
Relationship between Nutrient Intake and Human Gut Microbiota in Monozygotic Twins

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Nutrition Data Collection
2.3. Fecal Sample Collection
2.4. DNA Extraction and 16S rRNA Gene Amplicon Sequencing
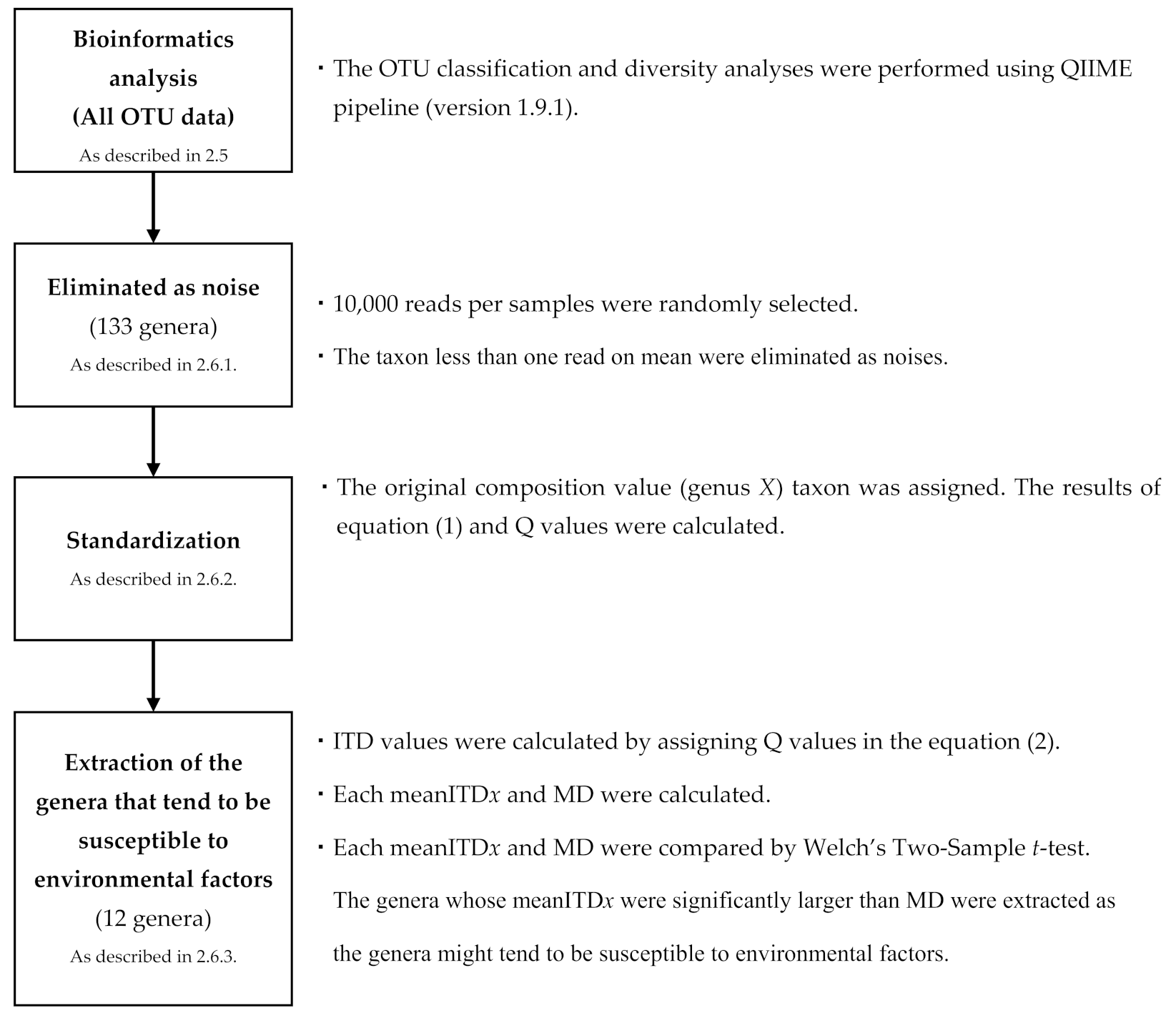
2.5. Bioinformatics Analysis
2.6. Statistical Analyses
2.6.1. Noise Processing
2.6.2. Standardization
2.6.3. Extraction of the Susceptible Genera to Environmental Factors
2.6.4. Correlation between the Target Genera and Nutrient Intake
3. Results
3.1. Characteristics of Participants
3.2. Susceptible Genera to Environmental Factors
3.3. Association of Specific Genera with Nutrient Intake
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ITD | Intra-twins difference |
| mean ITDx | the mean ITD values of genus X for each of the 133 genera |
| MD | the mean of ITD values of 28 pairs for all 133 genera |
| ITND | Intra-twins nutrient difference |
References
- Kamada, N.; Seo, S.-U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Caviglia, G.P.; Rosso, C.; Ribaldone, D.G.; Dughera, F.; Fagoonee, S.; Astegiano, M.; Pellicano, R. Physiopathology of intestinal barrier and the role of zonulin. Minerva Biotecnol. 2019, 31, 83–92. [Google Scholar] [CrossRef]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, S.J.D.; Li, J.V.; Lahti, L.; Ou, J.; Carbonero, F.; Mohammed, K.; Posma, J.M.; Kinross, J.; Wahl, E.; Ruder, E.; et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 2015, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.S.; Koller, K.R.; Ramaboli, M.C.; Nesengani, L.T.; Ocvirk, S.; Chen, C.; Flanagan, C.A.; Sapp, F.R.; Merritt, Z.T.; Bhatti, F.; et al. Diet and the Human Gut Microbiome: An International Review. Dig. Dis. Sci. 2020, 65, 723–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marco, M.L.; Hill, C.; Hutkins, R.; Slavin, J.; Tancredi, D.J.; Merenstein, D.; Sanders, M.E. Should There Be a Recommended Daily Intake of Microbes? J. Nutr. 2020, 150, 3061–3067. [Google Scholar] [CrossRef] [PubMed]
- Mohr, A.E.; Jäger, R.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Townsend, J.R.; West, N.P.; Black, K.; Gleeson, M.; Pyne, D.B.; et al. The athletic gut microbiota. J. Int. Soc. Sports Nutr. 2020, 17, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Useros, N.; Nova, E.; González-Zancada, N.; Díaz, L.E.; Gómez-Martínez, S.; Marcos, A. Microbiota and Lifestyle: A Special Focus on Diet. Nutrients 2020, 12, 1776. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Davenport, E.R.; Beaumont, M.; Jackson, M.A.; Knight, R.; Ober, C.; Spector, T.D.; Bell, J.T.; Clark, A.G.; Ley, R.E. Genetic Determinants of the Gut Microbiome in UK Twins. Cell Host Microbe 2016, 19, 731–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awany, D.; Allali, I.; Dalvie, S.; Hemmings, S.; Mwaikono, K.S.; Thomford, N.E.; Gomez, A.; Mulder, N.; Chimusa, E.R. Host and Microbiome Genome-Wide Association Studies: Current State and Challenges. Front. Genet. 