
Three New Diatom Species from Spring Habitats in the Northern Apennines (Emilia-Romagna, Italy)

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cantonati, M.; Segadelli, S.; Spitale, D.; Gabrieli, J.; Gerecke, R.; Angeli, N.; De Nardo, M.T.; Ogata, K.; Wehr, J.D. Geological and hydrochemical prerequisites of unexpectedly high biodiversity in spring ecosystems at the landscape level. Sci. Total Environ. 2020, 740, 140157. [Google Scholar] [CrossRef]
- Cantonati, M.; Füreder, L.; Gerecke, R.; Jüttner, I.; Cox, E.J. Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 2012, 31, 463–480. [Google Scholar] [CrossRef]
- Cantonati, M.; Stevens, L.; Segadelli, S.; Springer, A.; Goldscheider, N.; Celico, F.; Filippini, M.; Ogata, K.; Gargini, A. Ecohydrogeology: The interdisciplinary convergence needed to improve the study and stewardship of springs and other groundwater-dependent habitats, biota, and ecosystems. Ecol. Indic. 2020, 110, 1. [Google Scholar] [CrossRef]
- Cantonati, M.; Fensham, R.J.; Stevens, L.E.; Gerecke, R.; Glazier, D.S.; Goldscheider, N.; Knight, R.L.; Richardson, J.S.; Springer, A.E.; Tockner, K. Urgent plea for global protection of springs. Conserv. Biol. 2021, 35, 378–382. [Google Scholar] [CrossRef]
- Cantonati, M.; Hofmann, G.; Spitale, D.; Werum, M.; Lange-Bertalot, H. Diatom Red Lists: Important tools to assess and preserve biodiversity and habitats in the face of direct impacts and environmental change. Biodivers. Conserv. 2021. Accepted. [Google Scholar]
- Cantonati, M.; Angeli, N.; Lange-Bertalot, H. Three new Fragilaria species (Bacillariophyta) from low-conductivity mountain freshwaters (Alps and Apennines). Phytotaxa 2019, 404, 261–274. [Google Scholar] [CrossRef]
- Bahls, L. Diatom indicators of climate change in Glacier National Park. Intermt J. Sci. 2007, 13, 99–109. [Google Scholar]
- Cantonati, M.; Segadelli, S.; Ogata, K.; Tran, H.; Sanders, D.; Gerecke, R.; Rott, E.; Filippini, M.; Gargini, A.; Celico, F. A global review on ambient Limestone-Precipitating Springs (LPS): Hydrogeological setting, ecology, and conservation. Sci. Total. Environ. 2016, 568, 624–637. [Google Scholar] [CrossRef]
- Cantonati, M.; Angeli, N.; Bertuzzi, E.; Spitale, D.; Lange-Bertalot, H. Diatoms in springs of the Alps: Spring types, environmental determinants, and substratum. Freshw. Sci. 2012, 31, 499–524. [Google Scholar] [CrossRef]
- Werum, M.; Lange-Bertalot, H. Diatoms in springs, from Central Europe and elsewhere under the influence of hydrogeology and anthropogenic impages. Iconogr. Diatomol. 2004, 13, 1–417. [Google Scholar]
- Cantonati, M.; Lange-Bertalot, H. Diatom biodiversity of springs in the berchtesgaden national park (north-eastern alps, germany), with the ecological and morphological characterization of two species new to science. Diatom Res. 2010, 25, 251–280. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H. Diatom monitors of close-to-pristine, very-low alkalinity habitats: Three new Eunotia species from springs in Nature Parks of the south-eastern Alps. J. Limnol. 2011, 70, 209–221. [Google Scholar] [CrossRef]
- Krammer, K. Cymbella Diatoms of Europe, Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag K.G.: Rugell, Liechtenstein, 2002; Volume 3, pp. 1–584. [Google Scholar]
- Maltsev, Y.; Maltseva, S.; Kociolek, J.P.; Jahn, R.; Kulikovskiy, M. Biogeography of the cosmopolitan terrestrial diatom Hantzschia amphioxys sensu lato based on molecular and morphological data. Sci. Rep. 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Geitler, L. Der Formwechsel der pennaten Diatomeen (Kieselalgen) Arch. Protistenkunde. 1932, 78, 1–226. [Google Scholar]
