Tomato Twisted Leaf Virus: A Novel Indigenous New World Monopartite Begomovirus Infecting Tomato in Venezuela

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Begomovirus Detection
2.2. Cloning of Full-Length Viral Genomes
2.3. Pairwise Identity, Phylogenetic, and Recombination Analyses
2.4. Construction of an Infectious Clone of Isolate Be6.6H for Agroinoculation
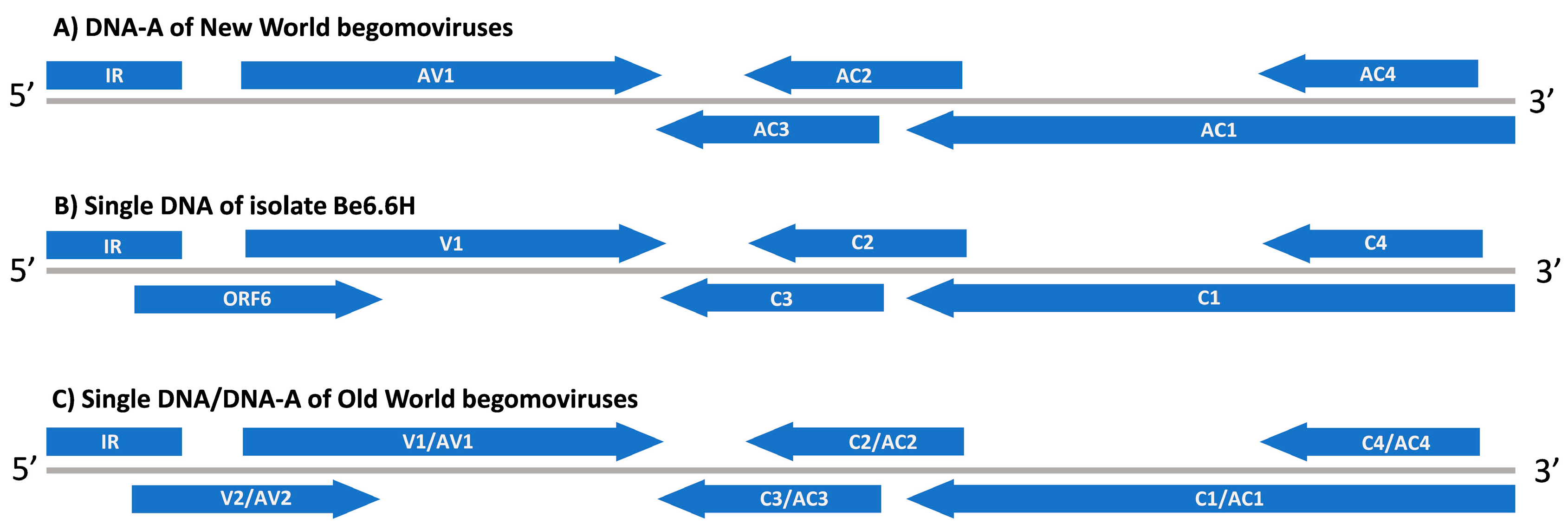
2.5. Genetic and Functional Analyses of a Sixth ORF in Be6.6H Isolate
3. Results
3.1. Sample Collection and Begomovirus Detection
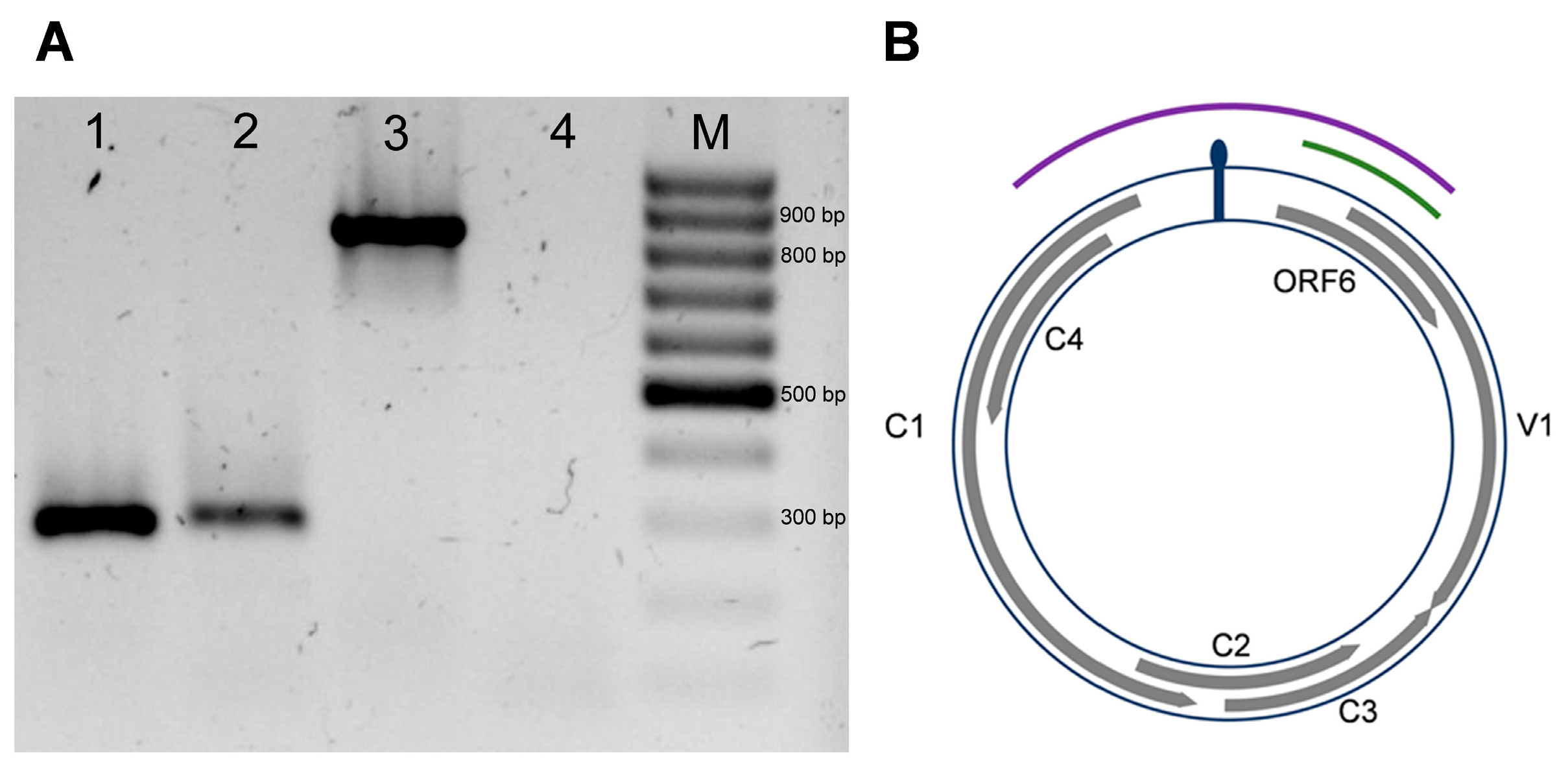
3.2. Cloning of Full-Length Viral Genome
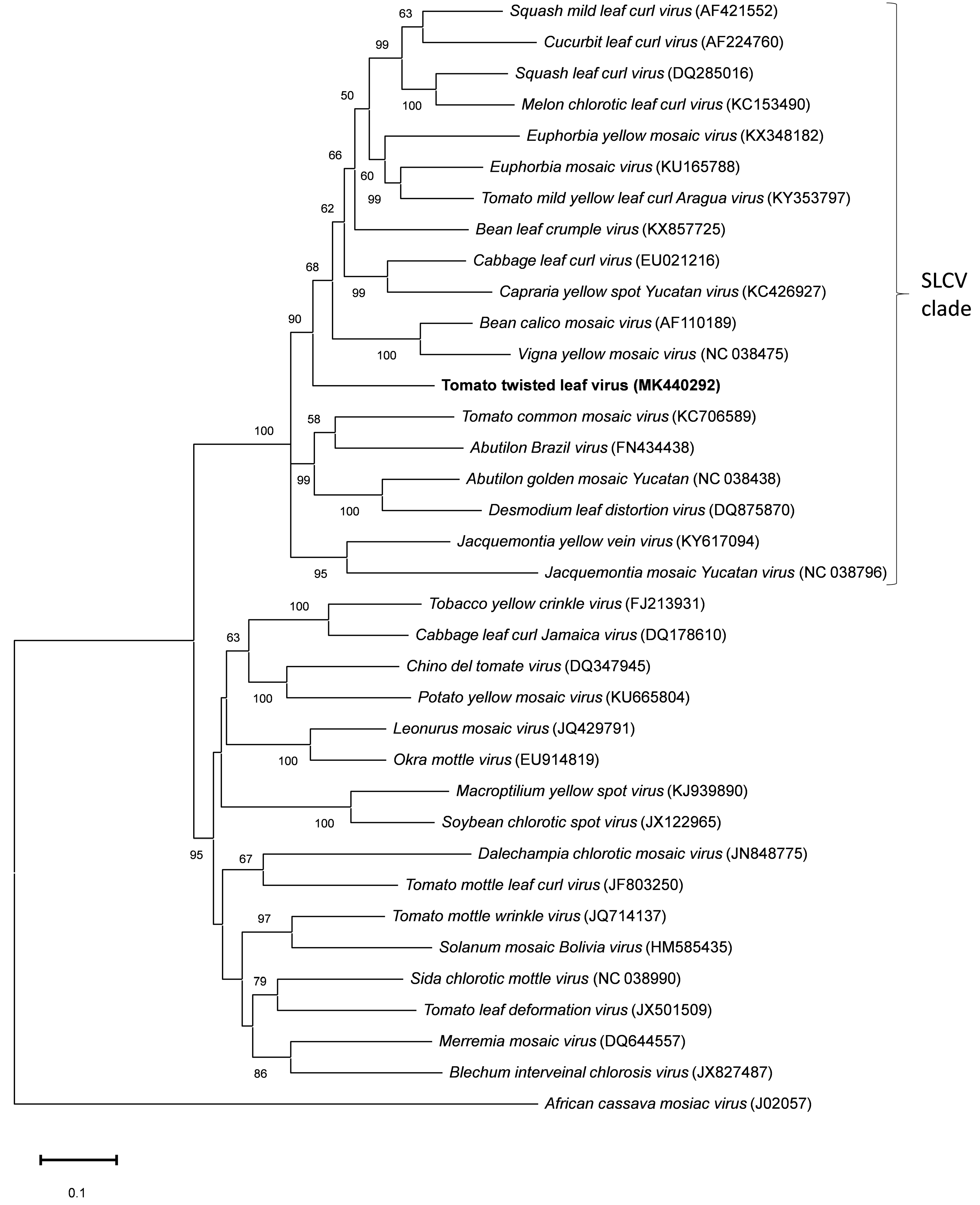
3.3. Pairwise, Phylogenetic, and Recombination Analyses
3.4. Infectivity of Clone Be6.6H in Tomato and N. Benthamiana Plants
3.5. Genetic Analysis of a Putative Novel Sixth ORF in Be6.6H Isolate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [Green Version]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the insect supervectors Bemisia tabaci and Frankliniella occidentalis in the emergence and global spread of plant viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.F.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy base on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef] [PubMed]
- Nawaz-Ul-Rehman, M.S.; Fauquet, C.M. Evolution of geminiviruses and their satellites. FEBS Lett. 2009, 583, 1825–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquez-Martin, B.; Aragon-Caballero, L.; Fiallo-Olive, E.; Navas-Castillo, J.; Moriones, E. Tomato leaf deformation virus, a novel begomovirus associated with a severe disease of tomato in Peru. Eur. Plant Pathol. 2011, 129, 1–7. [Google Scholar] [CrossRef]
- Melgarejo, T.A.; Kon, T.; Rojas, M.R.; Paz-Carrasco, L.; Zerbini, F.M.; Gilbertson, R.L. Characterization of a New World monopartite begomovirus causing leaf curl disease of tomato in Ecuador and Peru reveals a new direction in geminivirus evolution. J. Virol. 2013, 87, 5397–5413. [Google Scholar] [CrossRef]