2019, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Peng, W.; Zhou, Y.; Ren, Y.; Zhao, J.; Fu, X.; Nie, Y. Host Genetic and Environmental Factors Shape the Composition and Function of Gut Microbiota in Populations Living at High Altitude. BioMed Res. Int. 2020, 2020, 1482109. [Google Scholar] [CrossRef] [PubMed]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.-Z.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Gallego, J.L.; Chou, S.-P.; Di Rienzi, S.C.; Goodrich, J.K.; Spector, T.D.; Bell, J.T.; Youngblut, N.D.; Hewson, I.; Reyes, A.; Ley, R.E. Virome Diversity Correlates with Intestinal Microbiome Diversity in Adult Monozygotic Twins. Cell Host Microbe 2019, 25, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-L.; Segovia, I.; Yuan, X.-L.; Gao, Z.-H. Controversial Roles of Gut Microbiota-Derived Short-Chain Fatty Acids (SCFAs) on Pancreatic β-Cell Growth and Insulin Secretion. Int. J. Mol. Sci. 2020, 21, 910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leccioli, V.; Oliveri, M.; Romeo, M.; Berretta, M.; Rossi, P. A New Proposal for the Pathogenic Mechanism of Non-Coeliac/Non-Allergic Gluten/Wheat Sensitivity: Piecing Together the Puzzle of Recent Scientific Evidence. Nutrients 2017, 9, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakkas, H.; Bozidis, P.; Touzios, C.; Kolios, D.; Athanasiou, G.; Athanasopoulou, E.; Gerou, I.; Gartzonika, C. Nutritional Status and the Influence of the Vegan Diet on the Gut Microbiota and Human Health. Medicine 2020, 56, 88. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tap, J.; Furet, J.; Bensaada, M.; Philippe, C.; Roth, H.; Rabot, S.; Lakhdari, O.; Lombard, V.; Henrissat, B.; Corthier, G.; et al. Gut microbiota richness promotes its stability upon increased dietary fibre intake in healthy adults. Environ. Microbiol. 2015, 17, 4954–4964. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Nishijima, S.; Suda, W.; Oshima, K.; Kim, S.-W.; Hirose, Y.; Morita, H.; Hattori, M. The gut microbiome of healthy Japanese and its microbial and functional uniqueness. DNA Res. 2016, 23, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, S.; Murakami, K.; Sasaki, S.; Okubo, H.; Hirota, N.; Notsu, A.; Fukui, M.; Date, C. Comparison of relative validity of food group intakes estimated by comprehensive and brief-type self-administered diet history questionnaires against 16 d dietary records in Japanese adults. Public Health Nutr. 2011, 14, 1200–1211. [Google Scholar] [CrossRef]
