
Establishment of a New Filamentous Cyanobacterial Genus, Microcoleusiopsis gen. nov. (Microcoleaceae, Cyanobacteria), from Benthic Mats in Open Channel, Jiangxi Province, China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Cultivation
2.2. Morphological and Ultrastructural Characterization
2.3. DNA Extraction and PCR Amplification
2.4. Detection for Cyanotoxin Synthesis Genes
2.5. Phylogenetic Analyses
2.6. Construction of Secondary Structure of 16S–23S Internal Transcribed Spacer (ITS)
3. Results
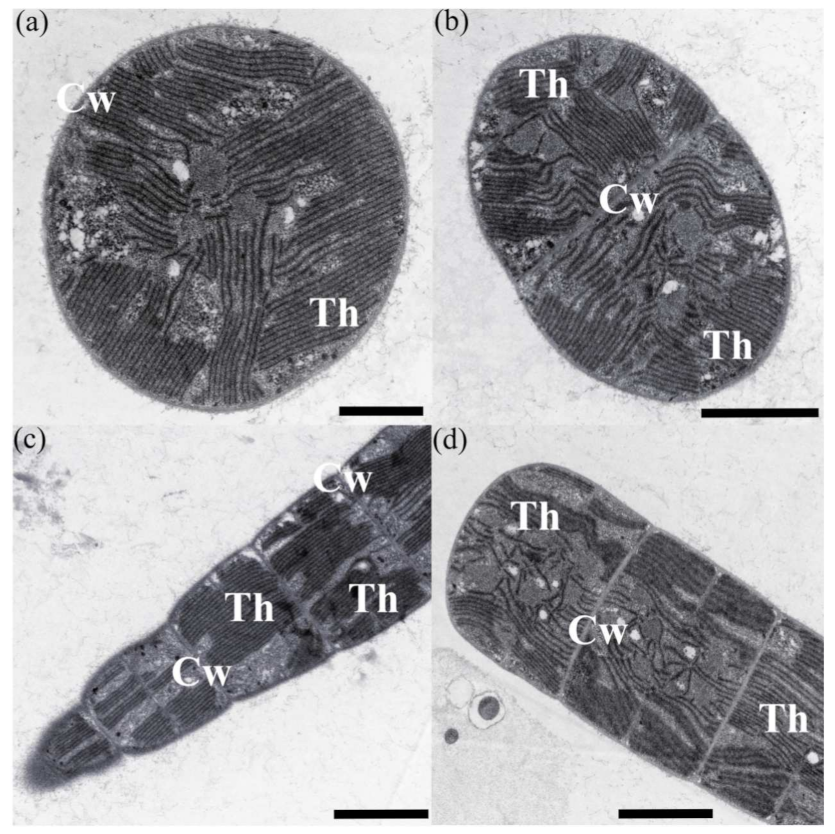
3.1. Morphological Description
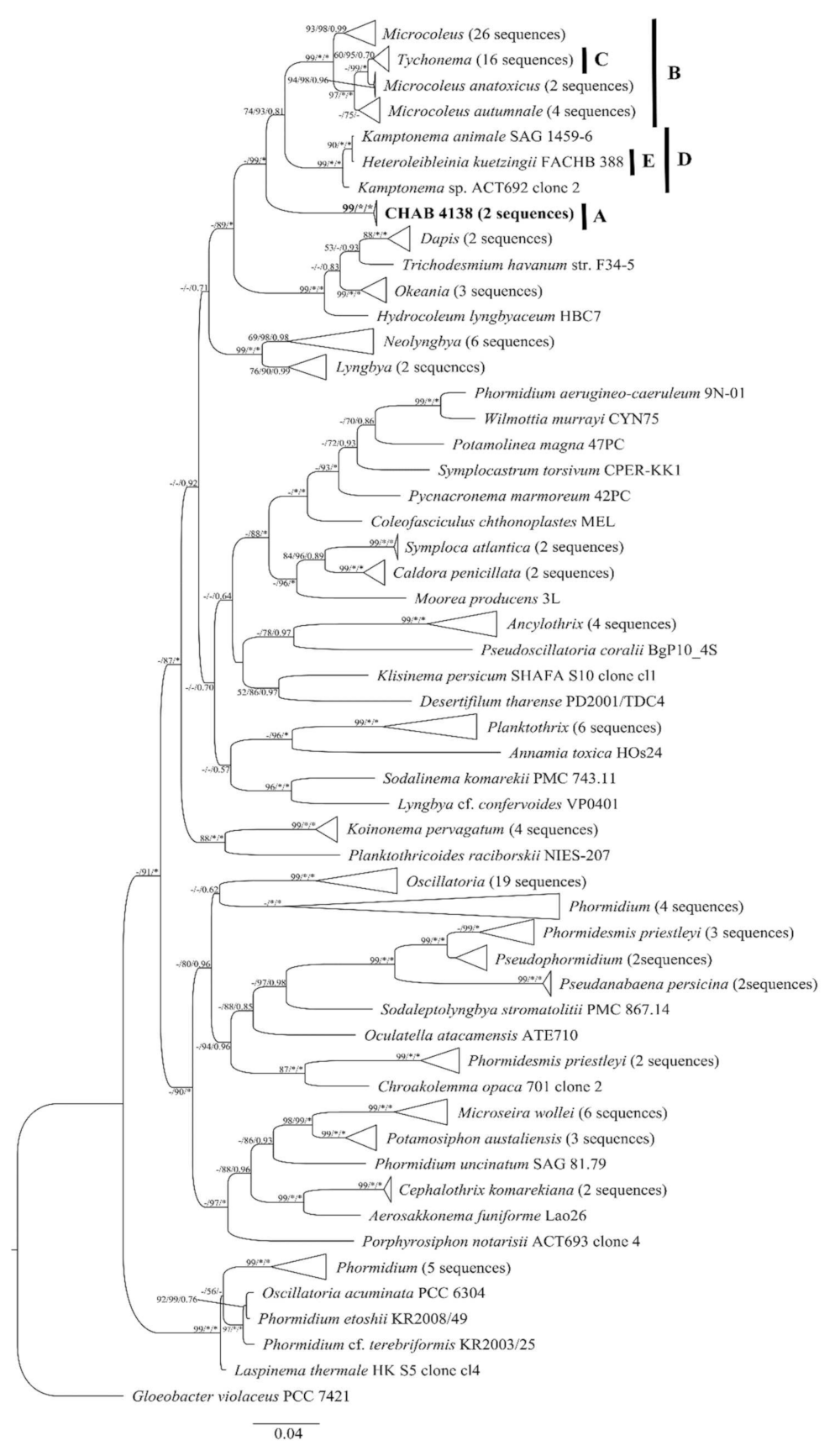
3.2. Molecular and Phylogeny Analyses
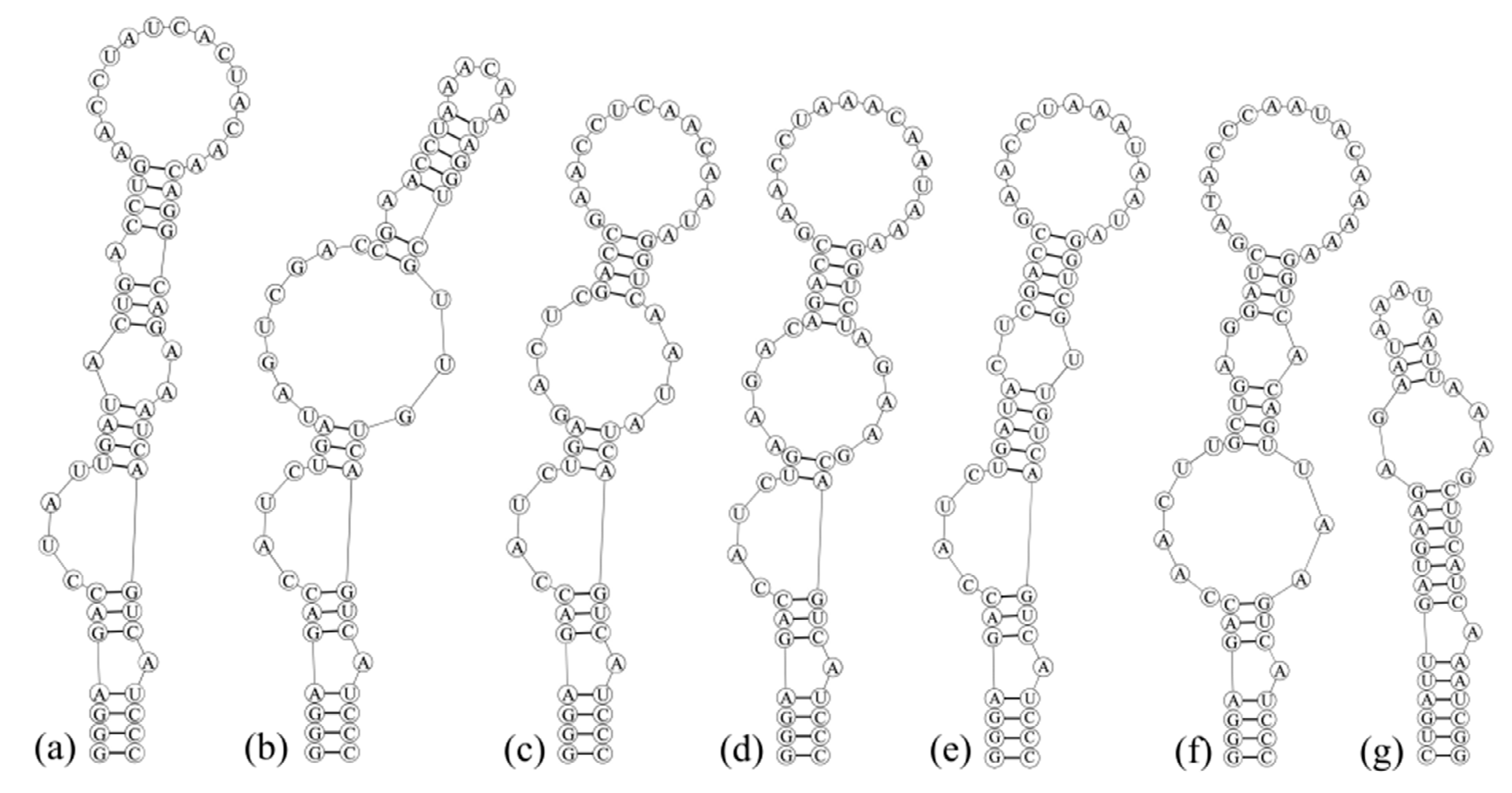
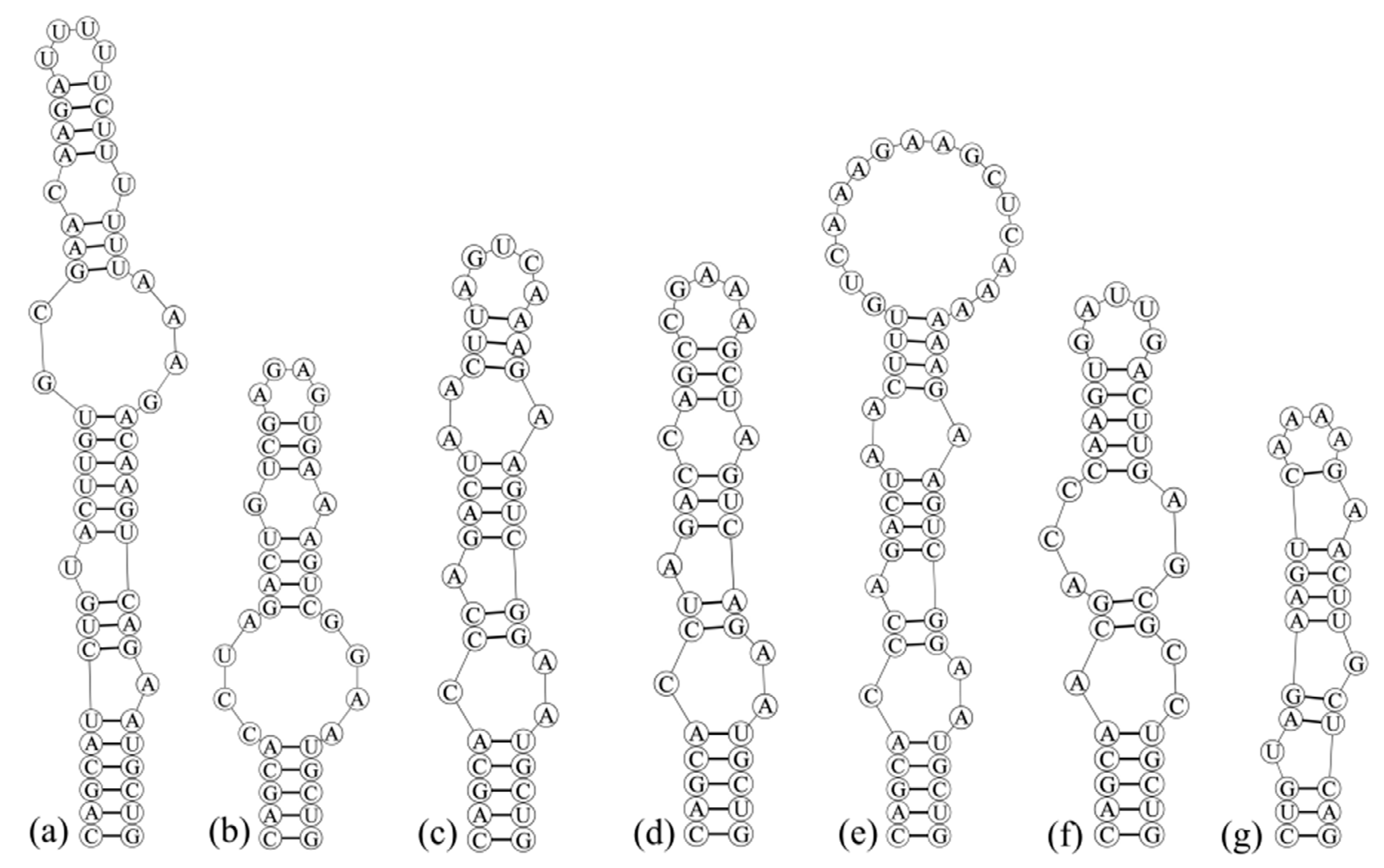
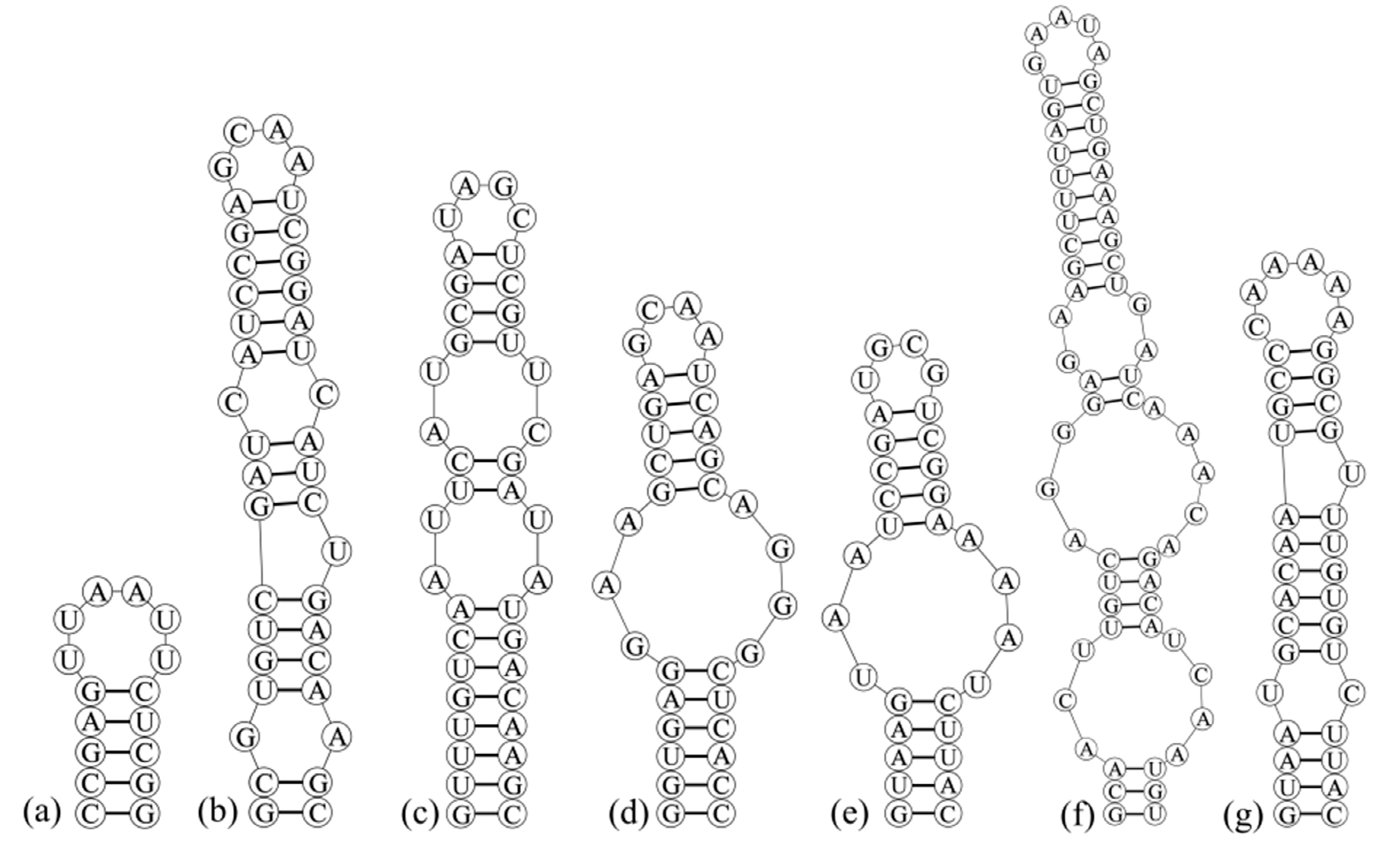
3.3. Analyses of ITS between 16S and 23S rRNA Gene and Secondary Structures
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia–Pichel, F.; Lopez–Cortes, A.; Nubel, U. Phylogenetic and morphological diversity of cyanobacteria in soil desert crusts from the Colorado plateau. Appl. Environ. Microbiol. 2001, 67, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komárek, J. Several problems of the polyphasic approach in the modern cyanobacterial system. Hydrobiologia 2018, 811, 7–17. [Google Scholar] [CrossRef]
