
Peptide Derivatives of Platelet-Derived Growth Factor Receptor Alpha Inhibit Cell-Associated Spread of Human Cytomegalovirus

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Indirect Immunofluorescence
2.3. Effects of Entry Inhibitors on HCMV and HSV Infection
2.4. Mode of Action Analysis on the Single Particle Level
2.5. Synchronizing Cell-Associated Spread Using the Inhibitory Peptide GD30
2.6. Statistical Analysis
3. Results
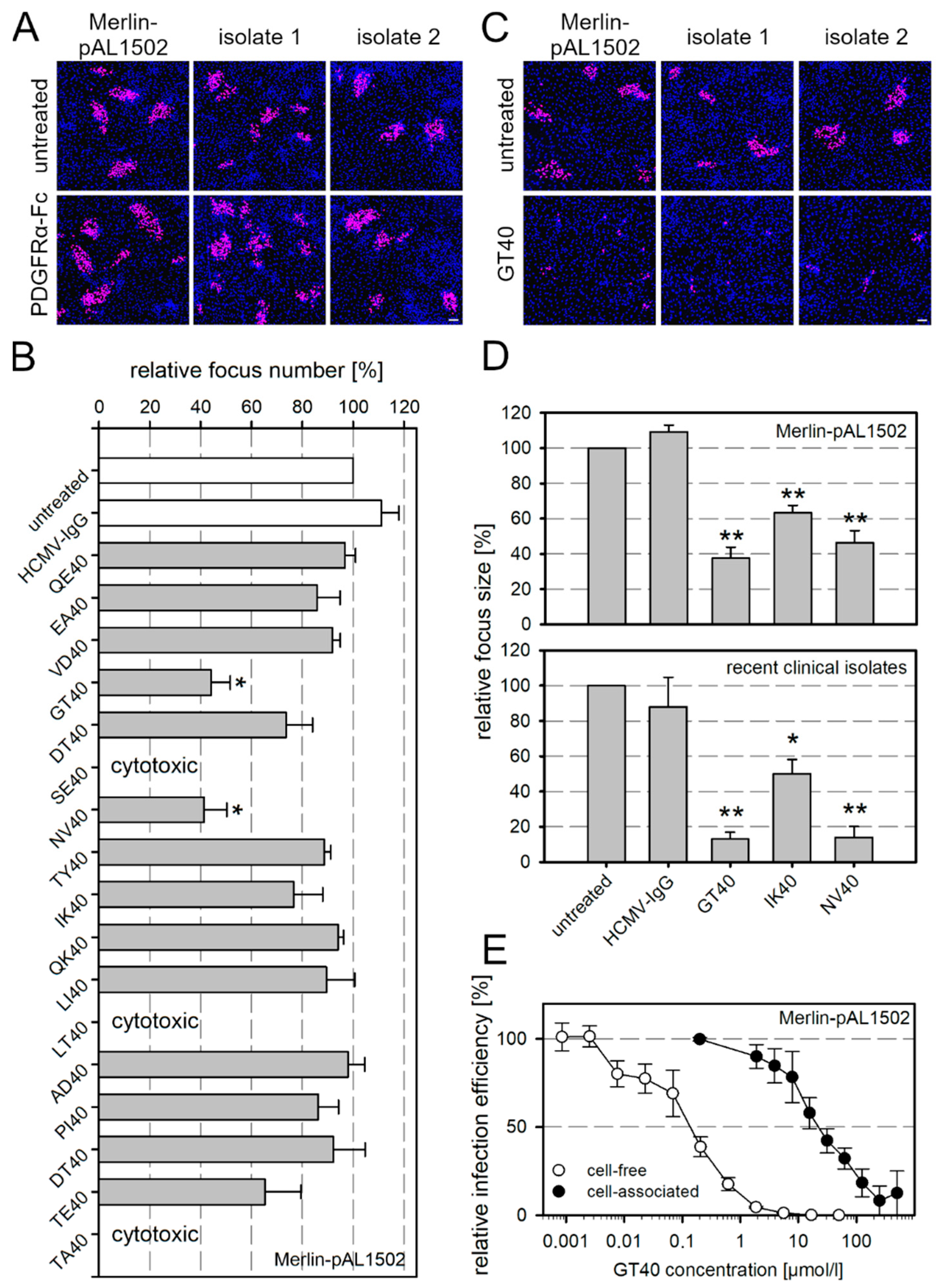
3.1. Cell-Associated Spread of HCMV Is Resistant to a Soluble Form of PDGFRα but Inhibited by Peptide Derivatives Thereof
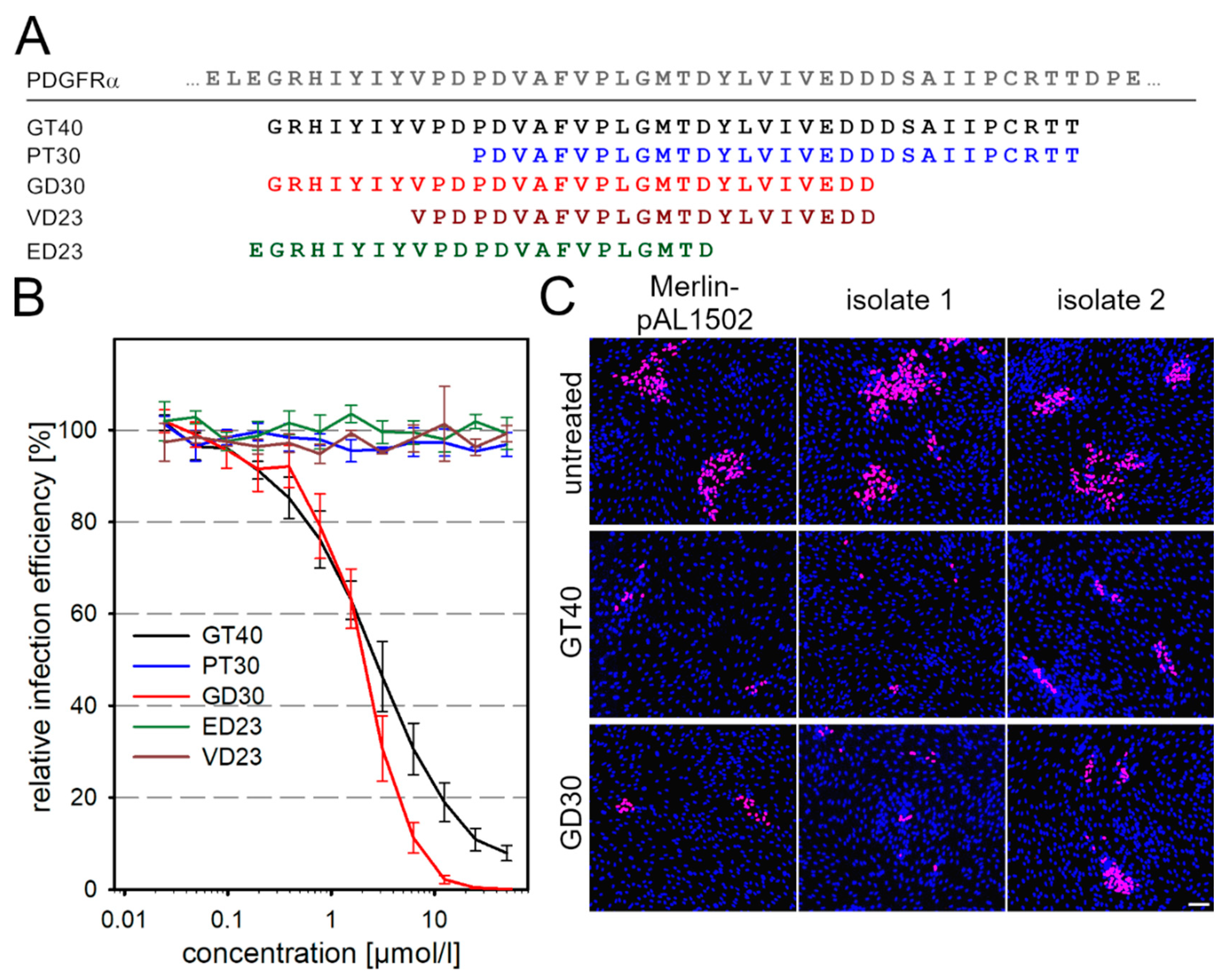
3.2. Targeted Modifications of the Peptide GT40 Enhance Its Efficiency against Cell-Free Virus but Not against Cell-Associated Spread
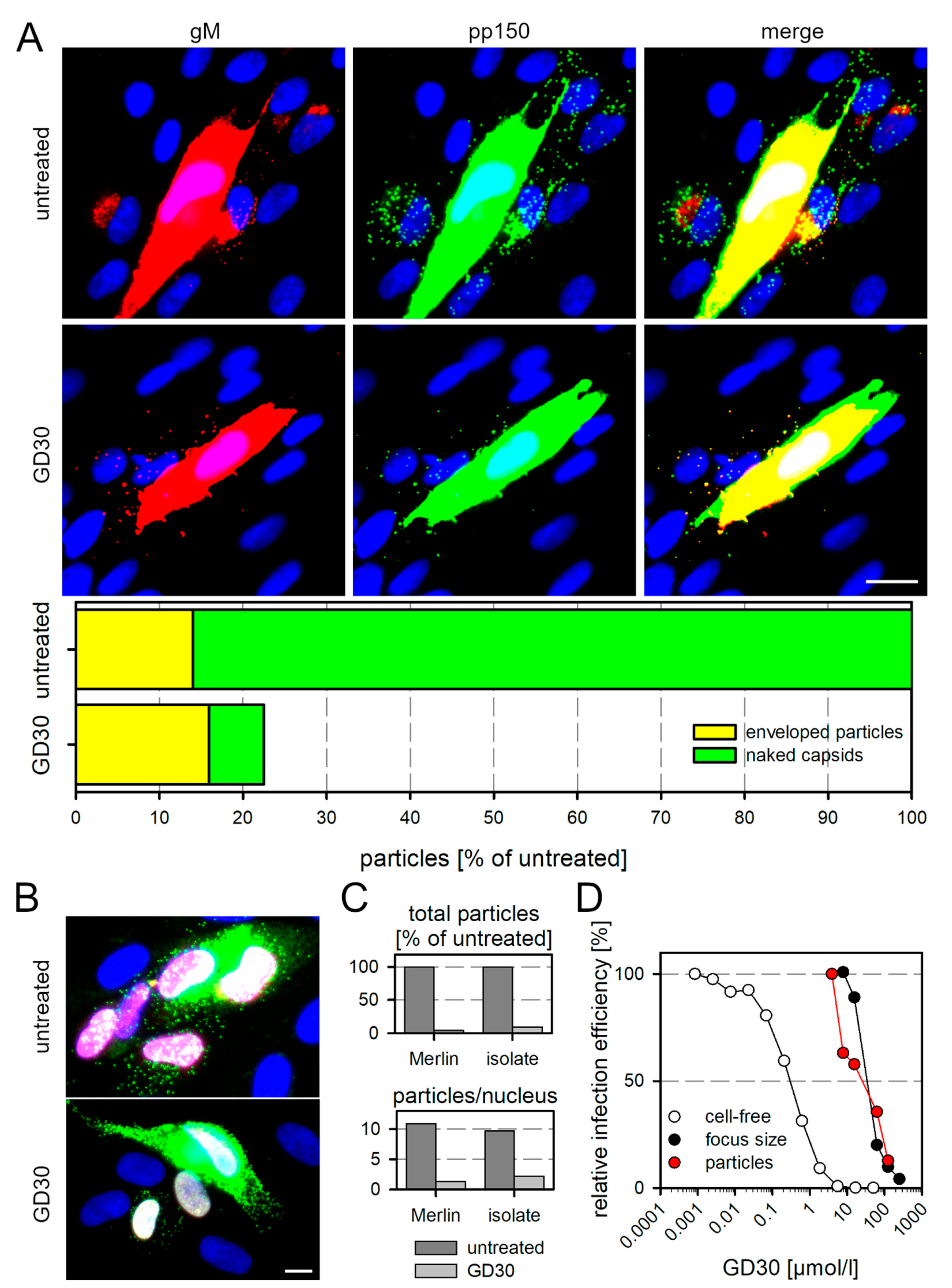
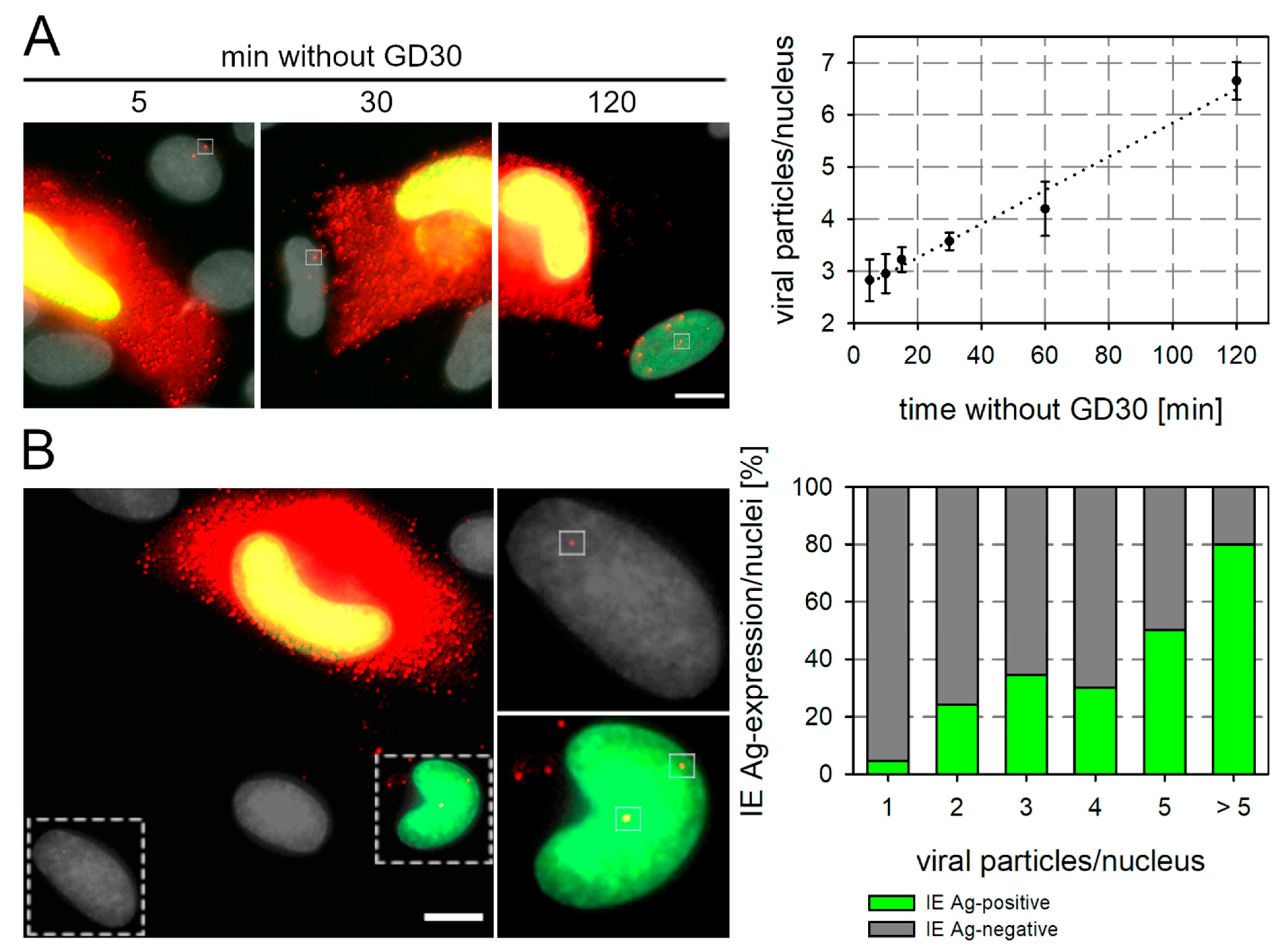
3.3. GD30 Reduces the Number of Transferred Viral Particles and Their Penetration Efficiency
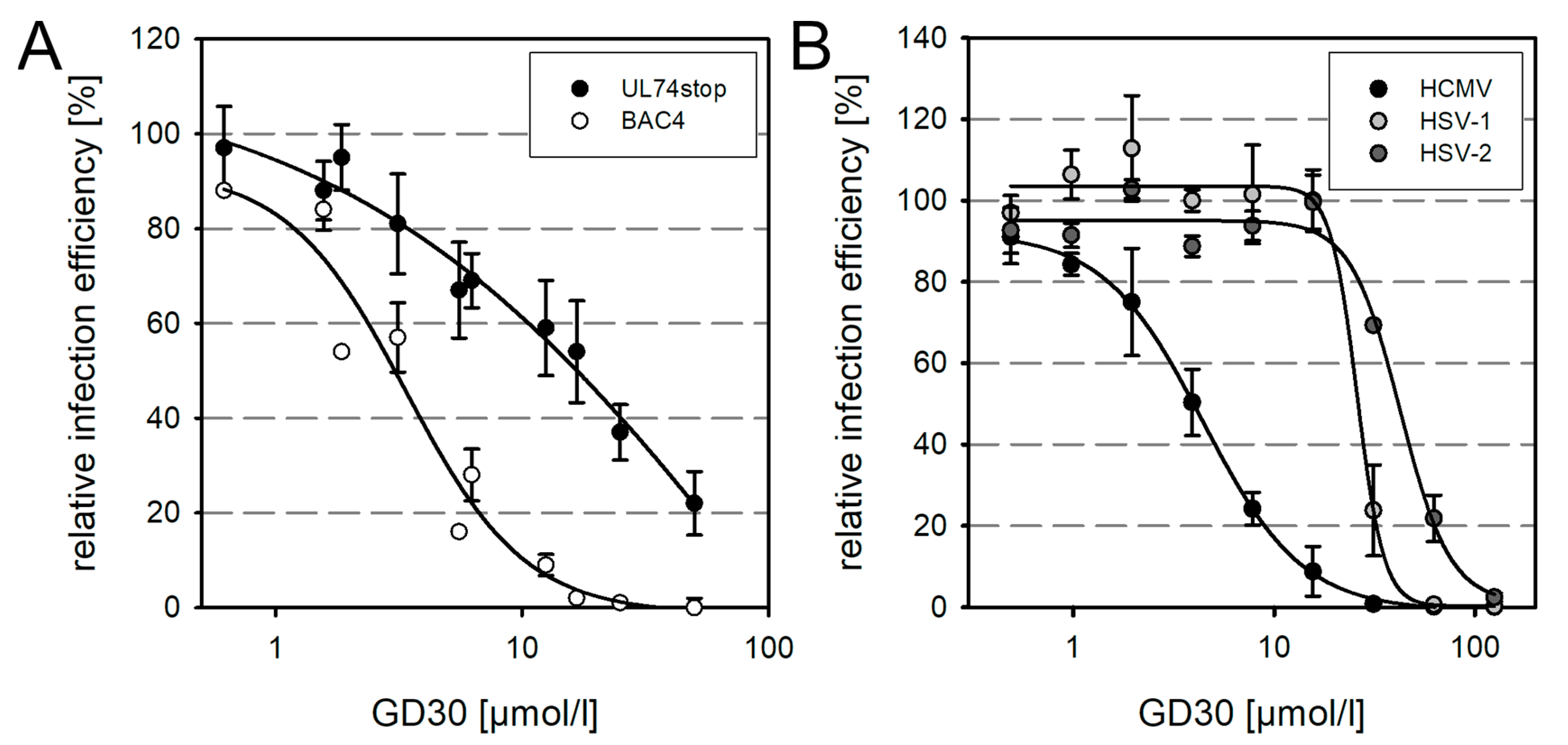
3.4. Evidence That GD30 Specifically Targets gO of HCMV but Also a Factor Shared with Other Herpesviruses
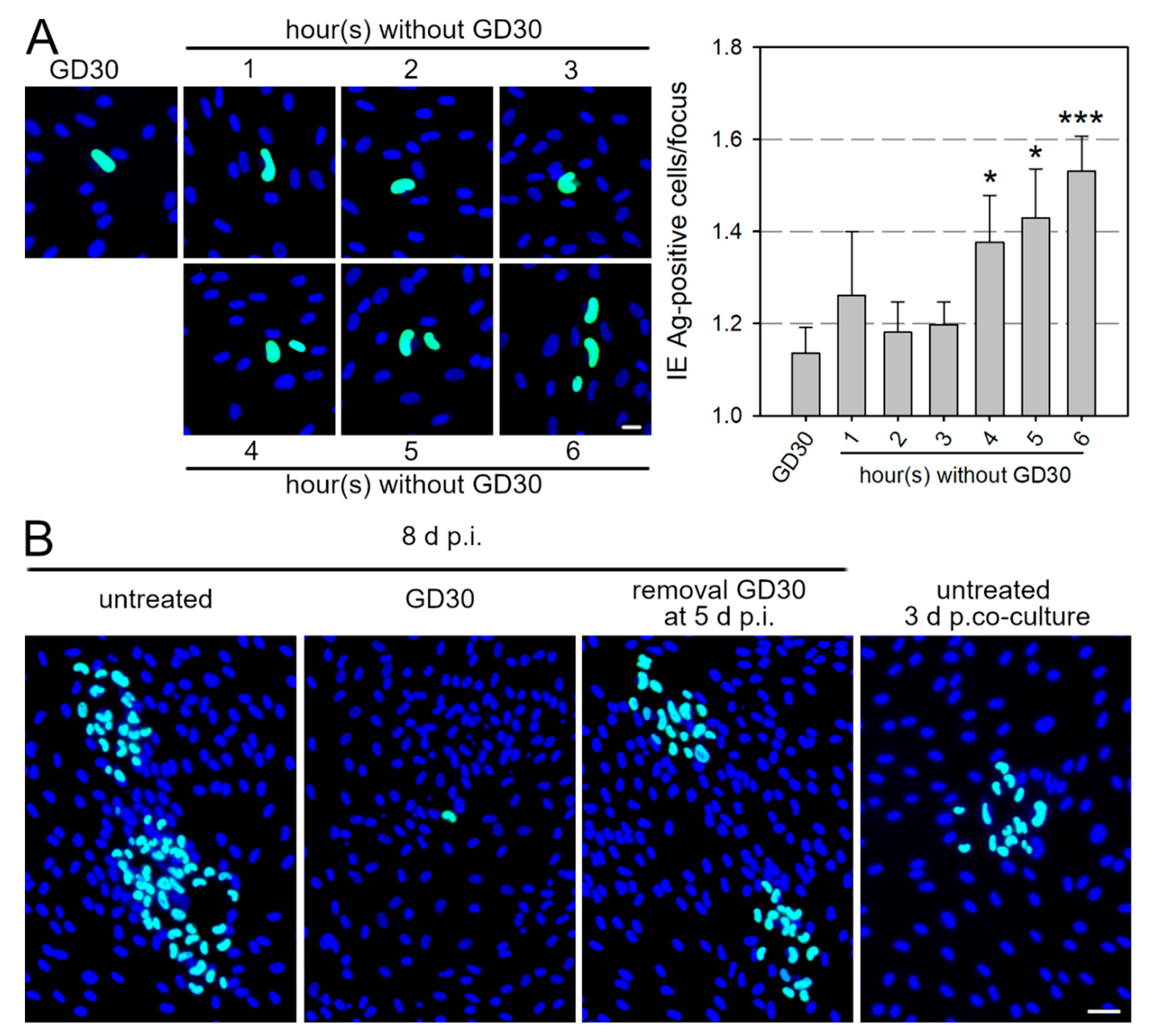
3.5. GD30 Inhibits Cell-Associated Spread of HCMV in a Reversible Manner
3.6. GD30 Can Be Used to Study Underlying Mechanisms of Cell-Associated Spread
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pass, R.F.; Zhang, C.; Evans, A.; Simpson, T.; Andrews, W.; Huang, M.-L.; Corey, L.; Hill, J.; Davis, E.; Flanigan, C.; et al. Vaccine Prevention of Maternal Cytomegalovirus Infection. N. Engl. J. Med. 2009, 360, 1191–1199. [Google Scholar] [CrossRef]
- Kagan, K.O.; Hamprecht, K. Cytomegalovirus Infection in Pregnancy. Arch. Gynecol. Obstet. 2017, 296, 15–26. [Google Scholar] [CrossRef]
- Kotton, C.N.; Kumar, D.; Caliendo, A.M.; Huprikar, S.; Chou, S.; Danziger-Isakov, L.; Humar, A. The Transplantation Society International CMV Consensus Group The Third International Consensus Guidelines on the Management of Cytomegalovirus in Solid-Organ Transplantation. Transplantation 2018, 102, 900–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljungman, P.; de la Camara, R.; Robin, C.; Crocchiolo, R.; Einsele, H.; Hill, J.A.; Hubacek, P.; Navarro, D.; Cordonnier, C.; Ward, K.N.; et al. Guidelines for the Management of Cytomegalovirus Infection in Patients with Haematological Malignancies and after Stem Cell Transplantation from the 2017 European Conference on Infections in Leukaemia (ECIL 7). Lancet Infect. Dis. 2019, 19, e260–e272. [Google Scholar] [CrossRef]