- Lange-Bertalot, H.; Bąk, M.; Witkowski, A. Eunotia and some related genera. In Diatoms of Europe. Diatoms of the European in-Land Water and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag K.G.: Rugell, Liechtenstein, 2011; Volume 6, pp. 1–747. [Google Scholar]
- Blasi, C.; Del Vico, E. High mountain vegetation of the Apennines. Ber. d. Reinh.-Tüxen-Ges. 2012, 24, 179–194. [Google Scholar]
- Cantonati, M.; Angeli, N.; Lange-Bertalot, H.; Levkov, Z. New Amphora and Halamphora (Bacillariophyta) species from springs in the northern Apennines (Emilia-Romagna, Italy). Plant Ecol. Evol. 2019, 152, 285–292. [Google Scholar] [CrossRef]
- Ehrenberg, C.G. Über ein aus Fossilen Infusorien Bestehendes, 1832 zu Brod Verbacknes Bergmehl von den Grenzen Lapplands in Schweden. Bericht über die zur Bekanntmachung Geeigneten Verhandlungen der Königl; Preuß. Akademie der Wissenschaften zu Berlin Erster Jarhrgang: Berlin, Germany, 1837; pp. 43–45. [Google Scholar]
- Novitski, L.; Kociolek, P. Preliminary Light and Scanning Electron Microscope Observations of Marine Fossil Eunotia Species with Comments on the Evolution of the Genus Eunotia. Diatom Res. 2005, 20, 137–143. [Google Scholar] [CrossRef]
- Luo, F.; You, Q.; Yu, P.; Pang, W.; Wang, Q. Eunotia (Bacillariophyta) biodiversity from high altitude, freshwater habitats in the Mugecuo Scenic Area, Sichuan Province, China. Phytotaxa 2019, 394, 133–147. [Google Scholar] [CrossRef] [Green Version]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, CA, USA, 1990; pp. 1–747. [Google Scholar]
- Kociolek, J.P.; Spaulding, S. Eunotioid and asymmetrical naviculoid diatoms. In Freshwater Algae of North America. Ecology and classification; Wehr, J.D., Sheath, R.G.F., Eds.; Academic Press Elsevier Science: London, UK, 2003; pp. 655–668. [Google Scholar]
- Furey, P.C.; Lowe, R.L.; Johansen, R.J. Eunotia Ehrenberg (Bacillariophyta) of the Great Smoky Mountains National Park, USA. Biblioth. Diatomol. 2011, 56, 1–133. [Google Scholar]
- Cox, E. Coscinodiscophyceae, Mediophyceae, Fragilariophyceae, Bacillariophyceae (Diatoms). In Syllabus of Plant Families. Adolf Engler’s Syllabus der Pflanzenfamilien. 2/1 Photoautotrophic Eukaryotic Algae; Frey, W., Ed.; Borntraeger Science Publishers: Stuttgart, Germany, 2015; pp. 64–103. [Google Scholar]
- Metzeltin, D.; Lange-Bertalot, H. Tropical diatoms of South America I: About 700 predominantly rarely known or new taxa representative of the neotropical flora. In Iconographia Diatomologica. Annotated Diatom Micrographs; Lange-Bertalot, H., Ed.; Diversity-Taxonomy-Geobotany, Koeltz Scientific Books: Königstein, Germany, 1998; Volume 5, pp. 1–695. [Google Scholar]
- Liu, Y.; Wang, Q.; Fu, C. Taxonomy and distribution of diatoms in the genus Eunotia from the Da’erbin Lake and Surrounding Bogs in the Great Xing’an Mountains, China. Nova Hedwig 2011, 92, 205–232. [Google Scholar] [CrossRef]
- Ruwer, D.T.; Blanco, S.; Rodrigues, L. Eunotia Ehrenberg (Eunotiaceae, Bacillariophyta) in a subtropical floodplain: A new species and taxonomic contributions. Phytotaxa 2021, 505, 157–175. [Google Scholar] [CrossRef]
- Round, F.E.; Bukhtiyarova, L. Four new genera based on Achnanthes (Achnanthidium) together with a re-definition of Achnanthidium. Diatom Res. 1996, 11, 345–361. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2021. Available online: https://www.algaebase.org (accessed on 1 October 2021).
