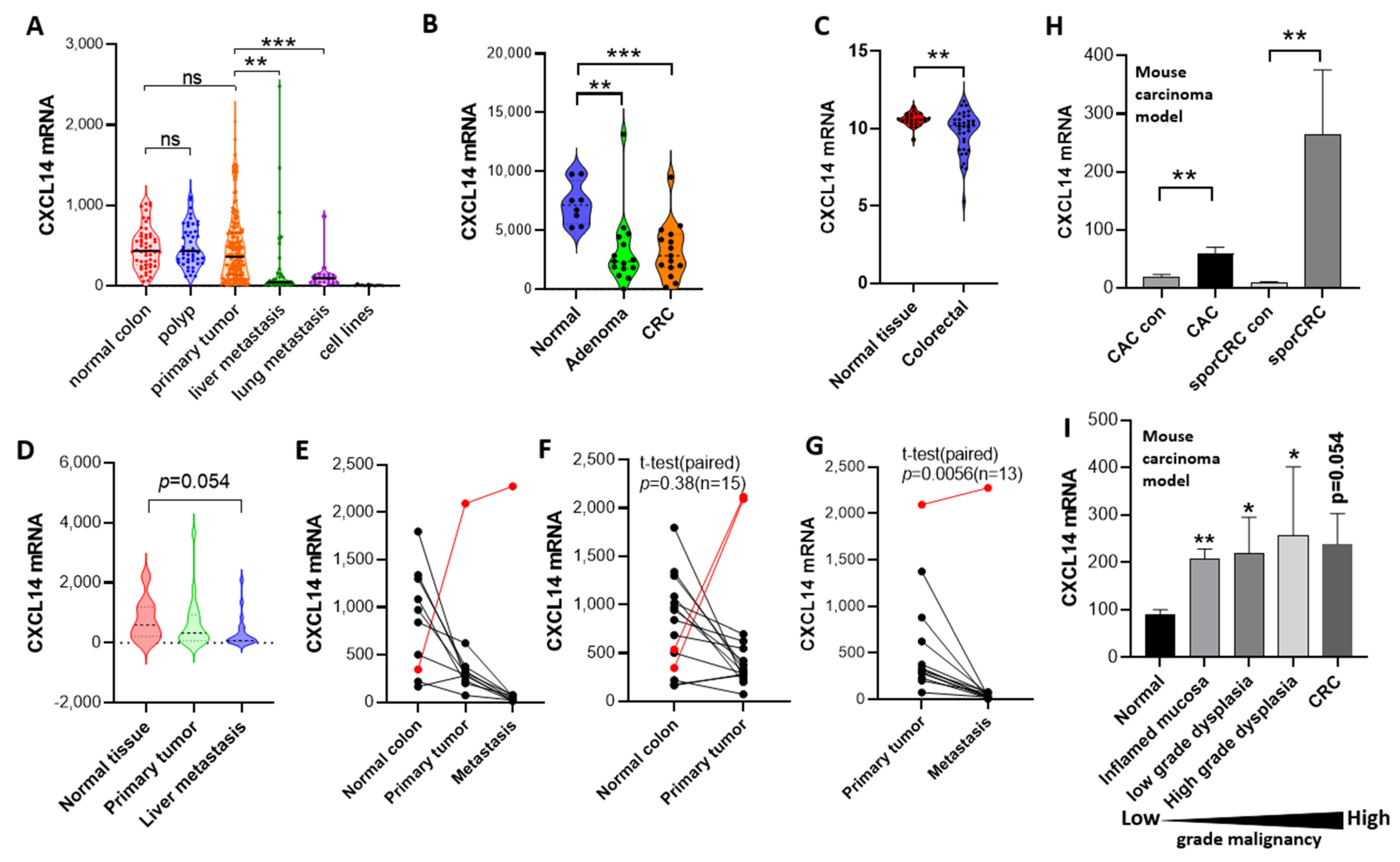
2.1. CXCL14 Silences as the Colorectal Cancer Progresses
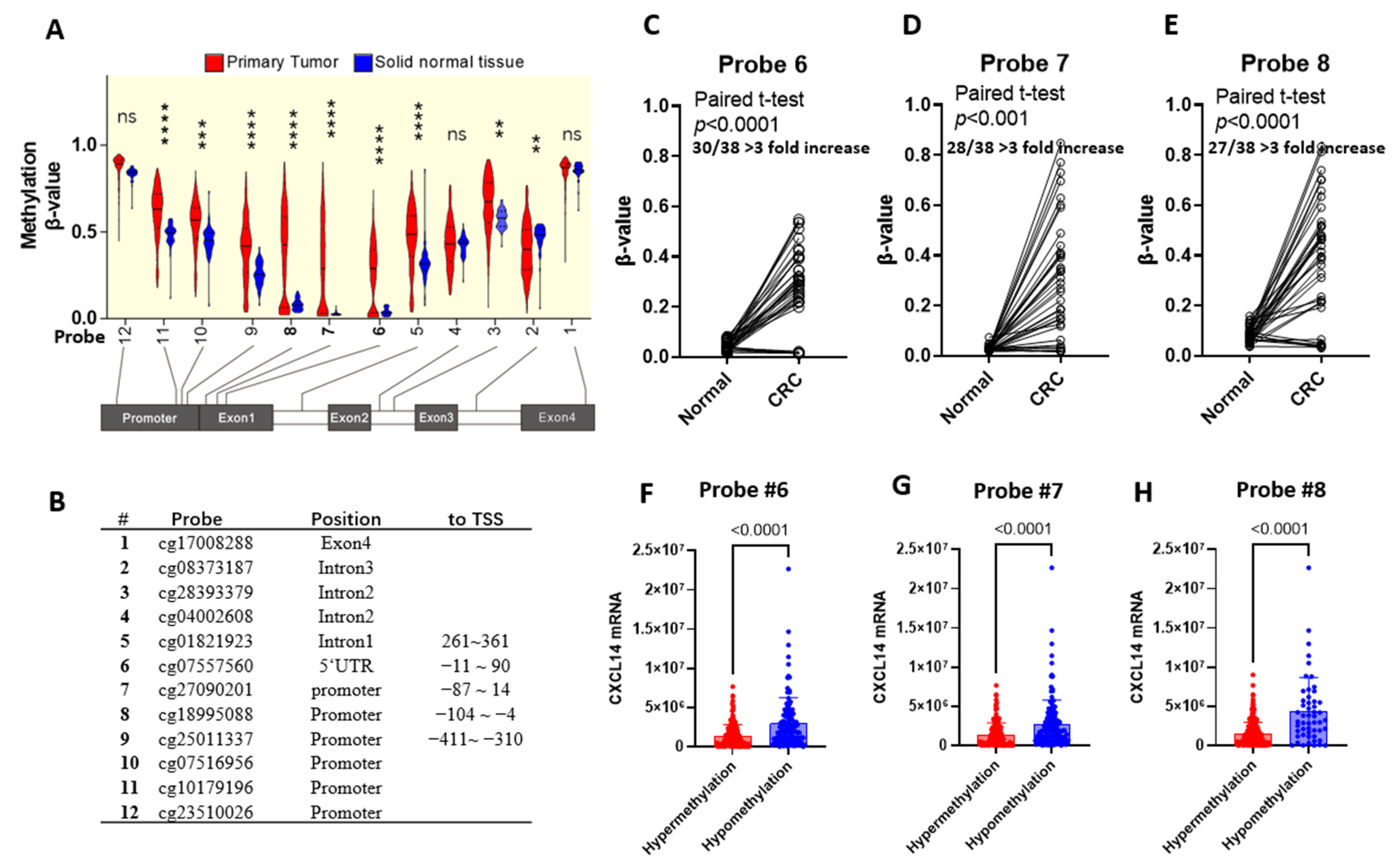
2.2. CXCL14 Silencing Is Associated with Promoter CpG Methylation
2.3. The Methylation of CXCL14 in Colon Cancer Is Closely Related to the Anatomical Site and Patient Age
2.4. CXCL14 Methylation and Senescence
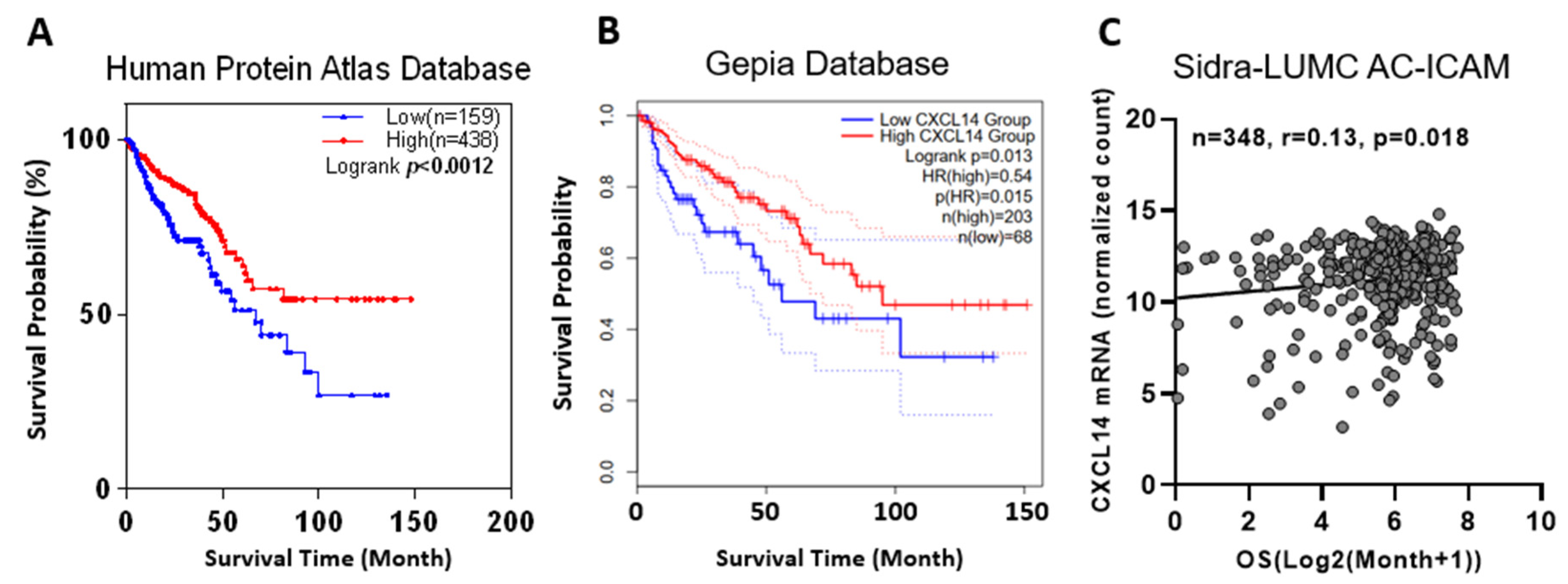
2.5. CXCL14 Methylation Was Associated with Survival
2.6. The Methylation and Demethylation of the CXCL14 Promoter Directly Regulate CXCL14 Expression
3. Methods
3.1. mRNA Expression Data
3.2. DNA Methylation Data
3.3. Survival Analysis
3.4. Cell Culture
3.5. Real-Time Quantitative PCR
3.6. Promoter Assay
3.7. Statistical Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shields, D.C. Molecular evolution of CXC chemokines and receptors. Trends Immunol. 2003, 24, 355. [Google Scholar] [CrossRef]
- Nara, N.; Nakayama, Y.; Okamoto, S.; Tamura, H.; Kiyono, M.; Muraoka, M.; Tanaka, K.; Taya, C.; Shitara, H.; Ishii, R.; et al. Disruption of CXC Motif Chemokine Ligand-14 in Mice Ameliorates Obesity-induced Insulin Resistance. J. Biol. Chem. 2007, 282, 30794–30803. [Google Scholar] [CrossRef] [PubMed]
- Shellenberger, T.D.; Wang, M.; Gujrati, M.; Jayakumar, A.; Strieter, R.M.; Burdick, M.D.; Ioannides, C.G.; Efferson, C.L.; El-Naggar, A.K.; Roberts, D.; et al. BRAK/CXCL14 is a potent inhibitor of angiogenesis and a chemotactic factor for immature dendritic cells. Cancer Res. 2004, 64, 8262–8270. [Google Scholar] [CrossRef] [PubMed]
- Kuang, H.; Chen, Q.; Zhang, Y.; Zhang, L.; Peng, H.; Ning, L.; Cao, Y.; Duan, E. The Cytokine Gene CXCL14 Restricts Human Trophoblast Cell Invasion by Suppressing Gelatinase Activity. Endocrinology 2009, 150, 5596–5605. [Google Scholar] [CrossRef] [PubMed]