- El Helou, G.; Razonable, R.R. Safety Considerations with Current and Emerging Antiviral Therapies for Cytomegalovirus Infection in Transplantation. Expert Opin. Drug Saf. 2019, 18, 1017–1030. [Google Scholar] [CrossRef]
- Marty, F.M.; Ljungman, P.; Chemaly, R.F.; Maertens, J.; Dadwal, S.S.; Duarte, R.F.; Haider, S.; Ullmann, A.J.; Katayama, Y.; Brown, J.; et al. Letermovir Prophylaxis for Cytomegalovirus in Hematopoietic-Cell Transplantation. N. Engl. J. Med. 2017, 377, 2433–2444. [Google Scholar] [CrossRef] [PubMed]
- Douglas, C.M.; Barnard, R.; Holder, D.; Leavitt, R.; Levitan, D.; Maguire, M.; Nickle, D.; Teal, V.; Wan, H.; van Alewijk, D.C.J.G.; et al. Letermovir Resistance Analysis in a Clinical Trial of Cytomegalovirus Prophylaxis for Hematopoietic Stem Cell Transplant Recipients. J. Infect. Dis. 2020, 221, 1117–1126. [Google Scholar] [CrossRef]
- Gerna, G.; Lilleri, D.; Baldanti, F. An Overview of Letermovir: A Cytomegalovirus Prophylactic Option. Expert Opin. Pharmacother. 2019, 20, 1429–1438. [Google Scholar] [CrossRef]
- Jung, S.; Michel, M.; Stamminger, T.; Michel, D. Fast Breakthrough of Resistant Cytomegalovirus during Secondary Letermovir Prophylaxis in a Hematopoietic Stem Cell Transplant Recipient. BMC Infect. Dis. 2019, 19, 388. [Google Scholar] [CrossRef]
- Lischka, P.; Michel, D.; Zimmermann, H. Characterization of Cytomegalovirus Breakthrough Events in a Phase 2 Prophylaxis Trial of Letermovir (AIC246, MK 8228). J. Infect. Dis. 2016, 213, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Gulick, R.M. Investigational Antiretroviral Drugs: What Is Coming Down the Pipeline. Top. Antivir. Med. 2018, 25, 127–132. [Google Scholar] [PubMed]
- Haqqani, A.A.; Tilton, J.C. Entry Inhibitors and Their Use in the Treatment of HIV-1 Infection. Antivir. Res. 2013, 98, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Bogomolov, P.; Alexandrov, A.; Voronkova, N.; Macievich, M.; Kokina, K.; Petrachenkova, M.; Lehr, T.; Lempp, F.A.; Wedemeyer, H.; Haag, M.; et al. Treatment of Chronic Hepatitis D with the Entry Inhibitor Myrcludex B: First Results of a Phase Ib/IIa Study. J. Hepatol. 2016, 65, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Han, B.; Zhang, W.; Wu, W. Clinical Effects of NTCP-Inhibitor Myrcludex B. J. Viral Hepat. 2021, 28, 852–858. [Google Scholar] [CrossRef]
- Loglio, A.; Ferenci, P.; Uceda Renteria, S.C.; Tham, C.Y.L.; van Bömmel, F.; Borghi, M.; Holzmann, H.; Perbellini, R.; Trombetta, E.; Giovanelli, S.; et al. Excellent Safety and Effectiveness of High-Dose Myrcludex-B Monotherapy Administered for 48 weeks in HDV-Related Compensated Cirrhosis: A Case Report of 3 Patients. J. Hepatol. 2019, 71, 834–839. [Google Scholar] [CrossRef]
