1. Introduction
Insulin resistance (IR), a prediabetic state of type 2 diabetes, is an important health issue with a rapidly increasing incidence worldwide [
1,
2]. Several studies reported positive associations between particulate matters (PMs) and serum levels of fasting glucose, an important IR indicator [
3,
4]. The adverse effect of PM
10 with a diameter of less than 10 μm on fasting glucose level was proven in the general population, regardless of short-term or long-term exposure [
3,
4]. Although short-term or long-term exposure to PM
2.5 with a diameter of less than 2.5 μm was also reported to induce IR in C57BL/6 mice and incident metabolic syndrome in KORA cohort [
5,
6], evidence for the effects of PM
2.5 and particularly PM
2.5–10, known as coarse PM, on fasting glucose levels was insufficient.
Oxidative stress is considered to be a major biological mechanism underlying IR [
7]. Although a variety of potential mechanisms of PMs inducing IR have been suggested, oxidative stress is still at the center of research attention [
8,
9,
10,
11]. Exposure to PM
2.5 in mice can cause vascular IR by inducing pulmonary oxidative stress [
8,
12]. Furthermore, the adverse effect of PM
2.5 on vascular IR can be reduced by removing superoxide from the lungs of mice [
11,
12]. The water-insoluble fraction of PM
10 can also induce oxidative stress in human lung epithelial A549 cells [
13]. Because PMs contain pro-oxidant molecules, such as chromium, iron, nickel, and zinc, that can induce reactive oxygen species [
10], oxidative stress is plausible as a major mechanism of PMs on IR. Because oxidative stress could induce oxidative DNA damages, cells need to up-regulate DNA repair genes to protect against oxidative DNA damage upon PMs exposure [
14]. In fact, an epidemiologic study reported that PMs exposure could even induce oxidative stress-associated DNA damage in healthy young adults exposed to low concentrations of ambient PM
2.5–10 and PM
2.5 [
15].
Poly(ADP-ribose) polymerase family member 4 (PARP4) and excision repair cross-complementation 1 (ERCC1), respectively, repair DNA damage using base excision and nucleotide excision repair pathways to remove oxidized DNA bases or nucleotides [
16,
17]. These DNA damage repairs are considered to be very important processes because they can protect the human body against oxidative stress. It has been reported that PARP is activated by DNA strand breakage through the excessive accumulation of reactive oxygen species in relation to hyperglycemia [
18]. Although
PARP1 and
PARP2, included in the same family as
PARP4, were reported to affect glucose metabolism and insulin sensitivity [
19], there is no evidence for the effect of
PARP4 on blood glucose level. On the other hand,
ERCC1 gene was reported to have an impact on glucose intolerance in a progeroid mouse model with ERCC1 deficiency, resulting in fat loss and IR by triggering an autoinflammatory response [
20]. Previous evidence showed that genetic variations of
PARP4 and
ERCC1 could potentially affect IR differently by changing the capacity of corresponding enzymes encoded by them.
As population aging increases, we need to pay attention to the elderly, who are more vulnerable to chronic diseases than others [
21]. With increasing age, the body’s function as well as immune system become more sensitive and vulnerable [
21]. The EpiAir study, an epidemiologic surveillance on air pollution and Italian health, indicated that elderly subjects were more vulnerable to exposure to particulate matters than to other pollutants [
21]. Therefore, the objective of the present study was to evaluate relations between PMs (PM
10, PM
2.5–10, and PM
2.5) exposure and the serum level of fasting glucose among the elderly population considering modification by genetic polymorphisms of
PARP4 and
ERCC1.