- Huising, M.O.; Stet, R.J.; Kruiswijk, C.P.; Savelkoul, H.F.; Kemenade, B.L.V.-V. Molecular evolution of CXC chemokines: Extant CXC chemokines originate from the CNS. Trends Immunol. 2003, 24, 306–312. [Google Scholar] [CrossRef]
- Chen, L.; Guo, L.; Tian, J.; He, H.; Marinova, E.; Zhang, P.; Zheng, B.; Han, S. Overexpression of CXC Chemokine Ligand 14 Exacerbates Collagen-Induced Arthritis. J. Immunol. 2010, 184, 4455–4459. [Google Scholar] [CrossRef]
- Tessema, M.; Klinge, D.M.; Yingling, C.M.; Do, K.; Van Neste, L.; A Belinsky, S. Re-expression of CXCL14, a common target for epigenetic silencing in lung cancer, induces tumor necrosis. Oncogene 2010, 29, 5159–5170. [Google Scholar] [CrossRef] [PubMed]
- Song, E.Y.; Shurin, M.R.; Tourkova, I.L.; Gutkin, D.W.; Shurin, G.V. Epigenetic Mechanisms of Promigratory Chemokine CXCL14 Regulation in Human Prostate Cancer Cells. Cancer Res 2010, 70, 4394–4401. [Google Scholar] [CrossRef]
- Hara, T.; Nakayama, Y. CXCL14 and insulin action. Vitam. Horm. 2009, 80, 107–123. [Google Scholar]
- Kouzeli, A.; Collins, P.J.; Metzemaekers, M.; Meyrath, M.; Szpakowska, M.; Artinger, M.; Struyf, S.; Proost, P.; Chevigne, A.; Legler, D.F.; et al. CXCL14 Preferentially Synergizes with Homeostatic Chemokine Receptor Systems. Front. Immunol. 2020, 11, 561404. [Google Scholar] [CrossRef]
- Von Hundelshausen, P.; Agten, S.M.; Eckardt, V.; Blanchet, X.; Schmitt, M.M.; Ippel, H.; Neideck, C.; Bidzhekov, K.; Leberzammer, J.; Wichapong, K.; et al. Chemokine interactome mapping enables tailored intervention in acute and chronic inflammation. Sci. Transl. Med. 2017, 9, eaah6650. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Tanegashima, K. Pleiotropic functions of the CXC-type chemokine CXCL14 in mammals. J. Biochem. 