- Wang, W.; Lempp, F.A.; Schlund, F.; Walter, L.; Decker, C.C.; Zhang, Z.; Ni, Y.; Urban, S. Assembly and Infection Efficacy of Hepatitis B Virus Surface Protein Exchanges in 8 Hepatitis D Virus Genotype Isolates. J. Hepatol. 2021, 75, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Kabanova, A.; Marcandalli, J.; Zhou, T.; Bianchi, S.; Baxa, U.; Tsybovsky, Y.; Lilleri, D.; Silacci-Fregni, C.; Foglierini, M.; Fernandez-Rodriguez, B.M.; et al. Platelet-Derived Growth Factor-α Receptor Is the Cellular Receptor for Human Cytomegalovirus GHgLgO Trimer. Nat. Microbiol. 2016, 1, 16082. [Google Scholar] [CrossRef]
- Stegmann, C.; Hochdorfer, D.; Lieber, D.; Subramanian, N.; Stöhr, D.; Laib Sampaio, K.; Sinzger, C. A Derivative of Platelet-Derived Growth Factor Receptor Alpha Binds to the Trimer of Human Cytomegalovirus and Inhibits Entry into Fibroblasts and Endothelial Cells. PLoS Pathog. 2017, 13, e1006273. [Google Scholar] [CrossRef]
- Wu, Y.; Prager, A.; Boos, S.; Resch, M.; Brizic, I.; Mach, M.; Wildner, S.; Scrivano, L.; Adler, B. Human Cytomegalovirus Glycoprotein Complex GH/GL/GO Uses PDGFR-α as a Key for Entry. PLoS Pathog. 2017, 13, e1006281. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.T.; Compton, T. The Human Cytomegalovirus UL74 Gene Encodes the Third Component of the Glycoprotein H-Glycoprotein L-Containing Envelope Complex. J. Virol. 1998, 72, 8191–8197. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Nelson, J.A.; Britt, W.J. Glycoprotein H-Related Complexes of Human Cytomegalovirus: Identification of a Third Protein in the GCIII Complex. J. Virol. 1997, 71, 3090–3097. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Lanchy, J.-M.; Ryckman, B.J. Human Cytomegalovirus GH/GL/GO Promotes the Fusion Step of Entry into All Cell Types, Whereas GH/GL/UL128-131 Broadens Virus Tropism through a Distinct Mechanism. J. Virol. 2015, 89, 8999–9009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dargan, D.J.; Douglas, E.; Cunningham, C.; Jamieson, F.; Stanton, R.J.; Baluchova, K.; McSharry, B.P.; Tomasec, P.; Emery, V.C.; Percivalle, E.; et al. Sequential Mutations Associated with Adaptation of Human Cytomegalovirus to Growth in Cell Culture. J. Gen. Virol. 2010, 91, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Frascaroli, G.; Sinzger, C. Distinct Properties of Human Cytomegalovirus Strains and the Appropriate Choice of Strains for Particular Studies. Methods Mol. Biol. 2021, 2244, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Schmidt, K.; Knapp, J.; Kahl, M.; Beck, R.; Waldman, J.; Hebart, H.; Einsele, H.; Jahn, G. Modification of Human Cytomegalovirus Tropism through Propagation in Vitro Is Associated with Changes in the Viral Genome. J. Gen. Virol. 1999, 80 (Pt 11), 2867–2877. [Google Scholar] [CrossRef]