- Kociolek, J.P.; Blanco, S.; Coste, M.; Ector, L.; Liu, Y.; Karthick, B.; Kulikovskiy, M.; Lundholm, N.; Ludwig, T.V.; Potapova, M.; et al. DiatomBase. Available online: http://www.diatombase.org (accessed on 27 October 2021).
- Jahn, R.; Abarca, N.; Gemeinholzer, B.; Mora, D.; Skibbe, O.; Kulikovskiy, M.; Gusev, E.; Kusber, W.-H.; Zimmermann, J. Planothidium lanceolatum and Planothidium frequentissimum reinvestigated with molecular methods and morphology: Four new species and the taxonomic importance of the sinus and cavum. Diatom Res. 2017, 32, 75–107. [Google Scholar] [CrossRef]
- Stancheva, R. Planothidium sheathii, a new monoraphid diatom species from rivers in California, USA. Phytotaxa 2019, 393, 131–140. [Google Scholar] [CrossRef]
- Wetzel, C.E.; Van de Vijver, B.; Blanco, S.; Ector, L. On some common and new cavum-bearing Planothidium (Bacillariophyta) species from freshwater. Fottea 2019, 19, 50–89. [Google Scholar] [CrossRef] [Green Version]
- Potapova, M.G. New species and combinations in monoraphid diatoms (family Achnanthidiaceae) from North America. Diatom Res. 2012, 27, 29–42. [Google Scholar] [CrossRef]
- Wetzel, C.E.; Van De Vijver, B.; Hoffmann, L.; Ector, L. Planothidium incuriatum sp. nov. a widely distributed diatom species (Bacillariophyta) and type analysis of Planothidium biporomum. Phytotaxa 2013, 138, 43–57. [Google Scholar] [CrossRef]
- Compère, P.; Riaux–Gobin, C. Diatomées de quelques biotopes marins, saumâtres et dulçaquicoles de Guinée (Afrique oc-cidentale) [Diatoms from some marine, brackish and freshwater biotopes of Guinea (West Africa)]. Syst. Geogr. Plants 2009, 79, 33–66. [Google Scholar]
- N’Guessan, K.R.; Wetzel, C.E.; Ector, L.; Coste, M.; Cocquyt, C.; Van de Vijver, B.; Yao, S.S.; Ouattara, A.; Kouamelan, E.P.; Tison-Rosebery, J. Planothidium comperei sp. nov. (Bacillariophyta), a new diatom species from Ivory Coast. Plant Ecol. Evol. 2014, 147, 455–462. [Google Scholar] [CrossRef]
- Blanco, S.; Álvarez–Blanco, I.; Cejudo–Figueiras, C.; Recio Espejo, J.M.; Borja Barrera, C.; Bécares, E.; Díaz del Olmo, F.; Cámara Artigas, R. The diatom flora in temporary ponds of Doñana National Park (southwest Spain): Five new taxa. Nord. J. Bot. 2013, 31, 489–499. [Google Scholar] [CrossRef]
- Lai, G.G.; Ector, L.; Padedda, B.M.; Wetzel, C.E. Planothidium marganaiensis sp. nov. (Bacillariophyta), a new cavum-bearing species from a karst spring in south-western Sardinia (Italy). Phytotaxa 2021, 489, 140–154. [Google Scholar] [CrossRef]
- Kulikovskiy, M.S.; Lange–Bertalot, H.; Kuznetsova, I.V. Lake Baikal: Hotspot of endemic diatoms, 2. Iconogr. Diatomol. 2015, 26, 1–656. [Google Scholar]
- Van de Vijver, B.; Wetzel, C.; Kopalová, K.; Zidarova, R.; Ector, L. Analysis of the type material of Achnanthidium lanceolatum Brébisson ex Kützing (Bacillariophyta) with the description of two new Planothidium species from the Antarctic Region. Fottea 2013, 13, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, C.E.; Ector, L. Planothidium lagerheimii comb. nov. (Bacillariophyta, Achnanthales) a forgotten diatom from South America. Phytotaxa 2014, 188, 261. [Google Scholar] [CrossRef]