- Mareš, J.; Strunecký, O.; Bučinská, L.; Wiedermannová, J. Evolutionary patterns of thylakoid architecture in cyanobacteria. Front. Microbiol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Comte, K.; Holland, D.P.; Walsby, A.E. Changes in cell turgor pressure related to uptake of solutes by Microcystis sp. strain 8401. FEMS Microbiol. Ecol. 2007, 61, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Sciuto, K.; Rascio, N.; Andreoli, C.; Moro, I. Polyphasic characterization of ITD–01, a cyanobacterium isolated from the Ischia Thermal District (Naples, Italy). Fottea. 2011, 11, 31–39. [Google Scholar] [CrossRef]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (Cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Dvořák, P.; Poulíčková, A.; Hašler, P.; Belli, M.; Casamatta, D.A.; Papini, A. Species concepts and speciation factors in cyanobacteria, with connection to the problems of diversity and classification. Biodivers. Conserv. 2015, 24, 739–757. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Johansen, J.R.; Šmarda, J.; Strunecký, O. Phylogeny and taxonomy of Synechococcus-like cyanobacteria. Fottea 2020, 20, 171–191. [Google Scholar] [CrossRef]
- Gomont, M. Monographie des Oscillariées (Nostocacées homocystées). Annal. Sci. Nat. Bot. 1892, 16, 91–264. [Google Scholar]
- Geitler, L. Schizophyta (Klasse Schizophyceae). In Natürliche Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Duncker & Humblot: Berlin, Germany, 1942; pp. 1–232. [Google Scholar]
- Drouet, F. Revision of the classifification of the Oscillatoriaceae. Monogr. Acad. Nat. Sci. Phil. 1968, 15, 1–370. [Google Scholar]
- Anagnostidis, K.; Komárek, J. Modern approach to the classification system of cyanophytes 3-Oscillatoriales. Algol. Stud. 1988, 50–53, 327–478. [Google Scholar]
- Boyer, S.L.; Johansen, J.R.; Flechtner, V.R. Phylogeny and Genetic Variance in Terrestrial Microcoleus (Cyanophyceae) Species Based on Sequence Analysis of the 16S rRNA Gene and Associated 16S–23S ITS Region. J. Phycol. 2002, 38, 1222–1235. [Google Scholar] [CrossRef]