- Yamane, Y.; Furukawa, T.; Plotkin, S.A. Supernatant Virus Release as a Differentiating Marker between Low Passage and Vaccine Strains of Human Cytomegalovirus. Vaccine 1983, 1, 23–25. [Google Scholar] [CrossRef]
- Griffiths, P.; Reeves, M. Pathogenesis of Human Cytomegalovirus in the Immunocompromised Host. Nat. Rev. Microbiol. 2021, 24, 1–15. [Google Scholar] [CrossRef]
- Jackson, J.W.; Sparer, T. There Is Always Another Way! Cytomegalovirus’ Multifaceted Dissemination Schemes. Viruses 2018, 10, 383. [Google Scholar] [CrossRef] [Green Version]
- Lemmermann, N.A.W.; Krmpotic, A.; Podlech, J.; Brizic, I.; Prager, A.; Adler, H.; Karbach, A.; Wu, Y.; Jonjic, S.; Reddehase, M.J.; et al. Non-Redundant and Redundant Roles of Cytomegalovirus GH/GL Complexes in Host Organ Entry and Intra-Tissue Spread. PLoS Pathog. 2015, 11, e1004640. [Google Scholar] [CrossRef]
- Zhong, P.; Agosto, L.M.; Munro, J.B.; Mothes, W. Cell-to-Cell Transmission of Viruses. Curr. Opin. Virol. 2013, 3, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Digel, M.; Sampaio, K.L.; Jahn, G.; Sinzger, C. Evidence for Direct Transfer of Cytoplasmic Material from Infected to Uninfected Cells during Cell-Associated Spread of Human Cytomegalovirus. J. Clin. Virol. 2006, 37, 10–20. [Google Scholar] [CrossRef]
- Gerna, G.; Percivalle, E.; Baldanti, F.; Sozzani, S.; Lanzarini, P.; Genini, E.; Lilleri, D.; Revello, M.G. Human Cytomegalovirus Replicates Abortively in Polymorphonuclear Leukocytes after Transfer from Infected Endothelial Cells via Transient Microfusion Events. J. Virol. 2000, 74, 5629–5638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.C.; Schröer, J.; Shenk, T. Human Cytomegalovirus Cell-to-Cell Spread in the Absence of an Essential Assembly Protein. Proc. Natl. Acad. Sci. USA 2005, 102, 2081–2086. [Google Scholar] [CrossRef] [Green Version]
- Falk, J.J.; Winkelmann, M.; Laib Sampaio, K.; Paal, C.; Schrezenmeier, H.; Alt, M.; Stanton, R.; Krawczyk, A.; Lotfi, R.; Sinzger, C. Large-Scale Screening of HCMV-Seropositive Blood Donors Indicates That HCMV Effectively Escapes from Antibodies by Cell-Associated Spread. Viruses 2018, 10, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiler, N.; Paal, C.; Adams, K.; Calcaterra, C.; Fischer, D.; Stanton, R.J.; Stöhr, D.; Laib Sampaio, K.; Sinzger, C. Role of Envelope Glycoprotein Complexes in Cell-Associated Spread of Human Cytomegalovirus. Viruses 2021, 13, 614. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Mangin, M.; Weinstock, C.; Topp, M.S.; Hebart, H.; Einsele, H.; Jahn, G. Effect of Serum and CTL on Focal Growth of Human Cytomegalovirus. J. Clin. Virol. 2007, 38, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Murrell, I.; Bedford, C.; Ladell, K.; Miners, K.L.; Price, D.A.; Tomasec, P.; Wilkinson, G.W.G.; Stanton, R.J. The Pentameric Complex Drives Immunologically Covert Cell-Cell Transmission of Wild-Type Human Cytomegalovirus. Proc. Natl. Acad. Sci. USA 2017, 114, 6104–6109. [Google Scholar] [CrossRef] [Green Version]
- Gerna, G.; Percivalle, E.; Sarasini, A.; Revello, M.G. Human Cytomegalovirus and Human Umbilical Vein Endothelial Cells: Restriction of Primary Isolation to Blood Samples and Susceptibilities of Clinical Isolates from Other Sources to Adaptation. J. Clin. Microbiol. 2002, 40, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prix, L.; Maierl, J.; Jahn, G.; Hamprecht, K. A Simplified Assay for Screening of Drug Resistance of Cell-Associated Cytomegalovirus Strains. J. Clin. Virol. 1998, 11, 29–37. [Google Scholar] [CrossRef]
- Waldman, W.J.; Sneddon, J.M.; Stephens, R.E.; Roberts, W.H. Enhanced Endothelial Cytopathogenicity Induced by a Cytomegalovirus Strain Propagated in Endothelial Cells. J. Med. Virol. 1989, 28, 223–230. [Google Scholar] [CrossRef]
- Jones, T.R.; Lee, S.-W.; Johann, S.V.; Razinkov, V.; Visalli, R.J.; Feld, B.; Bloom, J.D.; O’Connell, J. Specific Inhibition of Human Cytomegalovirus Glycoprotein B-Mediated Fusion by a Novel Thiourea Small Molecule. J. Virol. 2004, 78, 1289–1300. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Heim, K.P.; Che, Y.; Chi, X.; Qiu, X.; Han, S.; Dormitzer, P.R.; Yang, X. Prefusion Structure of Human Cytomegalovirus Glycoprotein B and Structural Basis for Membrane Fusion. Sci. Adv. 2021, 7, eabf3178. [Google Scholar] [CrossRef]
- Stanton, R.J.; Baluchova, K.; Dargan, D.J.; Cunningham, C.; Sheehy, O.; Seirafian, S.; McSharry, B.P.; Neale, M.L.; Davies, J.A.; Tomasec, P.; et al. Reconstruction of the Complete Human Cytomegalovirus Genome in a BAC Reveals RL13 to Be a Potent Inhibitor of Replication. J. Clin. Investig. 2010, 120, 3191–3208. [Google Scholar] [CrossRef] [Green Version]