- Hosomi, K.; Ohno, H.; Murakami, H.; Natsume-Kitatani, Y.; Tanisawa, K.; Hirata, S.; Suzuki, H.; Nagatake, T.; Nishino, T.; Mizuguchi, K.; et al. Method for preparing DNA from feces in guanidine thiocyanate solution affects 16S rRNA-based profiling of human microbiota diversity. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Mohsen, A.; Park, J.; Chen, Y.A.; Kawashima, H.; Mizuguchi, K. Impact of quality trimming on the efficiency of reads joining and diversity analysis of Illumina Paired-end reads in the context of QIIME1 and QIIME2 microbiome analysis frameworks. BMC Bioinform. 2019, 20, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Zar, J.H. Significance Testing of the Spearman Rank Correlation Coefficient. J. Am. Stat. Assoc. 1972, 67, 578–580. [Google Scholar] [CrossRef]
- Sun, C.L.; Relman, D.A. Microbiota’s ’little helpers’: Bacteriophages and antibiotic-associated responses in the gut microbiome. Genome Biol. 2013, 14, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theriot, C.M.; Koenigsknecht, M.J.; Carlson, P.E.; Hatton, G.E.; Nelson, A.M.; Li, B.; Huffnagle, G.B.; Li, J.Z.; Young, V.B. Antibiotic-induced shifts in the mouse gut microbiome and metabolome increase susceptibility to Clostridium difficile infection. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Francino, M.P. Antibiotics and the Human Gut Microbiome: Dysbioses and Accumulation of Resistances. Front. Microbiol. 2016, 6, 1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purohit, V.; Bode, J.C.; Bode, C.; Brenner, D.A.; Choudhry, M.A.; Hamilton, F.; Kang, Y.J.; Keshavarzian, A.; Rao, R.; Sartor, R.B.; et al. Alcohol, intestinal bacterial growth, intestinal permeability to endotoxin, and medical consequences: Summary of a symposium. Alcohol 2008, 42, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Bull-Otterson, L.; Feng, W.; Kirpich, I.; Wang, Y.; Qin, X.; Liu, Y.; Gobejishvili, L.; Joshi-Barve, S.; Ayvaz, T.; Petrosino, J.; et al. Metagenomic Analyses of Alcohol Induced Pathogenic Alterations in the Intestinal Microbiome and the Effect of Lactobacillus rhamnosus GG Treatment. PLoS ONE 2013, 8, e53028. [Google Scholar] [CrossRef]
- Bergström, A.; Skov, T.H.; Bahl, M.I.; Roager, H.M.; Christensen, L.B.; Ejlerskov, K.T.; Mølgaard, C.; Michaelsen, K.F.; Licht, T.R. Establishment of Intestinal Microbiota during Early Life: A Longitudinal, Explorative Study of a Large Cohort of Danish Infants. Appl. Environ. Microbiol. 2014, 80, 2889–2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tang, H.; Zhang, C.; Zhao, Y.; Derrien, M.; Rocher, E.; Vlieg, J.E.T.V.-H.; Strissel, K.J.; Zhao, L.; Obin, M.S.; et al. Modulation of gut microbiota during probiotic-mediated attenuation of metabolic syndrome in high fat diet-fed mice. ISME J. 2015, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nirmalkar, K.; Murugesan, S.; Pizano-Zárate, M.L.; Villalobos-Flores, L.E.; García-González, C.; Morales-Hernández, R.M.; Nuñez-Hernández, J.A.; Hernández-Quiroz, F.; Romero-Figueroa, M.D.S.; Hernández-Guerrero, C.; et al. Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents. Nutrients 2018, 10, 2009. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, M.Y.; You, H.J.; Yoon, H.S.; Kwon, B.; Lee, J.Y.; Lee, S.; Song, Y.-M.; Lee, K.; Sung, J.; Ko, G. The effect of heritability and host genetics on the gut microbiota and metabolic syndrome. Gut 2016, 66, 1031–1038. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Bagarolli, R.A.; Tobar, N.; Oliveira, A.G.; Araújo, T.G.; Carvalho, B.M.; Rocha, G.Z.; Vecina, J.F.; Calisto, K.; Guadagnini, D.; Prada, P.O.; et al. Probiotics modulate gut microbiota and improve insulin sensitivity in DIO mice. J. Nutr. Biochem. 2017, 50, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Juntao, H.; Ning, G.; Xiaolan, L.; Ming, L.; Xiaokang, W.; Zeshi, L.; Yaofeng, J.; Jiali, L.; Jiru, X.; et al. Probiotics may delay the progression of nonalcoholic fatty liver disease by restoring the gut microbiota structure and improving intestinal endotoxemia. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-S.; Choi, J.W.; Jhun, J.; Kwon, J.Y.; Lee, B.-I.; Yang, C.W.; Park, S.-H.; Cho, M.-L. Lactobacillus acidophilus Improves Intestinal Inflammation in an Acute Colitis Mouse Model by Regulation of Th17 and Treg Cell Balance and Fibrosis Development. J. Med. Food 2018, 21, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulik, C.M.; Sullivan, P.F.; Kendler, K.S. Genetic and environmental contributions to obesity and binge eating. Int. J. Eat. Disord. 2003, 33, 293–298. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Cardoneanu, A.; Mihai, C.; Rezus, E.; Burlui, A.; Popa, I.; Prelipcean, C.C. Gut microbiota changes in inflammatory bowel diseases and ankylosing spondilytis. J. Gastrointest. Liver Dis. 2021. [Google Scholar] [CrossRef]
- Elhag, D.A.; Kumar, M.; Al Khodor, S. Exploring the Triple Interaction between the Host Genome, the Epigenome, and the Gut Microbiome in Type 1 Diabetes. Int. J. Mol. Sci. 2020, 22, 125. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, H.; Eida, T.; Takizawa, T.; Tokunaga, T.; Tashiro, Y. Effects of Fructooligosaccharides on Intestinal Flora and Human Health. Bifidobact. Microflora 1986, 5, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Li, Q.; Hu, K.; He, Y.; Ai, Q.; Hu, L.; Yu, J. Vitamin A and Retinoic Acid Exhibit Protective Effects on Necrotizing Enterocolitis by Regulating Intestinal Flora and Enhancing the Intestinal Epithelial Barrier. Arch. Med. Res. 2018, 49, 1–9. [Google Scholar] [CrossRef]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pataky, Z.; Genton, L.; Spahr, L.F.J.; Lazarevic, V.; Terraz, S.; Gaïa, N.; Rubbia-Brandt, L.; Golay, A.; Schrenzel, J.; Pichard, C. Impact of Hypocaloric Hyperproteic Diet on Gut Microbiota in Overweight or Obese Patients with Nonalcoholic Fatty Liver Disease: A Pilot Study. Dig. Dis. Sci. 2016, 61, 2721–2731. [Google Scholar] [CrossRef] [PubMed]