- Siegesmund, M.A.; Johansen, J.R.; Karsten, U.; Friedl, T. Coleofasciculus gen. nov. (Cyanobacteria): Morphological and molecular criteria for revision of the genus Microcoleus Gomont. J. Phycol. 2008, 44, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Strunecký, O.; Komárekjeffrey, J.; Johansenjeffrey, R. Molecular and morphological criteria for revision of the genus Microcoleus (Oscillatoriales, Cyanobacteria). J. Phycol. 2013, 49, 1167–1180. [Google Scholar] [CrossRef] [PubMed]
- Niiyama, Y.; Tuji, A. Microcoleus pseudautumnalis sp. nov. (Cyanobacteria, Oscillatoriales) producing 2–methylisoborneol. Bull. Natl. Mus. Nat. Sci. Ser. B 2019, 45, 93–101. [Google Scholar]
- Conklin, K.Y.; Stancheva, R.; Otten, T.G. Molecular and morphological characterization of a novel dihydroanatoxin-a producing Microcoleus species (cyanobacteria) from the Russian River, California, USA. Harmful Algae 2020, 93, 101767. [Google Scholar] [CrossRef]
- Ichimura, T. Isolation and culture methods of algae. Methods Phycol. Stud. 1979, 2, 294–305. [Google Scholar]
- Spurr, A.R. A low–viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Goodman, A.E. Genetic diversity and phylogeny of toxic cyanobacteria determined by DNA polymorphisms within the phycocyanin locus. Appl. Environ. Microbiol. 