- Sanchez-Campos, S.; Martinez-Ayala, A.; Marquez-Martin, B.; Aragon-Caballero, L.; Navas-Castillo, J.; Moriones, E. Fulfilling Koch’s postulates confirms the monopartite nature of tomato leaf deformation virus, a begomovirus native to the New World. Virus Res. 2013, 173, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Geraud-Pouey, F.; Chirinos, D.T.; Galindo-Castro, I.; Franco, M.A.; Santana, M.A.; Gillis, A.; Romay, G. Occurrence of six begomoviruses infecting tomato fields in Venezuela and genetic characterization of Potato yellow mosaic virus isolates. J. Phytopathology 2016, 164, 697–703. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Rojas, M.R.; Russell, D.R.; Maxwell, D.P. Use of the asymmetric polymerase chain reaction and DNA sequencing to determine genetic variability of bean golden mosaic geminivirus in the Dominican Republic. J. Gen. Virol. 1991, 72, 2843–2848. [Google Scholar] [CrossRef]
- Rojas, M.A.; Gilbertson, R.L.; Russell, D.R.; Maxwell, D.P. Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses. Plant Dis. 1993, 77, 340–347. [Google Scholar] [CrossRef]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: a user-friendly biologically sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Fourment, M.; Gibbs, A.J.; Gibbs, M.J. SWeBLAST: a sliding window web-based BLAST tool for recombinant analysis. J. Virol. Methods 2008, 152, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Murrell, B.; Golden, M.; Khoosa, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, K.; Geraud-Pouey, F.; Chirinos, D.; Romay, G.; Marys, E. Tomato chlorotic leaf distortion virus, a new bipartite begomovirus infecting Solanum lycopersicum and Capsicum chinense in Venezuela. Arch. Virol. 2011, 56, 2263–22636. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, F.R.; Albuquerque, L.C.; Oliveira, C.L.; Cruz, A.R.R.; Rocha, W.B.; Pereira, T.G.; Naito, F.Y.B.; Dias, N.d.M.; Nagata, T.; Faria, J.C.; et al. Molecular and biological characterization of a new Brazilian begomovirus, Euphorbia yellow mosaic virus (EuYMV), infecting Euphorbia heterophylla plants. Arch. Virol. 2011, 156, 2063–2069. [Google Scholar] [CrossRef] [PubMed]