- Smith-Brown, P.; Morrison, M.; Krause, L.; Davies, P.S.W. Dairy and plant based food intakes are associated with altered faecal microbiota in 2 to 3 year old Australian children. Sci. Rep. 2016, 6, 32385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mompeo, O.; Spector, T.D.; Hernandez, M.M.; Le Roy, C.; Istas, G.; Le Sayec, M.; Mangino, M.; Jennings, A.; Rodriguez-Mateos, A.; Valdes, A.M.; et al. Consumption of Stilbenes and Flavonoids is Linked to Reduced Risk of Obesity Independently of Fiber Intake. Nutrients 2020, 12, 1871. [Google Scholar] [CrossRef]
- Yang, C.; Deng, Q.; Xu, J.; Wang, X.; Hu, C.; Tang, H.; Huang, F. Sinapic acid and resveratrol alleviate oxidative stress with modulation of gut microbiota in high-fat diet-fed rats. Food Res. Int. 2019, 116, 1202–1211. [Google Scholar] [CrossRef]
- Joffe, Y.T.; Collins, M.; Goedecke, J.H. The Relationship between Dietary Fatty Acids and Inflammatory Genes on the Obese Phenotype and Serum Lipids. Nutrients 2013, 5, 1672–1705. [Google Scholar] [CrossRef] [Green Version]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [Green Version]
- Udayappan, S.; Manneras-Holm, L.; Chaplin-Scott, A.; Belzer, C.; Herrema, H.; Dallinga-Thie, G.M.; Duncan, S.H.; Stroes, E.S.G.; Groen, A.K.; Flint, H.J.; et al. Oral treatment with Eubacterium hallii improves insulin sensitivity in db/db mice. NPJ Biofilms Microbiomes 2016, 2, 16009. [Google Scholar] [CrossRef] [Green Version]
- Meehan, C.J.; Beiko, R.G. A Phylogenomic View of Ecological Specialization in the Lachnospiraceae, a Family of Digestive Tract-Associated Bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Hiippala, K.; Jouhten, H.; Ronkainen, A.; Hartikainen, A.; Kainulainen, V.; Jalanka, J.; Satokari, R. The Potential of Gut Commensals in Reinforcing Intestinal Barrier Function and Alleviating Inflammation. Nutrients 2018, 10, 988. [Google Scholar] [CrossRef] [Green Version]
- Munukka, E.; Rintala, A.; Toivonen, R.; Nylund, M.; Yang, B.; Takanen, A.; Hänninen, A.; Vuopio, J.; Huovinen, P.; Jalkanen, S.; et al. Faecalibacterium prausnitzii treatment improves hepatic health and reduces adipose tissue inflammation in high-fat fed mice. ISME J. 2017, 11, 1667–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]
- Vamanu, E.; Gatea, F. Correlations between Microbiota Bioactivity and Bioavailability of Functional Compounds: A Mini-Review. Biomedicine 2020, 8, 39. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Mean | ± | SD | (Min | – | Max) | Median | ||
|---|---|---|---|---|---|---|---|---|
| Age | 59.3 | ± | 19.4 | 20 | – | 80 | 67.5 | |
| Body mass index | 23.4 | ± | 3.9 | 15.5 | – | 32.9 | 23.8 | |
| Nutrients | ||||||||
| Energy intake | (kcal/day) | 2057.5 | ± | 587.4 | 858.9 | – | 3564.9 | 1946.5 |
| Protein (PRT) | (g/1000 kcal) | 38.5 | ± | 7.4 | 20.3 | – | 54.6 | 37.3 |