1995, 61, 3875–3883. [Google Scholar] [CrossRef] [Green Version]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Rajaniemi, P.; Vardaka, E.; Moustaka–gouni, M.; Lanaras, T.; Sivonen, K. Limnothrix redekei (Van Goor) Meffert (Cyanobacteria) strains from Lake Kastoria, Greece form a separate phylogenetic group. Microb. Ecol. 2005, 49, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef]
- Al-Tebrineh, J.; Pearson, L.A.; Yasar, S.A.; Neilan, B.A. A multiplex qPCR targeting hepato- and neurotoxigenic cyanobacteria of global significance. Harmful Algae 2012, 15, 19–25. [Google Scholar] [CrossRef]
- Mcgregor, G.B.; Sendall, B.C. Phylogeny and toxicology of Lyngbya wollei (Cyanobacteria, Oscillatoriales) from north-eastern Australia, with a description of Microseira gen. nov. J. Phycol. 2015, 51, 109–119. [Google Scholar] [CrossRef]
- Rantala-Ylinen, A.; Känä, S.; Wang, H. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum–Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Page, R.D.M. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [PubMed] [Green Version]
- Mathews Lab. RNAstructure Version 5.6. Available online: http://rna.urmc.rochester.edu/RNAstructure.html (accessed on 13 May 2021).