- Riaux-Gobin, C.; Witkowski, A.; Igersheim, A.; Lobban, C.S.; Al-Handal, A.Y.; Compère, P. Planothidium juandenovense sp. nov. (Bacillariophyta) from Juan de Nova (Scattered Islands, Mozambique Channel) and other tropical environments: A new addition to the Planothidium delicatulum complex. Fottea 2018, 18, 106–119. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.T.; Steinitz-Kannan, M.; Kallmeyer, D.E. Bacillariophyta: The Diatoms. In Algae: Source of Treatment; Ripley, M.G., Ed.; American Water Works Association: Denver, CO, USA, 2010; pp. 207–249. [Google Scholar]
- Stancheva, R.; Kristan, N.V.; Iii, W.B.K.; Sheath, R.G. Diatom genus Planothidium (Bacillariophyta) from streams and rivers in California, USA: Diversity, distribution and autecology. Phytotaxa 2020, 470, 1–30. [Google Scholar] [CrossRef]
- Bąk, M.; Halabowski, D.; Kryk, A.; Lewin, I.; Sowa, A. Mining salinisation of rivers: Its impact on diatom (Bacillariophyta) assemblages. Fottea 2020, 20, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Halabowski, D.; Bąk, M.; Lewin, U. Distribution and ecology of two interesting diatom species Navicula flandriae Van de Vijver et Mertens and Planothidium nanum Bąk, Kryk et Halabowski in rivers of Southern Poland and their spring areas. Oceanol. Hydrobiol. Stud. 2021, 50, 137–149. [Google Scholar] [CrossRef]
- Wynne, M.J. Delicatophycus gen. nov.: A validation of “Delicata Krammer” inval. (Gomphonemataceae, Bacillariophyta). Not. Algarum 2019, 97, 1–3. [Google Scholar]
- Krammer, K. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. In Diatoms of Europe, Diatoms of the Euro-Pean Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag, K.G.: Rugell, Liechtenstein, 2003; Volume 4, pp. 1–529. [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress, Shenzhen, China, 23–29 July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018; Volume 159, pp. 1–253. [Google Scholar]
- Cohu, R.L.; Lange-Bertalot, H.; Van de Viver, B.; Tudesque, L. Analysis and critical evaluation of structural features in four Cymbellaceae taxa from New Caledonia. Fottea 2020, 20, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.X. Flora algarum Sinicarum Aquae Dulcis, Vol. XVI, Bacillariophyta Cymbellaceae; Science Press: Beijing, China, 2013; pp. 1–217. [Google Scholar]
- Liu, B.; Blanco, S.; Qing-Yan, L. Ultrastructure of Delicata sinensis Krammer et Metzeltin and D. williamsii sp. nov. (Bacillariophyta) from China. Fottea 2018, 18, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Metzeltin, D.; Lange-Bertalot, H. Tropical Diatoms of South America II. Special remarks on biogeography disjunction. Iconogr. Diatomol. 2007, 18, 1–877. [Google Scholar]
- Bahls, L.L. Diatoms from Western North America 1. Some New and Notable Biraphid Species; Edited and Published by the Author: Helena, MT, USA, Montana Diatom Collection; 2017; pp. 1–52. [Google Scholar]
- Liu, W.; Li, Y.-L.; Wu, H.; Kociolek, J.P. Delicatophycus liuweii sp. nov., a new cymbelloid diatom (Bacillariophyceae) from an upper tributary of the Liujiang River, Guangxi, China. Phytotaxa 2021, 505, 63–70. [Google Scholar] [CrossRef]
- EN 13946. Water Quality. Guidance Standard for the Routine Sampling and Pretreatment of Benthic Diatoms from Rivers. 2003. Available online: http://http://www.safrass.com/partners_area/BSI%20Benthic%20diatoms.pdf (accessed on 1 October 2021).
