
Continuing Orthohantavirus Circulation in Deer Mice in Western Montana

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biosafety
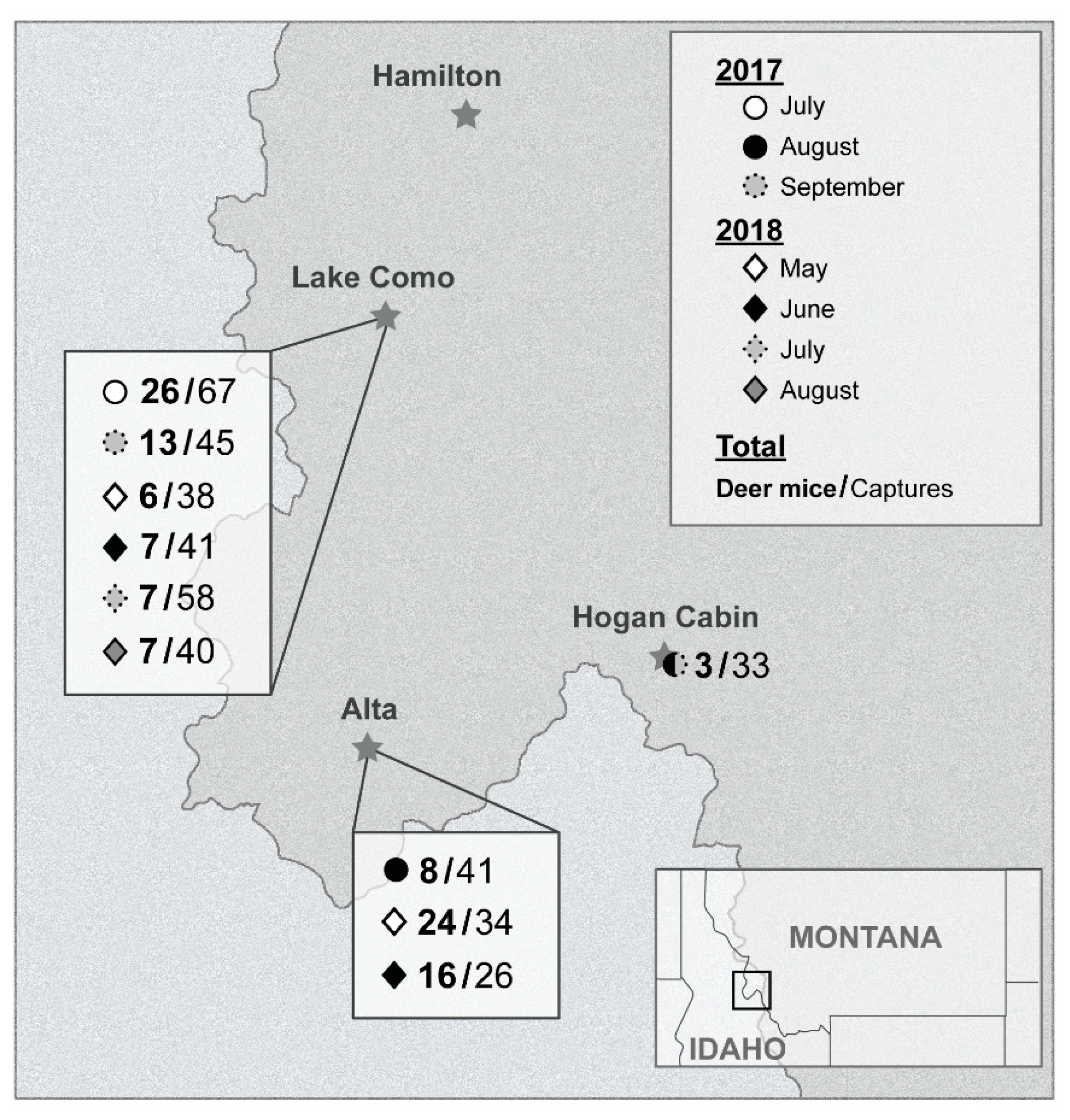
2.2. Field Locations
2.3. Animal Trapping
2.4. Sample Processing
2.5. qRT-PCR
2.6. Serology
2.7. Sequencing and Phylogenetic Analysis
2.8. Virus Isolation In Vitro and In Vivo
3. Results
3.1. Ecological Investigation
3.2. Molecular Investigation
3.3. Phylogenetic Analysis
3.4. Virus Isolation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laenen, L.; Vergote, V.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Maes, P. Hantaviridae: Currentclassification and future perspectives. Viruses 2019, 11, 788. [Google Scholar] [CrossRef] [Green Version]
- Avšič-Županc, T.; Saksida, A.; Korva, M. Hantavirus infections. Clin. Microbiol. Infect. 2019, 21, e6–e16. [Google Scholar] [CrossRef] [Green Version]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.E.; Rollin, P.; Ksiazek, T.G.; Feldmann, H.; Sanchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993, 262, 914–917. [Google Scholar] [CrossRef] [PubMed]
- Childs, J.E.; Ksiazek, T.G.; Spiropoulou, C.F.; Krebs, J.W.; Morzunov, S.; Maupin, G.O.; Gage, K.L.; Rollin, P.E.; Sarisky, J.; Enscore, R.E.; et al. Serologic and genetic identification of Peromyscus maniculatus as the primary rodent reservoir for a new hantavirus in the southwestern United States. J. Infect. Dis. 1994, 169, 1271–1280. [Google Scholar] [CrossRef] [Green Version]
- Frampton, J.W.; Lanser, S.; Nichols, C.R.; Ettestad, P.J. Sin nombre virus infection in 1959. Lancet 2006, 346, 781–782. [Google Scholar] [CrossRef]
- Monroe, M.C.; Morzunov, S.P.; Johnson, A.M.; Bowen, M.D.; Artsob, H.; Yates, T.; Peters, C.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Genetic Diversity and Distribution of Peromyscus-Borne Hantaviruses in North America. Emerg. Infect. Dis. 1999, 5, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Carver, S.; Mills, J.N.; Parmenter, C.A.; Parmenter, R.R.; Richardson, K.S.; Harris, R.L.; Douglass, R.J.; Kuenzi, A.J.; Luis, A.D. Toward a Mechanistic Understanding of Environmentally Forced Zoonotic Disease Emergence: Sin Nombre Hantavirus. Bioscience 2015, 65, 651–666. [Google Scholar] [CrossRef] [Green Version]
- Hantavirus Disease, by State of Reporting. Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/hantavirus/surveillance/reporting-state.html (accessed on 13 December 2019).