- Sinzger, C.; Hahn, G.; Digel, M.; Katona, R.; Sampaio, K.L.; Messerle, M.; Hengel, H.; Koszinowski, U.; Brune, W.; Adler, B. Cloning and Sequencing of a Highly Productive, Endotheliotropic Virus Strain Derived from Human Cytomegalovirus TB40/E. J. Gen. Virol. 2008, 89, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Falk, J.J.; Laib Sampaio, K.; Stegmann, C.; Lieber, D.; Kropff, B.; Mach, M.; Sinzger, C. Generation of a Gaussia Luciferase-Expressing Endotheliotropic Cytomegalovirus for Screening Approaches and Mutant Analyses. J. Virol. Methods 2016, 235, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Stegmann, C.; Abdellatif, M.E.A.; Laib Sampaio, K.; Walther, P.; Sinzger, C. Importance of Highly Conserved Peptide Sites of Human Cytomegalovirus GO for Formation of the GH/GL/GO Complex. J. Virol. 2016, 91, e01339-16. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, V.; Greis, K.D.; Sztul, E.; Britt, W.J. Accumulation of Virion Tegument and Envelope Proteins in a Stable Cytoplasmic Compartment during Human Cytomegalovirus Replication: Characterization of a Potential Site of Virus Assembly. J. Virol. 2000, 74, 975–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laib Sampaio, K.; Lutz, C.; Engels, R.; Stöhr, D.; Sinzger, C. Selection of Human Cytomegalovirus Mutants with Resistance against PDGFRα-Derived Entry Inhibitors. Viruses 2021, 13, 1094. [Google Scholar] [CrossRef]
- Braun, B.; Sinzger, C. Transmission of Cell-Associated Human Cytomegalovirus Isolates between Various Cell Types Using Polymorphonuclear Leukocytes as a Vehicle. Med. Microbiol. Immunol. 2021, 210, 197–209. [Google Scholar] [CrossRef]
- Kschonsak, M.; Rougé, L.; Arthur, C.P.; Hoangdung, H.; Patel, N.; Kim, I.; Johnson, M.C.; Kraft, E.; Rohou, A.L.; Gill, A.; et al. Structures of HCMV Trimer Reveal the Basis for Receptor Recognition and Cell Entry. Cell 2021, 184, 1232–1244.e16. [Google Scholar] [CrossRef]
- Sattentau, Q.J. The Direct Passage of Animal Viruses between Cells. Curr. Opin. Virol. 2011, 1, 396–402. [Google Scholar] [CrossRef]
- Singh, B.K.; Pfaller, C.K.; Cattaneo, R.; Sinn, P.L. Measles Virus Ribonucleoprotein Complexes Rapidly Spread across Well-Differentiated Primary Human Airway Epithelial Cells along F-Actin Rings. mBio 2019, 10, e02434-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mothes, W.; Sherer, N.M.; Jin, J.; Zhong, P. Virus Cell-to-Cell Transmission. J. Virol. 2010, 84, 8360–8368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, C.M.; Shenk, T. Human Cytomegalovirus PUS27 G Protein-Coupled Receptor Homologue Is Required for Efficient Spread by the Extracellular Route but Not for Direct Cell-to-Cell Spread. J. Virol. 2011, 85, 3700–3707. [Google Scholar] [CrossRef] [Green Version]