- Cox, E.J. Identification of Freshwater Diatoms from Live Material; Chapman and Hall: London, UK, 1996; p. 158. [Google Scholar]
- Team, R.R. A language and Environment for Statistical Computing, Vienna, R Foundation for Statistical Computing. 2009. Available online: http://www.R-project.org (accessed on 27 October 2021).
- Rumrich, U.; Lange-Bertalot, H.; Rumrich, M. Diatomeen der Anden von Venezuela bis Patagonien/Feuerland und zwei weitere Beiträge. Diatoms of the Andes from Venezuela to Patagonia/Tierra del Fuego and two additional contributions. Iconogr. Diatomol. 2009, 9, 1–673. [Google Scholar]
- Sonneman, J.A.; Sincock, A.; Fluin, J.; Reid, M.A.; Newall, P.; Tibby, J.; Gell, P. All Illustrated Guide to Common Stream Di-atom Species from Temperate Australia; Cooperative Research Centre for Freshwater Ecology: Thurgoona, Australia, 2000; pp. 1–168. [Google Scholar]
- Dorofeyuk, N.I.; Kulikovskiy, M.S. Diatoms of Mongolia; Inst. Ecol. Evol. RAN: Moscow, Russia, 2012; pp. 1–367. [Google Scholar]
- Metzeltin, D.; Lange-Bertalot, H.; Soninkhishig, N. Diatoms in Mongolia. Icon. Diatom. 2009, 20, 1–686. [Google Scholar]
- Blanco, S.; Cejudo-Figueiras, C.; Álvarez-Blanco, I.; Bécares, E.; Hoffmann, L.; Ector, L. Atlas de las Diatomeas de la cuenca del Duero. Diatom atlas of the Duero Basin; Universidad de León: León, Spain, 2010; pp. 1–382. [Google Scholar]