2012, 151, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Kinzler, K.W. Cancer genes and the pathways they control. Nat. Med. 2004, 10, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Zardo, G.; Tiirikainen, M.I.; Hong, C.; Misra, A.; Feuerstein, B.G.; Volik, S.; Collins, C.C.; Lamborn, K.R.; Bollen, A.; Pinkel, D.; et al. Integrated genomic and epigenomic analyses pinpoint biallelic gene inactivation in tumors. Nat. Genet. 2002, 32, 453–458. [Google Scholar] [CrossRef]
- Cao, B.; Yang, Y.; Pan, Y.; Jia, Y.; Brock, M.V.; Herman, J.G.; Guo, M. Epigenetic silencing of CXCL14 induced colorectal cancer migration and invasion. Discov. Med. 2013, 16, 137–147. [Google Scholar]
- Hu, C.; Lin, F.; Zhu, G.; Xue, X.; Ding, Y.; Zhao, Z.; Zhang, L.; Shen, X. Abnormal hypermethylation of promoter region downregulates chemokine CXC ligand 14 expression in gastric cancer. Int. J. Oncol. 2013, 43, 1487–1494. [Google Scholar] [CrossRef]
- Sheffer, M.; Bacolod, M.D.; Zuk, O.; Giardina, S.F.; Pincas, H.; Barany, F.; Paty, P.B.; Gerald, W.L.; Notterman, D.A.; Domany, E. Association of survival and disease progression with chromosomal instability: A genomic exploration of colorectal cancer. Proc. Natl. Acad. Sci. USA 2009, 106, 7131–7136. [Google Scholar] [CrossRef]
- Galamb, O.; Györffy, B.; Sipos, F.; Spisák, S.; Németh, A.M.; Miheller, P.; Tulassay, Z.; Dinya, E.; Molnár, B. Inflammation, Adenoma and Cancer: Objective Classification of Colon Biopsy Specimens with Gene Expression Signature. Dis. Markers 2008, 25, 586721. [Google Scholar] [CrossRef]
- Del Rio, M.; Mollevi, C.; Vezzio-Vie, N.; Bibeau, F.; Ychou, M.; Martineau, P. Specific Extracellular Matrix Remodeling Signature of Colon Hepatic Metastases. PLoS ONE 2013, 8, e74599. [Google Scholar] [CrossRef]