| Fat (FAT) | (g/1000 kcal) | 30.2 | ± | 6.6 | 18.5 | – | 46.1 | 30.2 |
| Saturated Fatty Acid (SFA) | (g/1000 kcal) | 7.9 | ± | 2.1 | 4.6 | – | 14.0 | 7.6 |
| Carbohydrates (CHO) | (g/1000 kcal) | 133.9 | ± | 19.7 | 94.7 | – | 172.0 | 135.3 |
| Total Dietary Fiber (TDF) | (g/1000 kcal) | 6.7 | ± | 1.8 | 2.5 | – | 10.2 | 6.8 |
| Sodium (Na) | (mg/1000 kcal) | 2307.0 | ± | 479.2 | 1300.8 | – | 3630.0 | 2271.6 |
| Potassium (K) | (mg/1000 kcal) | 1459.7 | ± | 429.4 | 564.2 | – | 2307.9 | 1412.4 |
| Calcium (Ca) | (mg/1000 kcal) | 319.4 | ± | 122.5 | 91.9 | – | 625.7 | 310.7 |
| Iron (Fe) | (mg/1000 kcal) | 4.5 | ± | 1.1 | 2.2 | – | 6.7 | 4.7 |
| Retinol equivalent (RTE) | (µg/1000 kcal) | 434.9 | ± | 197.6 | 98.2 | – | 907.4 | 390.2 |
| Vitamin D (VD) | (µg/1000 kcal) | 7.7 | ± | 4.6 | 0.2 | – | 23.2 | 6.6 |
| Alpha-tocopherol (ATC) | (µg/1000 kcal) | 4.2 | ± | 1.2 | 2.1 | – | 8.1 | 4.3 |
| Vitamin K (VK) | (µg/1000 kcal) | 192.1 | ± | 91.7 | 33.9 | – | 441.9 | 189.9 |
| Vitamin B1 (VB1) | (mg/1000 kcal) | 0.4 | ± | 0.1 | 0.2 | – | 0.7 | 0.4 |
| Vitamin B2 (VB2) | (mg/1000 kcal) | 0.7 | ± | 0.2 | 0.3 | – | 1.3 | 0.7 |
| Vitamin B6 (VB6) | (mg/1000 kcal) | 0.7 | ± | 0.2 | 0.3 | – | 1.6 | 0.7 |
| Vitamin B12 (VB12) | (µg/1000 kcal) | 4.9 | ± | 2.6 | 0.3 | – | 14.2 | 4.3 |
| Vitamin C (VC) | (mg/1000 kcal) | 64.6 | ± | 28.1 | 17.6 | – | 151.3 | 60.2 |
| Family | Genus | The Mean Intra-Twin Differences (Mean ITDx) | p Value | |
|---|---|---|---|---|
| ALL | 0.668 | ― | ||
| Lactobacillaceae | Lactobacillus | 0.380 | 0.015 | * |
| Bacteroidaceae | Bacteroides | 1.338 | 0.000 | * |
| Bifidobacteriaceae | Gardnerella | 1.073 | 0.035 | * |
| Lachnospiraceae | Lachnospiraceae UCG-008 group | 1.163 | 0.006 | * |
| Lachnospiraceae | Lachnospiraceae UCG-004 group | 1.075 | 0.018 | * |
| Lachnospiraceae | Lachnospiraceae ND3007 group | 1.063 | 0.022 | * |
| Lachnospiraceae | Lachnospiraceae FCS020 group | 1.060 | 0.026 | * |
| Lachnospiraceae | Roseburia | 1.146 | 0.020 | * |
| Lachnospiraceae | Eubacterium hallii group | 0.987 | 0.032 | * |
| Lachnospiraceae | Lachnospira | 1.037 | 0.041 | * |
| Porphyromonadaceae | Parabacteroides | 1.064 | 0.028 | * |
| Ruminococcaceae | Faecalibacterium | 1.043 | 0.028 | * |
| Ruminococcaceae | Ruminococcaceae UCG-003 group | 1.030 | 0.038 | * |
| Family | Genus | Protein | Fat | Carbohydrate | Mineral | Vitamin | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PRT | FAT | SFA | CHO | TDF | NA. | K | CA | FE | RTE | VD | ATC | VK | VB1 | VB2 | VB6 | VB12 | VC | ||
| Bacteroidaceae | Bacteroides | 0.11 | −0.10 | 0.00 | 0.10 | 0.01 | −0.12 | −0.05 | 0.27 | −0.06 | 0.38 * | 0.12 | −0.03 | −0.07 | −0.10 | 0.13 | 0.00 | 0.14 | −0.14 |
| Bifidobacteriaceae | Gardnerella | 0.13 | 0.02 | 0.07 | −0.20 | −0.20 | 0.11 | −0.14 | −0.15 | −0.21 | −0.23 | 0.19 | −0.24 | −0.23 | −0.10 | −0.15 | 0.01 | 0.17 | −0.33 |