- Schultz, E.P.; Lanchy, J.-M.; Day, L.Z.; Yu, Q.; Peterson, C.; Preece, J.; Ryckman, B.J. Specialization for Cell-Free or Cell-to-Cell Spread of BAC-Cloned Human Cytomegalovirus Strains Is Determined by Factors beyond the UL128-131 and RL13 Loci. J. Virol. 2020, 94, e00034-20. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.L.; Lamorte, L.; Sepulveda, E.; Lorenz, I.C.; Gauthier, A.; Franti, M. Neutralizing Antibodies Are Unable to Inhibit Direct Viral Cell-to-Cell Spread of Human Cytomegalovirus. Virology 2013, 444, 140–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abela, I.A.; Berlinger, L.; Schanz, M.; Reynell, L.; Günthard, H.F.; Rusert, P.; Trkola, A. Cell-Cell Transmission Enables HIV-1 to Evade Inhibition by Potent CD4bs Directed Antibodies. PLoS Pathog. 2012, 8, e1002634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, C.J.A.; Williams, J.P.; Schiffner, T.; Gärtner, K.; Ochsenbauer, C.; Kappes, J.; Russell, R.A.; Frater, J.; Sattentau, Q.J. High-Multiplicity HIV-1 Infection and Neutralizing Antibody Evasion Mediated by the Macrophage-T Cell Virological Synapse. J. Virol. 2014, 88, 2025–2034. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, A.; Arndt, M.A.; Grosse-Hovest, L.; Weichert, W.; Giebel, B.; Dittmer, U.; Hengel, H.; Jager, D.; Schneweis, K.E.; Eis-Hubinger, A.M.; et al. Overcoming Drug-Resistant Herpes Simplex Virus (HSV) Infection by a Humanized Antibody. Proc. Natl. Acad. Sci. USA 2013, 110, 6760–6765. [Google Scholar] [CrossRef] [Green Version]
- Malbec, M.; Porrot, F.; Rua, R.; Horwitz, J.; Klein, F.; Halper-Stromberg, A.; Scheid, J.F.; Eden, C.; Mouquet, H.; Nussenzweig, M.C.; et al. Broadly Neutralizing Antibodies That Inhibit HIV-1 Cell to Cell Transmission. J. Exp. Med. 2013, 210, 2813–2821. [Google Scholar] [CrossRef] [Green Version]
- Schiffner, T.; Sattentau, Q.J.; Duncan, C.J.A. Cell-to-Cell Spread of HIV-1 and Evasion of Neutralizing Antibodies. Vaccine 2013, 31, 5789–5797. [Google Scholar] [CrossRef]
- Cui, X.; Lee, R.; Adler, S.P.; McVoy, M.A. Antibody Inhibition of Human Cytomegalovirus Spread in Epithelial Cell Cultures. J. Virol. Methods 2013, 192, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.J.; Adler, B.; Sampaio, K.L.; Digel, M.; Jahn, G.; Ettischer, N.; Stierhof, Y.-D.; Scrivano, L.; Koszinowski, U.; Mach, M.; et al. UL74 of Human Cytomegalovirus Contributes to Virus Release by Promoting Secondary Envelopment of Virions. J. Virol. 2008, 82, 2802–2812. [Google Scholar] [CrossRef] [Green Version]
- Laib Sampaio, K.; Stegmann, C.; Brizic, I.; Adler, B.; Stanton, R.J.; Sinzger, C. The Contribution of PUL74 to Growth of Human Cytomegalovirus Is Masked in the Presence of RL13 and UL128 Expression. J. Gen. Virol. 2016, 97, 1917–1927. [Google Scholar] [CrossRef] [Green Version]