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J. Cyanoprokaryota—3.Teil/ 3rd Part: Heterocytous Genera. In Süsswasserflora von Mitteleuropa Freshwater Flora of Central Europe 19/2; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Springer/Spektrum: Heidelberg, Germany, 2013. [Google Scholar]
- Anagnostidis, K.; Komárek, J. Modern approach to the classification system of cyanophytes. 1-Introduction. Algol. Stud. Arch. Für Hydrobiol. 1985, 38–39, 291–302. [Google Scholar]
- Johansen, J.R.; Casamatta, D.A. Recognizing cyanobacterial diversity through adoption of a new species paradigm. Algol. Studies 2005, 117, 71–93. [Google Scholar] [CrossRef]
- Casamatta, D.A.; Johansen, J.R.; Vis, M.L. Molecular and morphological characterization of ten polar and near–polar strains with the Oscillatoriales (Cyanobacteria). J. Phycol. 2010, 41, 421–438. [Google Scholar] [CrossRef]
- Iii, R.; Johansen, J.R.; Kovácik, L. A unique Pseudanabaenalean (cyanobacteria) genus Nodosilinea gen. nov. based on morphological and molecular data1. J. Phycol. 2011, 47, 1397–1412. [Google Scholar]
- Gugger, M.; Molica, R.; Le, B.B. Genetic diversity of Cylindrospermopsis strains (Cyanobacteria) isolated from four continents. Appl. Environ. Microbiol. 2005, 71, 1097–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strunecký, O.; Komárek, J.; Šmarda, J. Kamptonema (Microcoleaceae, Cyanobacteria), a new genus derived from the polyphyletic Phormidium on the basis of combined molecular and cytomorphological markers. Preslia 2014, 86, 193–207. [Google Scholar]
- Wang, Z.J.; Song, G.F.; Li, Y.G. The diversity, origin, and evolutionary analysis of geosmin synthase gene in cyanobacteria. Sci. Total. Environ. 2019, 689, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Starr, M.P. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Stackebrandt, E.; Goebel, B.M. Taxonomic note: A placefor DNA–DNA reassociation and 16S rRNA sequence analysisin the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiology 2006, 33, 152–155. [Google Scholar]
- Iteman, I.; Rippka, R.; Marsac, N.T.D.; Herdman, M. Comparison of conserved structural and regulatory domains within divergent 16s rRNA–23s rRNA spacer sequences of cyanobacteria. Microbiology 2000, 146, 1275–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaccarino, M.A.; Johansen, J.R. Brasilonema angustatum sp. nov. (Nostocales), a new filamentous cyanobacterial species from the Hawaiian Islands. J. Phycol. 2012, 48, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Osorio–Santos, K.; Pietrasiak, N.; Bohunická, M.; Miscoe, L.H.; Kováčik, L.; Martin, M.P.; Johansen, J.R. Seven new species of Oculatella (Pseudanabaenales, Cyanobacteria): Taxonomically recognizing cryptic diversification. Eur. J. Phycol. 2014, 49, 450–470. [Google Scholar] [CrossRef] [Green Version]
- Pietrasiak, N.; Mühlsteinová, R.; Siegesmund, M.A.; Johansen, J.R. Phylogenetic placement of Symplocastrum (Phormidiaceae, Cyanophyceae) with a new combination S. californicum and two new species: S. flechtnerae and S. torsivum. Phycologia 2014, 53, 529–541. [Google Scholar] [CrossRef]
- Mareš, J. Multilocus and SSU rRNA gene phylogenetic analyses of available cyanobacterial genomes, and their relation to the current taxonomic system. Hydrobiologia 2018, 811, 19–34. [Google Scholar] [CrossRef]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Fetscher, A.E.; Howard, M.D.; Stancheva, R.; Kudela, R.M.; Stein, E.D.; Sutula, M.A.; Busse, L.B.; Sheath, R.G. Wadeable streams as widespread sources of benthic cyanotoxins in California, USA. Harmful Algae 2015, 49, 105–116. [Google Scholar] [CrossRef]