- Neufert, C.; Becker, C.; Türeci, Ö.; Waldner, M.J.; Backert, I.; Floh, K.; Atreya, I.; Leppkes, M.; Jefremow, A.; Vieth, M.; et al. Tumor fibroblast–derived epiregulin promotes growth of colitis-associated neoplasms through ERK. J. Clin. Investig. 2013, 123, 1428–1443. [Google Scholar] [CrossRef]
- Tang, A.; Li, N.; Li, X.; Yang, H.; Wang, W.; Zhang, L.; Li, G.; Xiong, W.; Ma, J.; Shen, S. Dynamic activation of the key pathways: Linking colitis to colorectal cancer in a mouse model. Carcinogenesis 2012, 33, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Roelands, J.; Kuppen, P.J.K.; Ahmed, E.I.; Mall, R.; Masoodi, T.; Singh, P.; Monaco, G.; Raynaud, C.; de Miranda, N.F.; Ferraro, L.; et al. An integrated tumor, immune and microbiome atlas of colon cancer. Nat. Med. 2023, 29, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Hoadley, K.A.; Yau, C.; Hinoue, T.; Wolf, D.M.; Lazar, A.J.; Drill, E.; Shen, R.; Taylor, A.M.; Cherniack, A.D.; Thorsson, V.; et al. Cell-of-Origin Patterns Dominate the Molecular Classification of 10,000 Tumors from 33 Types of Cancer. Cell 2018, 173, 291–304.e6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, Y.; Wei, M.; Du, K.; Lin, J.; Wei, L. Comprehensive analysis of CXCL14 uncovers its role during liver metastasis in colon cancer. BMC Gastroenterol. 2023, 23, 1–12. [Google Scholar] [CrossRef]
- Liu, J.; Wang, D.; Zhang, C.; Zhang, Z.; Chen, X.; Lian, J.; Liu, J.; Wang, G.; Yuan, W.; Sun, Z.; et al. Identification of liver metastasis-associated genes in human colon carcinoma by mRNA profiling. Chin. J. Cancer Res. 2018, 30, 633–646. [Google Scholar] [CrossRef]
- Lin, K.; Zou, R.; Lin, F.; Zheng, S.; Shen, X.; Xue, X. Expression and effect of CXCL14 in colorectal carcinoma. Mol. Med. Rep. 2014, 10, 1561–1568. [Google Scholar] [CrossRef]