- Soroceanu, L.; Akhavan, A.; Cobbs, C.S. Platelet-Derived Growth Factor-Alpha Receptor Activation Is Required for Human Cytomegalovirus Infection. Nature 2008, 455, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Chong, H.; Zhu, Y.; Yu, D.; He, Y. Structural and Functional Characterization of Membrane Fusion Inhibitors with Extremely Potent Activity against Human Immunodeficiency Virus Type 1 (HIV-1), HIV-2, and Simian Immunodeficiency Virus. J. Virol. 2018, 92, e01088-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dell’Oste, V.; Biolatti, M.; Galitska, G.; Griffante, G.; Gugliesi, F.; Pasquero, S.; Zingoni, A.; Cerboni, C.; De Andrea, M. Tuning the Orchestra: HCMV vs. Innate Immunity. Front. Microbiol. 2020, 11, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, M.; Stamminger, T. Emerging Role of PML Nuclear Bodies in Innate Immune Signaling. J. Virol. 2016, 90, 5850–5854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, E.-M.; Scherer, M.; Stamminger, T. Intrinsic Immune Mechanisms Restricting Human Cytomegalovirus Replication. Viruses 2021, 13, 179. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GT40 | GRHIYIYVPDPDVAFVPLGMTDYLVIVEDDDSAIIPCRTT |
| GT40/G1C_cyc | CRHIYIYVPDPDVAFVPLGMTDYLVIVEDDDSAIIPCRTT |
| GT40/G1C/D30C/C37S_cyc | CRHIYIYVPDPDVAFVPLGMTDYLVIVEDCDSAIIPSRTT |
| GT40/P9C/S32C/C37S_cyc | GRHIYIYVCDPDVAFVPLGMTDYLVIVEDDDCAIIPSRTT |
| LT-53_cyc | TCYYNHTQTEENELEGRHIYIYVPDPDVAFVPLGMTDYLVIVEDDDSAIIPCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braun, B.; Fischer, D.; Laib Sampaio, K.; Mezger, M.; Stöhr, D.; Stanton, R.J.; Sinzger, C. Peptide Derivatives of Platelet-Derived Growth Factor Receptor Alpha Inhibit Cell-Associated Spread of Human Cytomegalovirus. Viruses 2021, 13, 1780. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091780
Braun B, Fischer D, Laib Sampaio K, Mezger M, Stöhr D, Stanton RJ, Sinzger C. Peptide Derivatives of Platelet-Derived Growth Factor Receptor Alpha Inhibit Cell-Associated Spread of Human Cytomegalovirus. Viruses. 2021; 13(9):1780. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091780
Chicago/Turabian StyleBraun, Berenike, Dina Fischer, Kerstin Laib Sampaio, Maja Mezger, Dagmar Stöhr, Richard James Stanton, and Christian Sinzger. 2021. "Peptide Derivatives of Platelet-Derived Growth Factor Receptor Alpha Inhibit Cell-Associated Spread of Human Cytomegalovirus" Viruses 13, no. 9: 1780. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091780