- Echenique-Subiabre, I.; Dalle, C.; Duval, C.; Heath, M.W.; Coute, A.; Wood, S.A.; Humbert, J.-F.; Quiblier, C. Application of a spectrofluorimetric tool (bbe BenthoTorch) for monitoring potentially toxic benthic cyanobacteria in rivers. Water Res. 2016, 101, 341–350. [Google Scholar] [CrossRef]
- Bouma-Gregson, K.; Olm, M.R.; Probst, A.J.; Anantharaman, K.; Power, M.E.; Banfield, J.F. Impacts of microbial assemblage and environmental conditions on the distribution of anatoxin-a producing cyanobacteria within a river network. ISME J. 2019, 13, 1618–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shams, S.; Capelli, C.; Cerasino, L.; Ballot, A.; Dietrich, D.R.; Sivonen, K.; Salmaso, N. Anatoxin-a producing Tychonema (cyanobacteria) in European waterbodies. Water Res. 2015, 69, 68–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nico, S.; Leonardo, C.; Adriano, B.; Camilla, C. Planktic Tychonema (cyanobacteria) in the large lakes south of the alps: Phylogenetic assessment and toxigenic potential. Fems Microbiol. Ecol. 2016, 92, fiw155. [Google Scholar]
- Jutta, F.; Camilla, B.; Britta, G.; Anja, H.; Roswitha, K.; Kinga, T. Fatal neurotoxicosis in dogs associated with tychoplanktic, anatoxin-a producing Tychonema sp. in mesotrophic lake tegel, Berlin. Toxins 2018, 10, 60. [Google Scholar]
- Blahova, L.; Sehnal, L.; Lepsova-Skacelova, O.; Szmucova, V.; Babica, P.; Hilscherova, K.; Teikari, J.; Sivonen, K.; Blaha, L. Occurrence of cylindrospermopsin, anatoxin-a and their homologs in the southern Czech Republic–Taxonomical, analytical, and molecular approaches. Harmful Algae 2021, 108, 102101. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1.Microcoleusiopsis ganfuensisCHAB 4138 clone 1 | ||||||||||||||||||||||||
| 2.Microcoleusiopsis ganfuensisCHAB 4138 clone 2 | 99.91 | |||||||||||||||||||||||
| 3. Microcoleus pseudautumnalis Ak1609 | 94.70 | 94.78 | ||||||||||||||||||||||
| 4. Microcoleus vaginatus CSU-U-KK1 | 94.96 | 95.04 | 99.39 | |||||||||||||||||||||
| 5. Microcoleus anatoxicus PTRS-2 | 94.26 | 94.35 | 98.70 | 98.35 | ||||||||||||||||||||
| 6. Microcoleus autumnale SAG 78.79 | 94.09 | 94.17 | 98.52 | 98.00 | 99.13 | |||||||||||||||||||
| 7. Oscillatoria princeps CCALA 1115 clone F3 | 92.00 | 92.09 | 93.04 | 92.61 | 92.78 | 92.52 | ||||||||||||||||||
| 8. Phormidium etoshii KR2008/49 | 91.74 | 91.83 | 92.70 | 92.78 | 92.70 | 92.78 | 92.61 | |||||||||||||||||
| 9. Tychonema bourrellyi FEM GT529 | 94.26 | 94.35 | 98.61 | 98.09 | 99.74 | 99.22 | 92.78 | 92.52 | ||||||||||||||||
| 10. Tychonema bornetii NIVA-CYA 60 | 94.26 | 94.35 | 98.61 | 98.09 | 99.74 | 99.22 | 92.78 | 92.52 | 100.00 | |||||||||||||||
| 11. Kamptonema animale SAG 1459-6 | 94.43 | 94.52 | 96.17 | 95.74 | 95.91 | 95.57 | 93.39 | 91.57 | 95.74 | 95.74 | ||||||||||||||
| 12. Heteroleibleinia kutzingii FACHB 388 | 94.43 | 94.52 | 96.17 | 95.74 | 95.91 | 95.57 | 93.39 | 91.57 | 95.74 | 95.74 | 100.00 | |||||||||||||
| 13. Lyngbya hieronymusii CN4-3 | 93.83 | 93.91 | 93.39 | 93.65 | 93.30 | 93.65 | 91.91 | 93.30 | 93.30 | 93.30 | 93.30 | 93.30 | ||||||||||||
| 14. Neolyngbya granulosa ALCB 114393 | 94.09 | 94.17 | 93.13 | 93.39 | 92.35 | 92.52 | 91.22 | 92.78 | 92.17 | 92.17 | 92.61 | 92.61 | 96.26 | |||||||||||
| 15. Coleofasciculus chthonoplastes MEL | 92.09 | 92.17 | 92.52 | 92.78 | 92.26 | 92.26 | 92.26 | 91.48 | 92.09 | 92.09 | 93.83 | 93.83 | 93.39 | 93.39 | ||||||||||
| 16. Hydrocoleum lyngbyaceum HBC7 | 92.96 | 93.04 | 93.74 | 93.48 | 92.96 | 93.13 | 91.57 | 91.74 | 92.96 | 92.96 | 93.57 | 93.57 | 93.39 | 93.30 | 92.52 | |||||||||
| 17. Okeania plumata NAC8-45 | 93.13 | 93.22 | 94.09 | 93.65 | 93.83 | 94.00 | 90.87 | 91.74 | 93.83 | 93.83 | 94.35 | 94.35 | 93.39 | 92.96 | 92.43 | 97.30 | ||||||||