- Polston, J.; Anderson, P. The emergence of whitefly-transmitted geminiviruses in tomato in the western hemisphere. Plant Dis. 1997, 81, 1358–1369. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Chirinos, D.T.; Geraud-Pouey, F.; Navas-Castillo, J. Complete genome sequence of jacquemontia yellow vein virus, a novel begomovirus infecting Jacquemontia tamnifolia in Venezuela. Arch. Virol. 2017, 162, 2463–2466. [Google Scholar] [CrossRef]
- Lefeuvre, P.; Lett, J.M.; Reynaud, B.; Martin, D.P. Avoidance of protein fold disruption in natural virus recombinants. PLoS Pathog. 2007, 3, 1782–1789. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Chirinos, D.T.; Geraud-Pouey, F.; Moriones, E.; Navas-Castillo, J. Complete genome sequence of Jacquemontia yellow mosaic virus, a novel begomovirus from Venezuela related to other New World bipartite begomoviruses infecting Convolvulaceae. Arch. Virol. 2014, 159, 1857–1860. [Google Scholar] [CrossRef] [PubMed]
- Romay, G.; Geraud-Pouey, F.; Chirinos, D.T.; Santana, M.A.; Galindo-Castro, I.; Márquez, L.M. Microsatellites reveal widespread predominance of an invasive over an indigenous Bemisia tabaci (Gennadius) in Venezuela. Phytoparasitica 2011, 39, 419–428. [Google Scholar] [CrossRef]
- Chirinos, D.T.; Geraud-Pouey, F.; Romay, G.; Fernandez, C.; Bastidas, L.; Flores, L.; Güerere, P. Infección por begomovirus en plantas de tomate propagadas bajo diferentes condiciones de protección física de semilleros. Bioagro 2014, 26, 57–62. [Google Scholar]
- Vu, S.; Melgarejo, T.A.; Chen, L.; Souza, J.O.; Macedo, M.A.; Inoue-Nagata, A.K.; Gilbertson, R.L. Evidence that tomato mottle leaf curl virus from Northeastern Brazil is an indigenous New World monopartite begomovirus. Phytopathology 2015, 105, S4.143. [Google Scholar]