- Douglass, R.J.; Semmens, W.J.; Mills, J.N.; Zanto, S.N.; Bond, C.W.; Wilson, T.; Van Horn, R.C. Longitudinal studies of Sin Nombre virus in deer mouse-dominated ecosystems of Montana. Am. J. Trop. Med. Hyg. 2001, 65, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kuenzi, A.J.; Bond, C.W.; Douglass, R.J.; Mills, J.N.; White, D. Antibody to sin nombre virus in rodents associated with peridomestic habitats in west central Montana. Am. J. Trop. Med. Hyg. 2001, 64, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.N.; Johnson, J.M.; Ksiazek, T.G.; Ellis, B.A.; Rollin, P.E.; Yates, T.L.; Minn, M.O.; Johnson, M.R.; Campbell, M.L.; Miyashiro, J.; et al. A survey of hantavirus antibody in small-mammal populations in selected United States National Parks. Am. J. Trop. Med. Hyg. 1998, 58, 525–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglass, R.J.; Vadell, M.V. How much effort is required to accurately describe the complex ecology of a rodent-borne viral disease? Ecosphere 2016, 7, 01368. [Google Scholar] [CrossRef] [PubMed]
- Kuenzi, A.J.; Douglass, R.J.; Bond, C.W. Sin Nombre virus in deer mice captured inside homes, southwestern Montana. Emerg. Infect. Dis. 2000, 6, 386–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuenzi, A.J.; Douglass, R.J.; Bond, C.W.; Calisher, C.H.; Mills, J.N. Long-term dynamics of Sin Nombre viral RNA and antibody in deer mice in Montana. J. Wildl. Dis. 2005, 41, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, F.; Shupert, W.L.; Haddock, E.; Twardoski, B.; Feldmann, H. Gamma Irradiation as an Effective Method for Inac-tivation of Emerging Viral Pathogens. Am. J. Trop. Med. Hyg. 2019, 100, 1275–1277. [Google Scholar] [CrossRef]
- Schountz, T.; Calisher, C.H.; Richens, T.R.; Rich, A.A.; Doty, J.B.; Hughes, M.T.; Beaty, B.J. Rapid Field Immunoassay for Detecting Antibody to Sin Nombre Virus in Deer Mice. Emerg. Infect. Dis. 2007, 13, 1604–1607. [Google Scholar] [CrossRef]
- Li, D.; Schmaljohn, A.L.; Anderson, K.; Schmaljohn, C.S. Complete Nucleotide Sequences of the M and S Segments of Two Hantavirus Isolates from California: Evidence for Reassortment in Nature among Viruses Related to Hantavirus Pulmonary Syndrome. Virology 1995, 206, 973–983. [Google Scholar] [CrossRef]
- Cases of Hantavirus in Montana 1993–2018 by County. Available online: www.dphhs.mt.gov/publichealth/cdepi/diseases/hantavirus (accessed on 13 December 2019).