| 18. Symploca atlantica PCC 8002 | 92.61 | 92.70 | 93.30 | 92.96 | 93.13 | 92.61 | 92.61 | 91.39 | 92.87 | 92.87 | 93.48 | 93.48 | 92.87 | 92.52 | 94.00 | 92.09 | 92.70 | |||||||
| 19. Wilmottia murrayi CYN75 | 92.43 | 92.52 | 93.04 | 93.22 | 93.22 | 92.87 | 91.91 | 90.78 | 92.96 | 92.96 | 93.22 | 93.22 | 92.52 | 92.26 | 94.09 | 91.65 | 92.70 | 93.74 | ||||||
| 20. Aerosakkonema funiforme Lao26 | 92.00 | 92.09 | 93.04 | 93.30 | 92.09 | 92.26 | 92.70 | 90.78 | 92.00 | 92.00 | 92.78 | 92.78 | 92.09 | 92.09 | 92.87 | 92.35 | 91.65 | 92.43 | 92.09 | |||||
| 21. Dapis pnigousa BCBC12-12 | 92.78 | 92.87 | 93.91 | 93.48 | 93.65 | 93.83 | 91.22 | 91.13 | 93.65 | 93.65 | 93.91 | 93.91 | 93.13 | 92.17 | 92.70 | 97.04 | 97.04 | 92.09 | 92.09 | 91.39 | ||||
| 22. Moorea producens 3L | 92.70 | 92.78 | 91.48 | 91.04 | 91.30 | 91.30 | 91.65 | 90.87 | 91.30 | 91.30 | 93.04 | 93.04 | 92.17 | 92.00 | 93.57 | 91.74 | 92.17 | 94.26 | 92.17 | 90.78 | 91.57 | |||
| 23. Caldora penicillata HMC13-9 | 92.52 | 92.61 | 92.78 | 92.35 | 92.26 | 92.26 | 92.87 | 92.09 | 92.17 | 92.17 | 93.30 | 93.30 | 92.52 | 92.78 | 94.87 | 92.09 | 92.26 | 96.09 | 93.65 | 92.17 | 92.43 | 94.43 | ||
| 24. Laspinema thermale HK S5 clone cl4 | 92.43 | 92.52 | 93.30 | 93.57 | 93.04 | 93.13 | 93.22 | 98.96 | 92.87 | 92.87 | 92.26 | 92.26 | 93.65 | 93.48 | 91.57 | 92.52 | 92.09 | 91.91 | 91.13 | 91.39 | 91.39 | 91.57 | 92.52 | |
| 25. Trichodesmium havanum str. F34-5 | 92.09 | 92.17 | 93.65 | 93.22 | 93.04 | 93.39 | 90.70 | 91.57 | 93.04 | 93.04 | 93.13 | 93.13 | 92.35 | 92.35 | 91.57 | 96.52 | 97.22 | 91.57 | 91.57 | 90.87 | 97.22 | 90.78 | 91.83 | 91.83 |
| Organisms | GenBank | ITS Total Length (nt) | D1–D1′ Helix Length (nt) | D2 Region | tRNAIle | tRNAAla | Box B Helix Length (nt) | Box A Spacer | V3 Helix Length (nt) |
|---|---|---|---|---|---|---|---|---|---|
| Microcoleusiopsis ganfuensisCHAB 4138 | OK422506 | 761 | 60 | CTTTCAAACTAG | + | + | 58 | GAACCTTGAAAA | 16 |
| Microcoleus pseudautumnalis Ak1609 | LC486302 | 545 | 58 | CTTTCAAACTAT | + | + | 38 | GAACCTTGAAAA | 39 |
| Microcoleus vaginatus CSU-U-KK1 | EF667962 | 586 | 60 | CTTTCAAACTAT | + | + | 40 | GAACCTTGAAAA | 40 |
| Microcoleus anatoxicus PTRS-2 | MT013208 | 548 | 63 | CTTTCAAACTAT | + | + | 37 | GAACCTTGAAAA | 33 |
| Oscillatoria princeps CCALA 1115 clone F3 | MG255277 | 746 | 60 | CTTTCAAACTAA | + | + | 37 | GAACCTTGAAAA | 62 |
| Microcoleus autumnale SAG 78.79 | AM778717 | 573 | 58 | CTTTCAAACTAT | + | + | 53 | GAACCTTGAAAA | 31 |
| Coleofasciculus chthonoplastes MEL | EF654038 | 526 | 44 | CTTTCAAACTGG | + | + | 27 | GAACCTTGAAAA | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, R.; Li, W.; Chao, A.; Guo, X.; Li, H.; Yu, G.; Li, R. Establishment of a New Filamentous Cyanobacterial Genus, Microcoleusiopsis gen. nov. (Microcoleaceae, Cyanobacteria), from Benthic Mats in Open Channel, Jiangxi Province, China. Diversity 2021, 13, 548. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110548
Geng R, Li W, Chao A, Guo X, Li H, Yu G, Li R. Establishment of a New Filamentous Cyanobacterial Genus, Microcoleusiopsis gen. nov. (Microcoleaceae, Cyanobacteria), from Benthic Mats in Open Channel, Jiangxi Province, China. Diversity. 2021; 13(11):548. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110548
Chicago/Turabian StyleGeng, Ruozhen, Wenke Li, Aimin Chao, Xiaoyu Guo, Hua Li, Gongliang Yu, and Renhui Li. 2021. "Establishment of a New Filamentous Cyanobacterial Genus, Microcoleusiopsis gen. nov. (Microcoleaceae, Cyanobacteria), from Benthic Mats in Open Channel, Jiangxi Province, China" Diversity 13, no. 11: 548. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110548