- Wojtal, A.Z.; Łukasz, S. The influence of substrates and physicochemical factors on the composition of diatom assemblages in karst springs and their applicability in water-quality assessment. Hydrobiol 2012, 695, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Van de Vijver, B.; Frenot, Y.; Beyens, L. Freshwater diatoms from Ile de la Possession (Crozet Archipelago, Subantarctica). Biblioth. Diatomol. 2002, 46, 1–412. [Google Scholar]
- Lange-Bertalot, H.; Krammer, K. Achnanthes Eine Monographie der Gattung. Biblioth. Diatomol. 1989, 18, 1–393. [Google Scholar]
- Geitler, L. Beiträge zur Entwicklungsgeschichte und Taxonomie einiger Achnanthes-Arten, Subgenus Microneis (Bacillariophyceae). Plant Syst. Evol. 1980, 134, 1–10. [Google Scholar] [CrossRef]
- Reichardt, E. Die Diatomeen der Altmühl (Beitragë zur Diatomeenflora der Altmühl 2). Biblioth. Diatomol. 1984, 6, 1–169. [Google Scholar]
- Zechmeister, H.; Mucina, L. Vegetation of European springs: High-rank syntaxa of the Montio-Cardaminetea. J. Veg. Sci. 1994, 5, 385–402. [Google Scholar] [CrossRef]
- Cantonati, M.; Kelly, M.G.; Lange-Bertalot, H. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Koeltz Botanical Books: Schmitten-Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Rogora, M.; Frate, L.; Carranza, M.L.; Freppaz, M.; Stanisci, A.; Bertani, I.; Bottarin, R.; Brambilla, A.; Canullo, R.; Carbognani, M.; et al. Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci. Total. Environ. 2018, 624, 1429–1442. [Google Scholar] [CrossRef] [Green Version]