- Starnes, T.; Rasila, K.K.; Robertson, M.J.; Brahmi, Z.; Dahl, R.; Christopherson, K.; Hromas, R. The chemokine CXCL14 (BRAK) stimulates activated NK cell migration: Implications for the downregulation of CXCL14 in malignancy. Exp. Hematol. 2006, 34, 1101–1105. [Google Scholar] [CrossRef]
- Shurin, G.V.; Ferris, R.; Tourkova, I.L.; Perez, L.; Lokshin, A.; Balkir, L.; Collins, B.; Chatta, G.S.; Shurin, M.R. Loss of New Chemokine CXCL14 in Tumor Tissue Is Associated with Low Infiltration by Dendritic Cells (DC), while Restoration of Human CXCL14 Expression in Tumor Cells Causes Attraction of DC Both In Vitro and In Vivo. J. Immunol. 2005, 174, 5490–5498. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, Q.; Wu, X.; Yu, Y.; Zhang, H. Effect of chemokine CXCL14 on in vitro angiogenesis of human hepatocellular carcinoma cells. Arch. Physiol. Biochem. 2022, 128, 1316–1322. [Google Scholar] [CrossRef]
- Su, Z.; Kon, N.; Yi, J.; Zhao, H.; Zhang, W.; Tang, Q.; Li, H.; Kobayashi, H.; Li, Z.; Duan, S.; et al. Specific regulation of BACH1 by the hotspot mutant p53R175H reveals a distinct gain-of-function mechanism. Nat. Cancer 2023, 4, 564–581. [Google Scholar] [CrossRef]
- Seale, K.; Horvath, S.; Teschendorff, A.; Eynon, N.; Voisin, S. Making sense of the ageing methylome. Nat. Rev. Genet. 2022, 23, 585–605. [Google Scholar] [CrossRef] [PubMed]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y.; et al. Genome-wide Methylation Profiles Reveal Quantitative Views of Human Aging Rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, S.; Kato, Y.; Kubota, E.; Hata, R.-I. BRAK/CXCL14 expression in oral carcinoma cells completely suppresses tumor cell xenografts in SCID mouse. Biomed. Res. 2009, 30, 315–318. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample Type | Sample Number | Accession No. | Reference | |
|---|---|---|---|---|---|