- Macedo, M.A.; Albuquerque, L.C.; Maliano, M.R.; Souza, J.O.; Rojas, M.R.; Inoue-Nagata, A.K.; Gilbertson, R.L. Characterization of tomato leaf curl purple vein virus, a new monopartite New World begomovirus infecting tomato in Northeast Brazil. Arch. Virol. 2018, 163, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos-Hernández, B.; Mauricio-Castillo, J.A.; Cardenas-Conejo, Y.; Guevara-González, R.G.; Arguello-Astorga, G.R. A new strain of tomato severe leaf curl virus and a unique variant of tomato yellow leaf curl virus from Mexico. Arch. Virol. 2012, 157, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Kvarnheden, A.; Marcenaro, D.; Valkonen, J.P. Sequence characterization of tomato leaf curl Sinaloa virus and tomato severe leaf curl virus: phylogeny of New World begomoviruses and detection of recombination. Arch. Virol. 2005, 150, 1281–1299. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Seah, Y.; Marr, C.; Varsani, A.; Kraberger, S.; Stainton, D.; Moriones, E.; Polston, J.; Duffy, S.; Breitbart, M. Vector-enabled metagenomic (VEM) surveys using whiteflies (Aleyrodidae) reveal novel begomovirus species in the New and Old Worlds. Viruses 2015, 7, 5553–5570. [Google Scholar] [CrossRef]
- Rothenstein, D.; Krenz, B.; Selchow, O.; Jeske, H. Tissue and cell tropism of Indian cassava mosaic virus (ICMV) and its AV2 (precoat) gene product. Virology 2007, 359, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Glick, E.; Zrachya, A.; Levy, Y.; Mett, A.; Gidoni, D.; Belausov, E.; Citovsky, V.; Gafni, Y. Interaction with host SGS3 is required for suppression of RNA silencing by tomato yellow leaf curl virus V2 protein. Proc. Natl. Acad. Sci. USA 2008, 105, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Ferro, C.G.; Silva, J.P.; Xavier, C.A.D.; Godinho, M.T.; Lima, A.T.M.; Mar, T.B.; Lau, D.; Zerbini, F.M. The ever-increasing diversity of begomoviruses infecting non-cultivated hosts: New species from Sida spp. and Leonurus sibiricus, plus two new world alphasatellites. Ann. Appl. Biol. 2017, 170, 204–218. [Google Scholar] [CrossRef]