4. Discussion
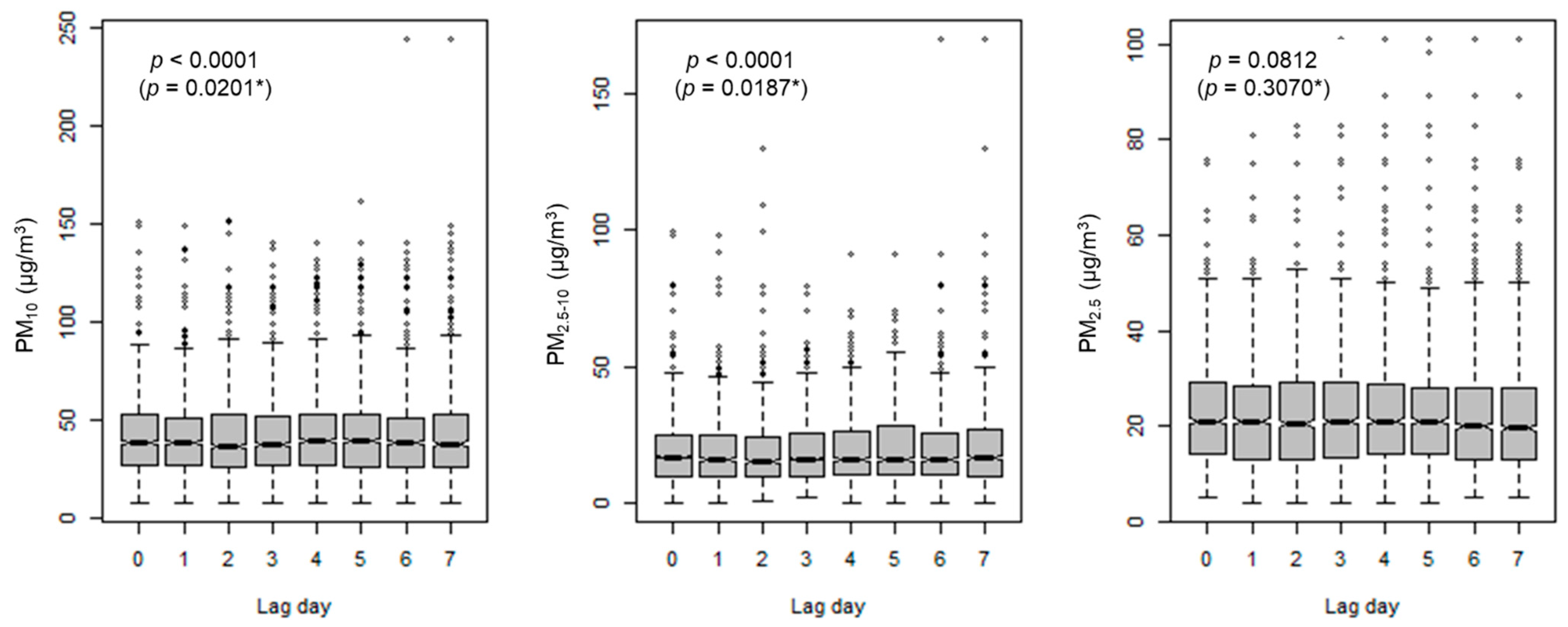
In the present study, all three PMs were strongly correlated with each other and PM2.5 had a higher daily variability than the other PMs. Furthermore, participants with PARP4 G-C-G and ERCC1 T-C haplotypes were susceptible to PMs exposure in relation to fasting glucose level.
The World Health Organization (WHO) has suggested a daily average level of 45 µg/m
3 for PM
10 and 15 µg/m
3 for PM
2.5 in their Global Air Quality Guidelines [
26]. Compared to the Global Air Quality Guidelines by WHO, our elderly population were exposed to a relatively and consistently high level of PMs, particularly PM
2.5. Considering correlations among three PMs on a specific lag day or among several lag days for the same PM in our study, PMs were temporally variable, particularly PM
2.5, although their levels were related to each other. In the present study, we chose lag day 4 for further analyses based on the strongest associations of PMs on lag day 4 with glucose level in the present study, and found the biggest change in glucose level after PM
2.5–10 exposure, followed by PM
2.5 and PM
10. The biggest change in glucose level after PM
2.5–10 exposure was supported by previously reported studies. Liang et al. [
27] showed that each increase of 10 µg/m
3 in 3-day moving averages of PM significantly increased the risk of outpatient visits of pneumonia, bronchiolitis, and asthma, regardless of PM size, with PM
2.5–10 showing the biggest effect size (4.36% of PM
10, 9.19% of PM
2.5–10, and 3.71% of PM
2.5 for outpatient visits of pneumonia; 3.12% of PM
10, 9.13% of PM
2.5–10, and 3.21% of PM
2.5 for bronchiolitis; and 3.33% of PM
10, 11.69% of PM
2.5–10, and 3.45% of PM
2.5 for asthma). Lei et al. [
28] showed that PM
2.5–10 had stronger associations with the loss of lung function than PM
2.5. The greatest effect size for PM
2.5–10 in our analyses could be attributable to the difference in the amount of lipopolysaccharide (LPS) among PMs. Biologic components intrinsic to PM such as LPS could directly activate Toll-like receptors, leading to inflammation [
29]. In fact, LPS was reported to induce metabolic syndromes, such as IR [
30,
31]. Moreover, LPS was known to be dominant in urbanized environments in Asia [
30]. The amount of LPS was reported to be higher in PM
10 than in PM
2.5 because of higher level of LPS in PM
2.5–10 [
30]. Because the effect size of PM
10 could be offset by that of PM
2.5 in our study, the effect size of PM
2.5–10 on glucose level was the greatest. However, there is still a debate about which PM could play a role in oxidative-stress-related DNA damage. Feng et al. [
32] indicated that DNA damage caused by fine particles, ranging from 0.43 to 2.1 μm in size, was greater than the damage caused by coarse particles, ranging from 4.7 to 10 μm in size. They also suggested that greater DNA damage in fine particles could be attributable to heavy metals enriched in fine particles [
32]. Therefore, in the future, we need to clarify which PM size is more important for the prevention of oxidative-stress-related DNA damage as well as their biological functions and mechanisms.