| 1 | Human | Primary colon adenocarcinomas, adenomas, metastasis, and corresponding normal mucosae | 390 | E-GEOD-41258 | [17] |
| 2 | Human | 15 CRC, 15 with adenoma, and 8 healthy normal controls | 38 | E-GEOD-4183 | [18] |
| 3 | Human | 36 CRC tissues and 24 non-cancerous colorectal tissue | 60 | E-GEOD-23878 | |
| 4 | Human | Paired primary tumors and hepatic metastases and normal controls | 57 | E-GEOD-49355 | [19] |
| 5 | Human | Colorectal adenomas, adenocarcinomas, and normal control | 336 †/556 | TCGA-COAD PanCancer Atlas | [23] |
| 6 | Mouse | AOM/DSS induced CAC, ApcMin/+/J spontaneous tumor and para-carcinoma tissue | 14 | E-GEOD-43338 | [20] |
| 7 | Mouse | Ulcerative CAC | 18 | E-GEOD-31106 | [21] |
| Clinical Parameters | Number | CXCL14 Methylation Status | ||||||
|---|---|---|---|---|---|---|---|---|
| Unmethylated n = 84 (28%) | Methylated n = 212 (72%) | p-Value (χ2) | ||||||
| Gender | Female | 136 | (55%) | 36 | (44%) | 100 | (47%) | 0.614 |
| Male | 112 | (45%) | 46 | (56%) | 112 | (53%) | ||
| Race | Asian | 12 | (4%) | 3 | (4%) | 9 | (5%) | 0.33 |
| Black | 58 | (21%) | 21 | (27%) | 37 | (19%) | ||
| White | 206 | (75%) | 54 | (69%) | 152 | (77%) | ||
| Age | <60 | 98 | (33%) | 43 | (52%) | 55 | (26%) | <0.0001 |
| ≥60 | 196 | (67%) | 39 | (48%) | 157 | (74%) | ||
| anatomic neoplasm subdivision | cecum | 75 | (27%) | 13 | (16%) | 62 | (31%) | <106 |
| ascending colon | 55 | (20%) | 11 | (14%) | 44 | (22%) | ||
| hepatic flexure | 22 | (8%) | 0 | (0%) | 22 | (11%) | ||
| transverse colon | 25 | (9%) | 6 | (8%) | 19 | (10%) | ||
| splenic flexure | 5 | (2%) | 3 | (4%) | 2 | (1%) | ||
| descending colon | 14 | (5%) | 9 | (11%) | 5 | (3%) | ||
| sigmoid colon | 83 | (30%) | 38 | (48%) | 45 | (23%) | ||
| Poly | Yes | 77 | (35%) | 24 | (39%) | 53 | (34%) | 0.457 |
| No | 140 | (65%) | 37 | (61%) | 103 | (66%) | ||