- Ho, E.S.; Kuchie, J.; Duffy, S. Bioinformatic analysis reveals genome size reduction and the emergence of tyrosine phosphorylation site in the movement protein of New World bipartite begomoviruses. PLoS ONE 2014, 9, e111957. [Google Scholar] [CrossRef] [PubMed]
- Cantú-Iris, M.; Pastor-Palacios, G.; Mauricio-Castillo, J.A.; Bañuelos-Hernández, B.; Avalos-Calleros, J.A.; Juárez-Reyes, A.; Rivera-Bustamante, R.; Argüello-Astorga, G.R. Analysis of a new begomovirus unveils a composite element conserved in the CP gene promoters of several Geminiviridae genera: Clues to comprehend the complex regulation of late genes. PLoS ONE 2019, 14, e0210485. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 Begomovirus Species | ToTLV | EuMV | CabLCV | ToMYLCV | BLCrV | JacYVV |
|---|---|---|---|---|---|---|
| ToTLV (Venezuela) | 100.0 | |||||
| EuMV (Cuba) | 82.4 | 100.0 | ||||
| CabLCV (Mexico) | 82.1 | 82.5 | 100.0 | |||
| ToMYLCV (Venezuela) | 81.7 | 88.1 | 82.9 | 100.0 | ||
| BLCrV (Colombia) | 81.2 | 82.6 | 83.3 | 84.1 | 100.0 | |
| JacYVV (Venezuela) | 81.1 | 81.0 | 79.3 | 79.8 | 78.9 | 100.0 |
| EuYMV (Brazil) | 80.7 | 82.3 | 80.7 | 84.6 | 80.5 | 77.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romay, G.; Geraud-Pouey, F.; Chirinos, D.T.; Mahillon, M.; Gillis, A.; Mahillon, J.; Bragard, C. Tomato Twisted Leaf Virus: A Novel Indigenous New World Monopartite Begomovirus Infecting Tomato in Venezuela. Viruses 2019, 11, 327. https://0-doi-org.brum.beds.ac.uk/10.3390/v11040327
Romay G, Geraud-Pouey F, Chirinos DT, Mahillon M, Gillis A, Mahillon J, Bragard C. Tomato Twisted Leaf Virus: A Novel Indigenous New World Monopartite Begomovirus Infecting Tomato in Venezuela. Viruses. 2019; 11(4):327. https://0-doi-org.brum.beds.ac.uk/10.3390/v11040327
Chicago/Turabian StyleRomay, Gustavo, Francis Geraud-Pouey, Dorys T. Chirinos, Mathieu Mahillon, Annika Gillis, Jacques Mahillon, and Claude Bragard. 2019. "Tomato Twisted Leaf Virus: A Novel Indigenous New World Monopartite Begomovirus Infecting Tomato in Venezuela" Viruses 11, no. 4: 327. https://0-doi-org.brum.beds.ac.uk/10.3390/v11040327