- Bahls, L.; Pierce, J.; Apfelbeck, R.; Olsen, L. Encyonema droseraphilum sp. nov. (Bacillariophyta) and other rare diatoms from undisturbed floating-mat fens in the northern Rocky Mountains, USA. Phytotaxa 2013, 127, 32. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
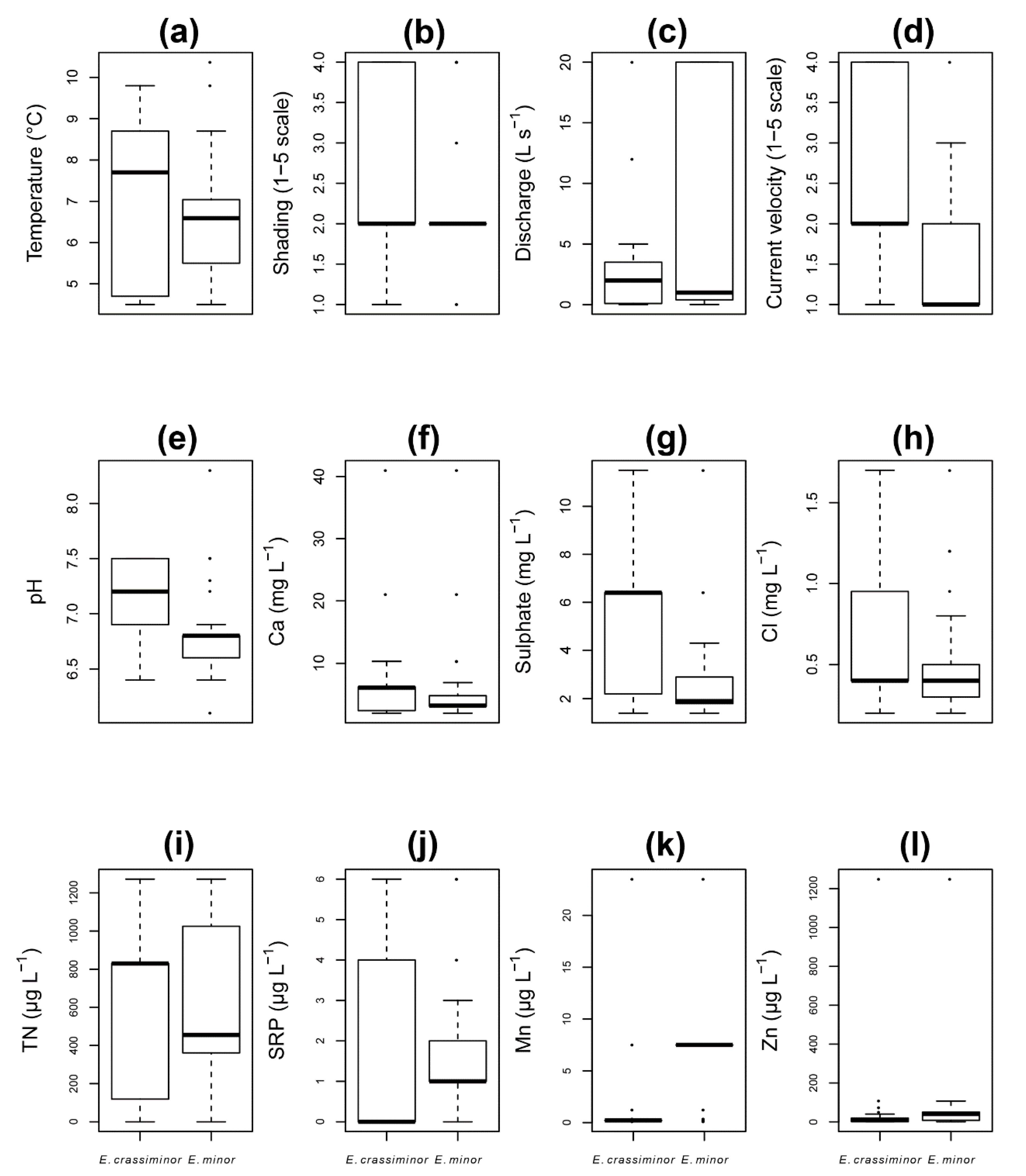
| Eunotia crassiminor | Eunotia minor | t-Tests | ||||||
|---|---|---|---|---|---|---|---|---|
| Factor | Weighted Average | Min | Max | Weighted Average | Min | Max | t-Value | p-Value |
| Temperature (°C) | 5.6 | 4.5 | 9.8 | 7.8 | 4.5 | 8.7 | 7.58 | 5.99 × 10−11 |
| Shading (1–5 scale) | 3 | 1 | 4 | 2 | 2 | 2 | −9.08 | 1.20 × 10−14 |
| Discharge (L s−1) | 2.0 | 0.0 | 5.0 | 4.3 | 0.0 | 20.0 | 2.0 | 4.85 × 10−2 |
| Current velocity (1–5 scale) | 3 | 1 | 4 | 2 | 1 | 3 | −6.07 | 2.33 × 10−8 |
| pH | 6.8 | 6.4 | 7.5 | 7.0 | 6.4 | 6.9 | 2.8 | 6.11 × 10−3 |
| Ca2+ (mg L−1) | 4.9 | 2.0 | 10.3 | 8.5 | 2.0 | 6.9 | 2.20 | 3.11 × 10−2 |
| Sulphates (mg L−1) | 5.1 | 1.4 | 11.5 | 2.2 | 1.4 | 4.3 | −7.39 | 1.45 × 10−11 |
| Cl−1 (mg L−1) | 0.4 | 0.2 | 1.7 | 0.8 | 0.2 | 0.8 | 4.53 | 2.28 × 10−5 |
| TN (µg L−1) | 340 | 0 | 1272 | 649 | 0 | 1272 | 4.29 | 7.69 × 10−5 |
| SRP (µg L−1) | 2 | 0 | 6 | 2 | 0 | 3 | −2.25 | 2.69 × 10−2 |
| Mn (µg L−1) | 0.6 | 0.2 | 0.2 | 9.8 | 7.5 | 7.5 | 4.95 | 2.59 × 10−5 |
| Zn (µg L−1) | 101 | 0 | 40 | 37 | 0 | 107 | −2.12 | 3.56 × 10−2 |
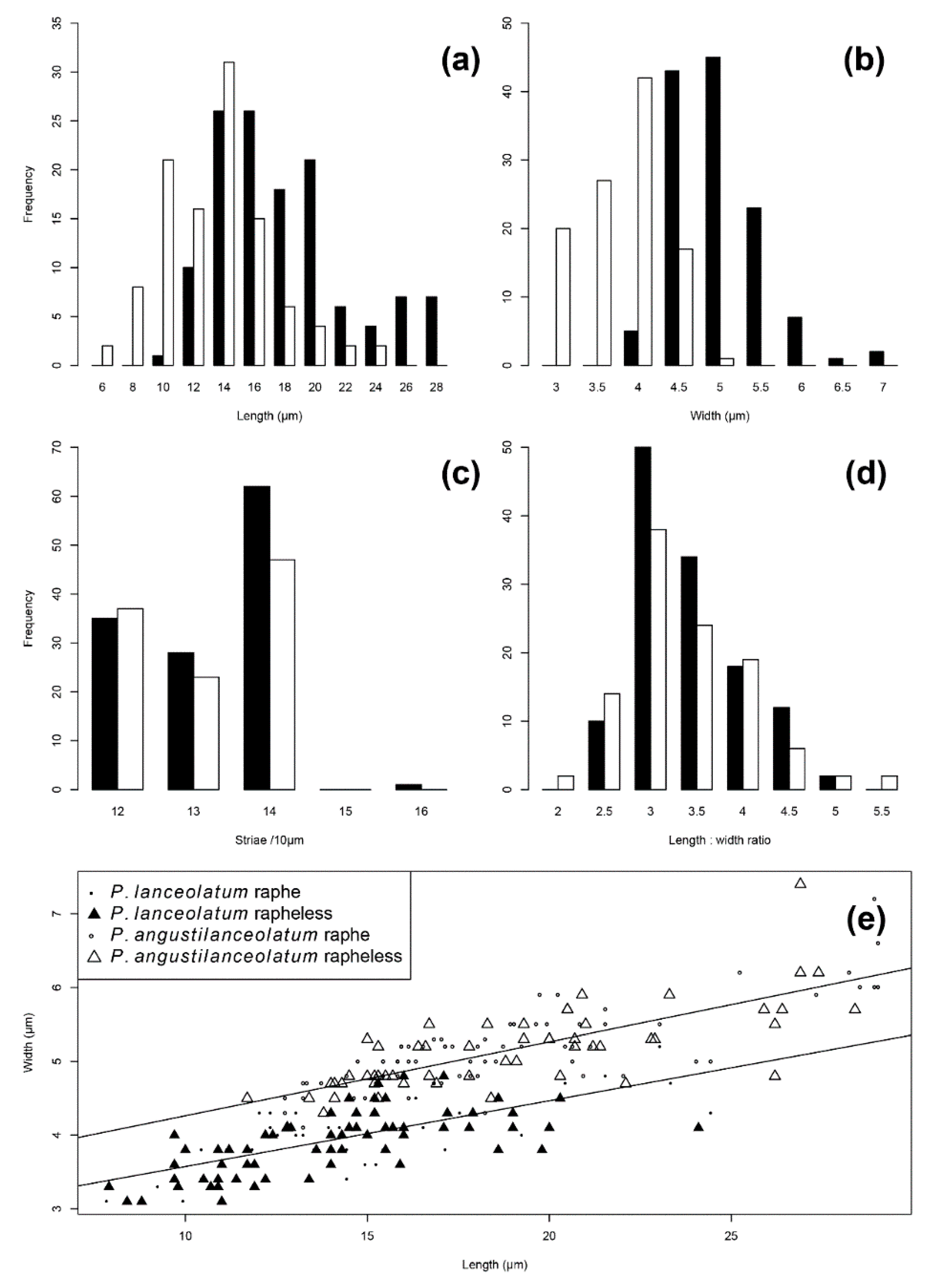
| P. angustilanceolatum | P. lanceolatum | t-Tests | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R-Valves | RL-Valves | R-Valves | RL-Valves | R-Valves | RL-Valves | |||||||||||||
| Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | t-Value | df | p-Value | t-Value | df | p-Value | |
| Length (µm) | 14.9 | 7.9 | 24.5 | 14.0 | 7.9 | 24.1 | 18.8 | 12.8 | 29.1 | 18.9 | 11.7 | 27.4 | 5.12 | 99 | 1.51 × 10−6 | 6.86 | 101 | 5.44 × 10−10 |
| Width (µm) | 4.1 | 3.1 | 5.2 | 3.9 | 3.1 | 4.8 | 5.2 | 4.1 | 7.2 | 5.1 | 4.1 | 7.4 | 11.09 | 98 | <2.2 × 10−16 | 13.41 | 100 | <2.2 × 10−16 |
| Length/Width | 3.6 | 2.6 | 5.7 | 3.6 | 2.4 | 5.8 | 3.6 | 2.8 | 4.9 | 3.7 | 2.6 | 5.4 | −0.09 | 75 | 9.28 × 10−1 | 0.78 | 117 | 4.38 × 10−1 |
| Striae in 10 µm | 13 | 12 | 14 | 13 | 12 | 14 | 13 | 12 | 16 | 13 | 12 | 14 | 1 | 88 | 1.84 × 10−1 | 0.21 | 114 | 8.37 × 10−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantonati, M.; Bilous, O.; Angeli, N.; van Wensen, L.; Lange-Bertalot, H. Three New Diatom Species from Spring Habitats in the Northern Apennines (Emilia-Romagna, Italy). Diversity 2021, 13, 549. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110549
Cantonati M, Bilous O, Angeli N, van Wensen L, Lange-Bertalot H. Three New Diatom Species from Spring Habitats in the Northern Apennines (Emilia-Romagna, Italy). Diversity. 2021; 13(11):549. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110549
Chicago/Turabian StyleCantonati, Marco, Olena Bilous, Nicola Angeli, Liesbeth van Wensen, and Horst Lange-Bertalot. 2021. "Three New Diatom Species from Spring Habitats in the Northern Apennines (Emilia-Romagna, Italy)" Diversity 13, no. 11: 549. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110549