DNA repair proteins such as PARP4 and ERCC1 can protect against metabolic dysfunction including IR [
20,
33]. Both PARP and ERCC family members were reported to be involved in the pathway of IR regulation through repairing oxidative DNA damage and inhibiting autoinflammatory response [
20,
33]. However, there is a lack of knowledge regarding the relation between PARP4, a particular member of PARP family, and fasting glucose level. Furthermore, the effect of
PARP4 and
ERCC1 variations on the relation between PMs exposure and glucose level has not yet been reported, although positive associations between DNA damage accumulation and the development of metabolic disorders were reported [
33]. Because genetic polymorphisms of two repair genes,
PARP4 and
ERCC1, can affect DNA repair efficiency [
34], their polymorphisms may modify the relation between PMs exposure and glucose level. In the present study, we found that participants with
PARP4 G-C-G and
ERCC1 T-C haplotypes were apparently susceptible to an increase in glucose level in relation to exposure to all three PMs, although the effect size was slightly different by PMs. Although no study reported the effects of
PARP4 and
ERCC1 polymorphisms on relation between PMs exposure and fasting glucose level, several studies supported the potential idea that those polymorphisms could affect the impact of PMs exposure on glucose level. For the
ERCC1 gene, the defective
ERCC1 gene could increase the incidence of vascular diseases [
35], and genetic polymorphisms of
ERCC1 could affect the efficiency of chemotherapy, as well as the susceptibility of a variety of diseases, including lung cancer and several cardiovascular diseases [
36,
37,
38]. Particularly, the
ERCC1 rs11615 T allele was found to be a risk factor for developing non-small cell lung cancer [
37]. Because the lungs can be directly exposed to PMs [
8,
11,
12,
13], the effects of these SNPs are plausible. In fact, the risk effect of rs11615 T allele [
37] was consistent with our results, which we obtained with regard to both PMs and glucose level. Studies on
ERCC1 rs3212961 were controversial, although the rs3212961 C allele was associated with a shorter overall survival in gastric cancer patients [
39] and a higher risk for non-Hodgkin lymphoma development [
40], which were consistent with results of the present study. However, we found no evidence for the relation between the three SNPs of the
PARP4 gene examined in the present study and DNA damage or lung oxidation, although rs17080653, another intronic variant in
PARP4, showed a protective effect on head and neck cancer [
41] and inter-individual differences in DNA repair processes [
42].
In the present study, we found interactions between PMs exposure and PARP4 genotypes or diplotypes in relation to glucose level. All three PMs, in particular PM10 and PM2.5, showed interactions with PARP4 diplotypes, although the direct effect of PARP4 gene was not found. This meant the active functions of both environmental and genetic factors in relation with fasting glucose level, with an emphasis on the environmental factors in regulating genes. However, we did not find any significant interactions between PMs exposure and ERCC1 diplotypes, although ERCC1 rs11615 showed marginal interactions with PM10 and PM2.5 in relation with glucose. Therefore, in the future, we need to validate the interactive effect of the ERCC1 gene with PMs in relation to glucose level with a larger sample size.
To the best of our knowledge, the present study is the first to explore the effects of genetic modifications of
PARP4 and
ERCC1 on relations between three PM species and fasting blood glucose level, targeting elders who are susceptible to environmental pollutants. Although we used a panel study design, which could increase statistical power, our study also had limitations. First, we did not control for other air pollutants (O
3 or NO
2) and
GST family genes polymorphisms, although we found adverse effects of O
3 and NO
2 exposures on the glucose level and modification of
GSTM1 and
T1, as well as an impact of
P1 polymorphisms on the relation between PM
10 exposure and glucose level, in a previous study [
4]. Because too many missing data could be produced if we matched O
3 and NO
2 to PM
10 and PM
2.5 on a daily basis, which could lead to non-significance due to small numbers of data used in the analyses, we did not control O
3 or NO
2 in our statistical models. This was the same for the
GSTM1,
T1, and
P1 genes. In the future, we need to confirm whether the modification effect of
PARP4 and
ERCC1 polymorphisms on the relations between PMs exposure and glucose level still remains after controlling for these factors with a larger sample size. Second, in the present study, we only explored the acute effects of PMs on glucose level. Therefore, we need to clarify whether our genetic modification effect still remains after long-term exposure and short-term exposure. Third, in the future, we need to consider the chemical nature of the particles to confirm the causality of particles as well as genes in relation to glucose level.




{kind=link}