- Mills, J.N.; Amman, B.R.; Glass, G.E. Ecology of Hantaviruses and Their Hosts in North America. Vector-Borne Zoonotic Dis. 2010, 10, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Safronetz, D.; Prescott, J.; Haddock, E.; Scott, D.P.; Feldmann, H.; Ebihara, H. Hamster-Adapted Sin Nombre Virus Causes Disseminated Infection and Efficiently Replicates in Pulmonary Endothelial Cells without Signs of Disease. J. Virol. 2013, 87, 4778–4782. [Google Scholar] [CrossRef] [Green Version]
- Safronetz, D.; Prescott, J.; Feldmann, F.; Haddock, E.; Rosenke, R.; Okumura, A.; Brining, D.; Dahlstrom, E.; Porcella, S.F.; Ebihara, H.; et al. Pathophysiology of hantavirus pulmonary syndrome in rhesus macaques. Proc. Natl. Acad. Sci. USA 2014, 111, 7114–7119. [Google Scholar] [CrossRef] [Green Version]
- Gardner, S.L.; Von Essen, S.; Berger, J.; Hjelle, B. Low Seroprevalence Among Farmers from Nebraska and Vicinity Suggests Low Level of Human Exposure to Sin Nombre Virus. J. Agromed. 2005, 10, 59–61. [Google Scholar] [CrossRef]
- Gonzalez, L.M.; Lindsey, A.; Hjelle, B.; Dominguez, D.; Brown, J.; Goade, D.; Jonsson, C.B. Prevalence of antibodies to Sin Nombre virus in humans living in rural areas of southern New Mexico and western Texas. Virus Res. 2001, 74, 177–179. [Google Scholar] [CrossRef]
- Wilken, J.A.; Jackson, R.; Materna, B.L.; Windham, G.C.; Enge, B.; Messenger, S.; Xia, D.; Knust, B.; Buttke, D.; Roisman, R.; et al. Assessing prevention measures and sin nombre hantavirus seroprevalence among workers at Yosemite National Park. Am. J. Ind. Med. 2015, 58, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Location | Year | Blood RT-PCR | Lung RT-PCR | Serology | |||
|---|---|---|---|---|---|---|---|
| No. Positive/No. Tested | % Positive | No. Positive/No. Tested | % Positive | No. Positive/No. Tested | % Positive | ||
| Lake Como | 2017 | 0/39 | 0 | 1/20 | 5 | 1/37 | 2.7 |
| Alta | 2/8 | 25 | 3/8 | 37.5 | 2/8 | 25 | |
| Hogan Cabin | 1/3 | 33.3 | 1/3 | 33.3 | 0/3 | 0 | |
| 2017 Total | 3/50 | 6 | 5/31 | 16.1 | 3/48 | 6.3 | |
| Lake Como | 2018 | 1/22 | 4.5 | 1/3 | 33.3 | 3/15 | 20 |
| Alta | 0/40 | 0 | 8/40 | 20 | 7/39 | 17.9 | |
| 2018 Total | 1/62 | 1.6 | 9/43 | 20.9 | 10/54 | 18.5 | |
| Total | 4/112 | 3.6 | 14/74 | 18.9 | 13/102 | 12.7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williamson, B.N.; Meade-White, K.; Boardman, K.; Schulz, J.E.; Telford, C.T.; Figueroa Acosta, D.M.; Bushmaker, T.; Fischer, R.J.; Rosenke, K.; Feldmann, H. Continuing Orthohantavirus Circulation in Deer Mice in Western Montana. Viruses 2021, 13, 1006. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061006
Williamson BN, Meade-White K, Boardman K, Schulz JE, Telford CT, Figueroa Acosta DM, Bushmaker T, Fischer RJ, Rosenke K, Feldmann H. Continuing Orthohantavirus Circulation in Deer Mice in Western Montana. Viruses. 2021; 13(6):1006. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061006
Chicago/Turabian StyleWilliamson, Brandi N., Kimberly Meade-White, Kristin Boardman, Jonathan E. Schulz, Carson T. Telford, Dania M. Figueroa Acosta, Trenton Bushmaker, Robert J. Fischer, Kyle Rosenke, and Heinz Feldmann. 2021. "Continuing Orthohantavirus Circulation in Deer Mice in Western Montana" Viruses 13, no. 6: 1006. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061006