| Porphyromonadaceae | Parabacteroides | 0.19 | −0.21 | 0.01 | −0.08 | 0.13 | 0.13 | 0.14 | 0.16 | 0.08 | −0.14 | 0.28 | 0.00 | −0.06 | 0.09 | 0.02 | 0.06 | 0.10 | 0.19 |
| Lachnospiraceae | Eubacterium hallii group | −0.10 | −0.06 | −0.02 | 0.08 | −0.15 | −0.17 | −0.05 | 0.07 | −0.10 | 0.33 | −0.21 | −0.13 | −0.18 | −0.14 | 0.10 | −0.24 * | −0.23 | −0.08 |
| Lachnospiraceae | Lachnospira | −0.38 * | 0.11 | 0.14 | 0.22 | −0.31 | −0.41 * | −0.37 | −0.20 | −0.39 * | 0.15 | −0.63 * | −0.32 | −0.25 | −0.36 | −0.10 | −0.42 | −0.49 * | −0.32 |
| Lachnospiraceae | Lachnospiraceae FCS020 group | −0.14 | −0.19 | −0.24 | 0.11 | −0.14 | −0.26 | −0.12 | −0.31 | −0.17 | −0.31 | −0.30 | −0.21 | 0.07 | −0.16 | −0.18 | −0.11 | −0.29 | 0.07 |
| Lachnospiraceae | Lachnospiraceae ND3007 group | 0.16 | 0.39 * | 0.32 | −0.28 | 0.25 | −0.04 | 0.12 | 0.24 | 0.16 | 0.02 | 0.00 | 0.15 | 0.32 | 0.22 | 0.12 | 0.11 | 0.03 | 0.16 |
| Lachnospiraceae | Lachnospiraceae UCG−004 | 0.23 | 0.13 | 0.20 | −0.25 | 0.14 | 0.09 | 0.15 | 0.22 | 0.14 | 0.06 | 0.16 | 0.07 | 0.18 | 0.15 | 0.21 | 0.17 | 0.13 | −0.02 |
| Lachnospiraceae | Lachnospiraceae UCG−008 | −0.08 | −0.32 | −0.45 * | 0.12 | −0.04 | −0.20 | 0.04 | −0.10 | −0.08 | 0.03 | 0.05 | 0.05 | −0.05 | −0.02 | −0.23 | −0.04 | −0.02 | 0.12 |
| Lachnospiraceae | Roseburia | 0.05 | −0.08 | −0.15 | 0.01 | −0.03 | −0.20 | −0.04 | −0.07 | −0.02 | 0.01 | −0.04 | −0.03 | −0.01 | 0.03 | −0.08 | −0.06 | −0.08 | 0.14 |
| Ruminococcaceae | Faecalibacterium | −0.16 | 0.00 | 0.07 | 0.16 | −0.14 | −0.10 | −0.09 | 0.00 | −0.14 | −0.14 | −0.26 | −0.17 | −0.30 | −0.17 | 0.02 | −0.30 | −0.31 | −0.02 |
| Ruminococcaceae | Ruminococcaceae UCG−003 | −0.03 | 0.17 | 0.16 | −0.18 | 0.09 | 0.03 | 0.14 | −0.02 | 0.10 | −0.18 | −0.13 | 0.04 | 0.27 | 0.17 | 0.10 | 0.15 | −0.11 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, N.; Park, J.; Tomizawa, R.; Kawashima, H.; Hosomi, K.; Mizuguchi, K.; Honda, C.; Ozaki, R.; Iwatani, Y.; Watanabe, M.; et al. Relationship between Nutrient Intake and Human Gut Microbiota in Monozygotic Twins. Medicina 2021, 57, 275. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57030275
Matsumoto N, Park J, Tomizawa R, Kawashima H, Hosomi K, Mizuguchi K, Honda C, Ozaki R, Iwatani Y, Watanabe M, et al. Relationship between Nutrient Intake and Human Gut Microbiota in Monozygotic Twins. Medicina. 2021; 57(3):275. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57030275
Chicago/Turabian StyleMatsumoto, Natsuko, Jonguk Park, Rie Tomizawa, Hitoshi Kawashima, Koji Hosomi, Kenji Mizuguchi, Chika Honda, Ritsuko Ozaki, Yoshinori Iwatani, Mikio Watanabe, and et al. 2021. "Relationship between Nutrient Intake and Human Gut Microbiota in Monozygotic Twins" Medicina 57, no. 3: 275. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57030275