| Kras gene mutation | Yes | 21 | (48%) | 11 | (61%) | 10 | (38%) | 0.139 |
| No | 23 | (52%) | 7 | (39%) | 16 | (62%) | ||
| lymphatic invasion | Yes | 79 | (31%) | 25 | (36%) | 54 | (29%) | 0.268 |
| No | 180 | (69%) | 45 | (64%) | 135 | (71%) | ||
| microsatellite instability | Yes | 11 | (13%) | 10 | (17%) | 1 | (3%) | 0.126 |
| No | 77 | (87%) | 48 | (82%) | 29 | (96%) | ||
| New tumor event | Yes | 62 | (23%) | 17 | (23%) | 45 | (23%) | 0.998 |
| No | 208 | (77%) | 57 | (77%) | 151 | (77%) | ||
| Perineural invasion present | Yes | 45 | (25%) | 14 | (30%) | 31 | (24%) | 0.423 |
| No | 132 | (75%) | 33 | (70%) | 99 | (76%) | ||
| neoplasm cancer status | Yes | 62 | (25%) | 23 | (33%) | 39 | (22%) | 0.069 |
| No | 187 | (75%) | 47 | (67%) | 140 | (78%) | ||
| Tumor stage | I | 44 | (15%) | 6 | (8%) | 38 | (18%) | 0.019 |
| II | 114 | (40%) | 30 | (39%) | 84 | (41%) | ||
| III | 85 | (30%) | 23 | (30%) | 62 | (30%) | ||
| IV | 41 | (14%) | 18 | (23%) | 23 | (11%) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, S.; Niu, Y.; Ma, B.; Li, J. Data Mining Suggests That CXCL14 Gene Silencing in Colon Cancer Is Due to Promoter Methylation. Int. J. Mol. Sci. 2023, 24, 16027. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216027
Wang Y, Wang S, Niu Y, Ma B, Li J. Data Mining Suggests That CXCL14 Gene Silencing in Colon Cancer Is Due to Promoter Methylation. International Journal of Molecular Sciences. 2023; 24(22):16027. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216027
Chicago/Turabian StyleWang, Yanjing, Siyi Wang, Yuchen Niu, Buyong Ma, and Jingjing Li. 2023. "Data Mining Suggests That CXCL14 Gene Silencing in Colon Cancer Is Due to Promoter Methylation" International Journal of Molecular Sciences 24, no. 22: 16027. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216027