
Exploring the Mechanisms of Recovery in Anorexia Nervosa through a Translational Approach: From Original Ecological Measurements in Human to Brain Tissue Analyses in Mice

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment 1: Clinical Assessment
2.2. Experiment 2: Preclinical Approach
2.2.1. Animals
2.2.2. Protocols
2.2.3. Euthanasia and Tissue Collection
2.2.4. Immunocytochemistry
2.2.5. Blood Assays
2.2.6. RT-PCR Extraction and Analysis
2.3. Statistical Analysis
3. Results
3.1. Experiment 1. Preliminary Data: Clinical Investigation of Partial Recovery
3.1.1. Clinical Characteristics: Acute AN versus Partially Recovered AN Patients
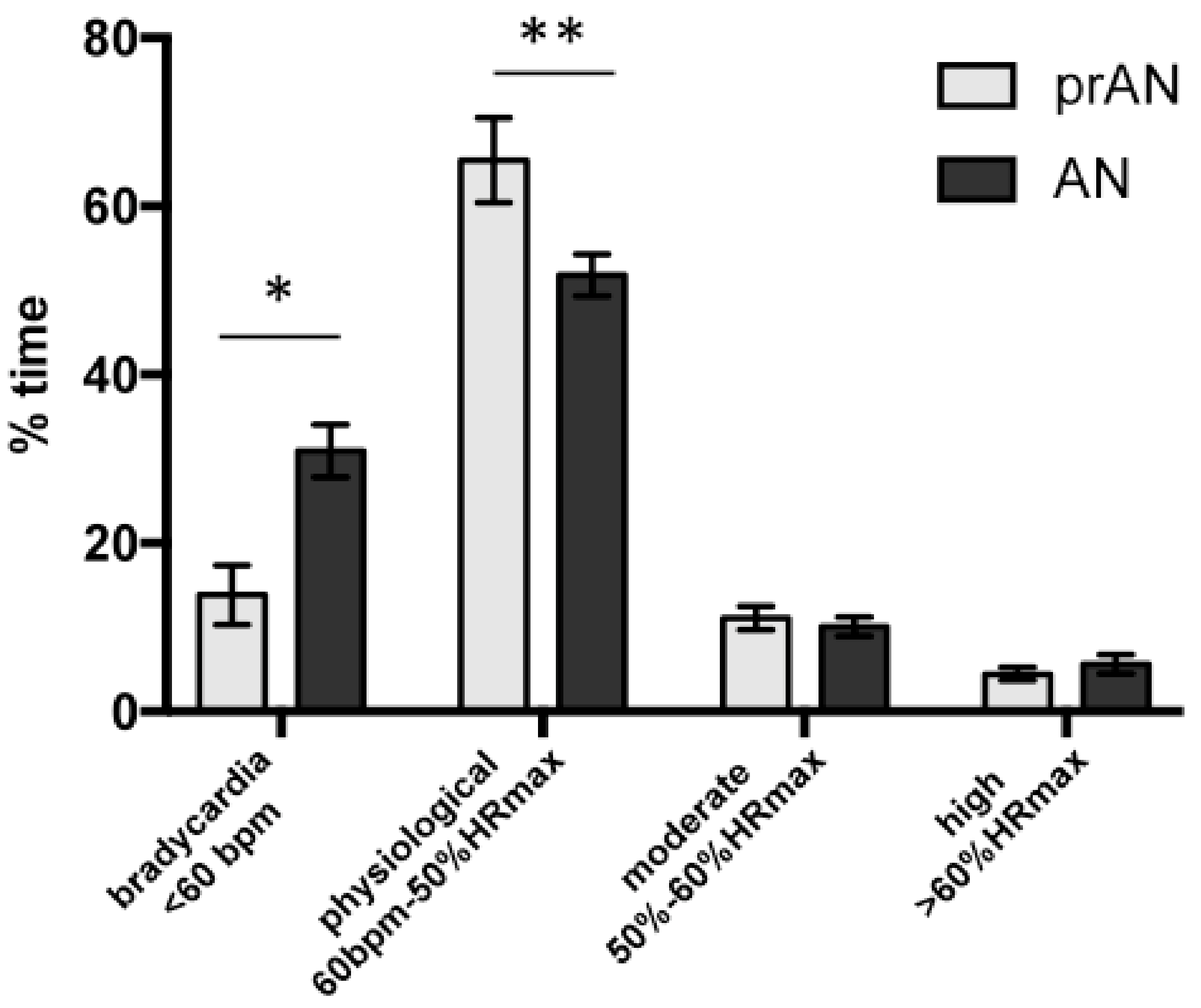
3.1.2. Continuous Cardiac Monitoring, Past and Actual BMI and Illness Duration
3.2. Experiment 2. Preclinical Investigation
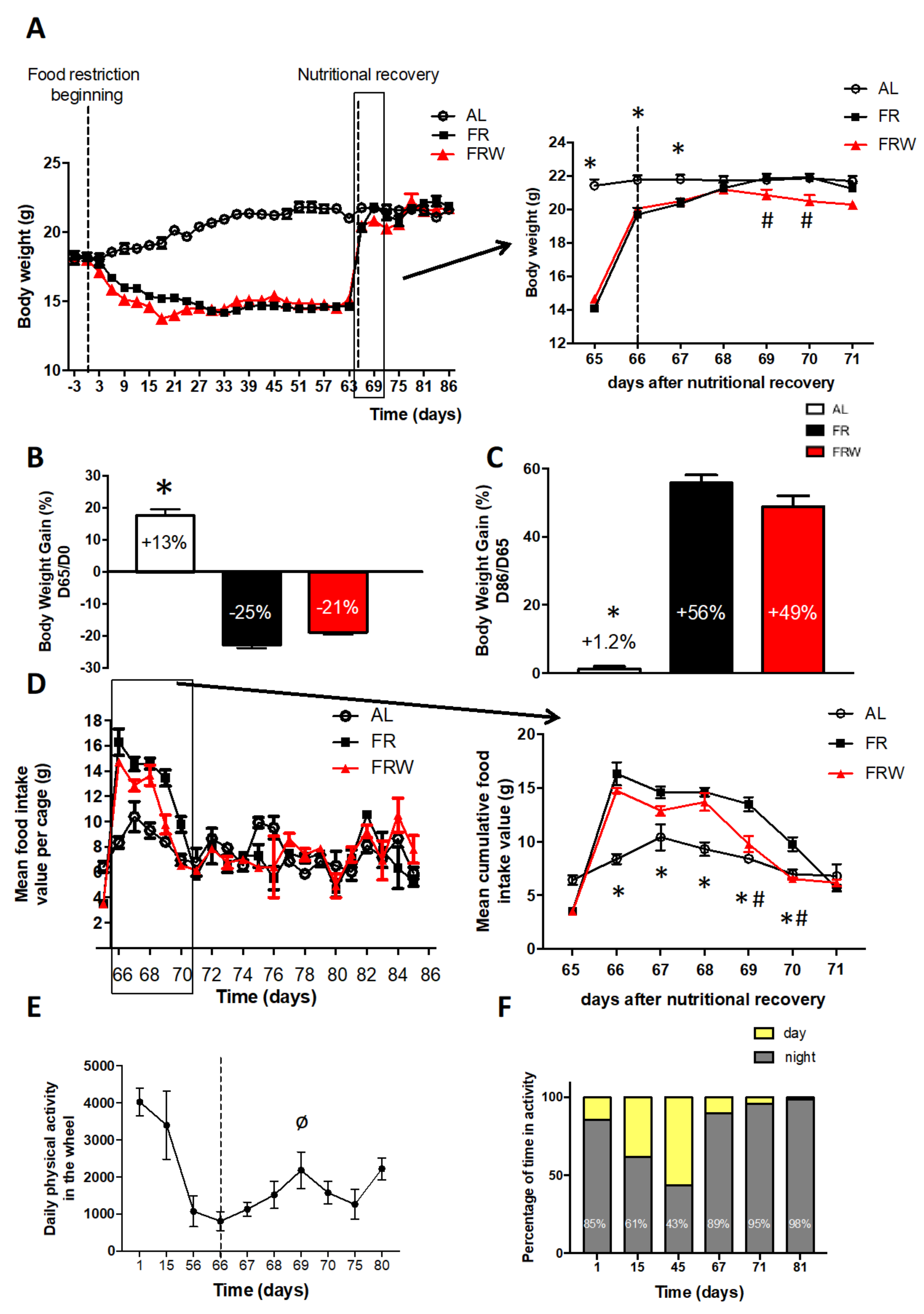
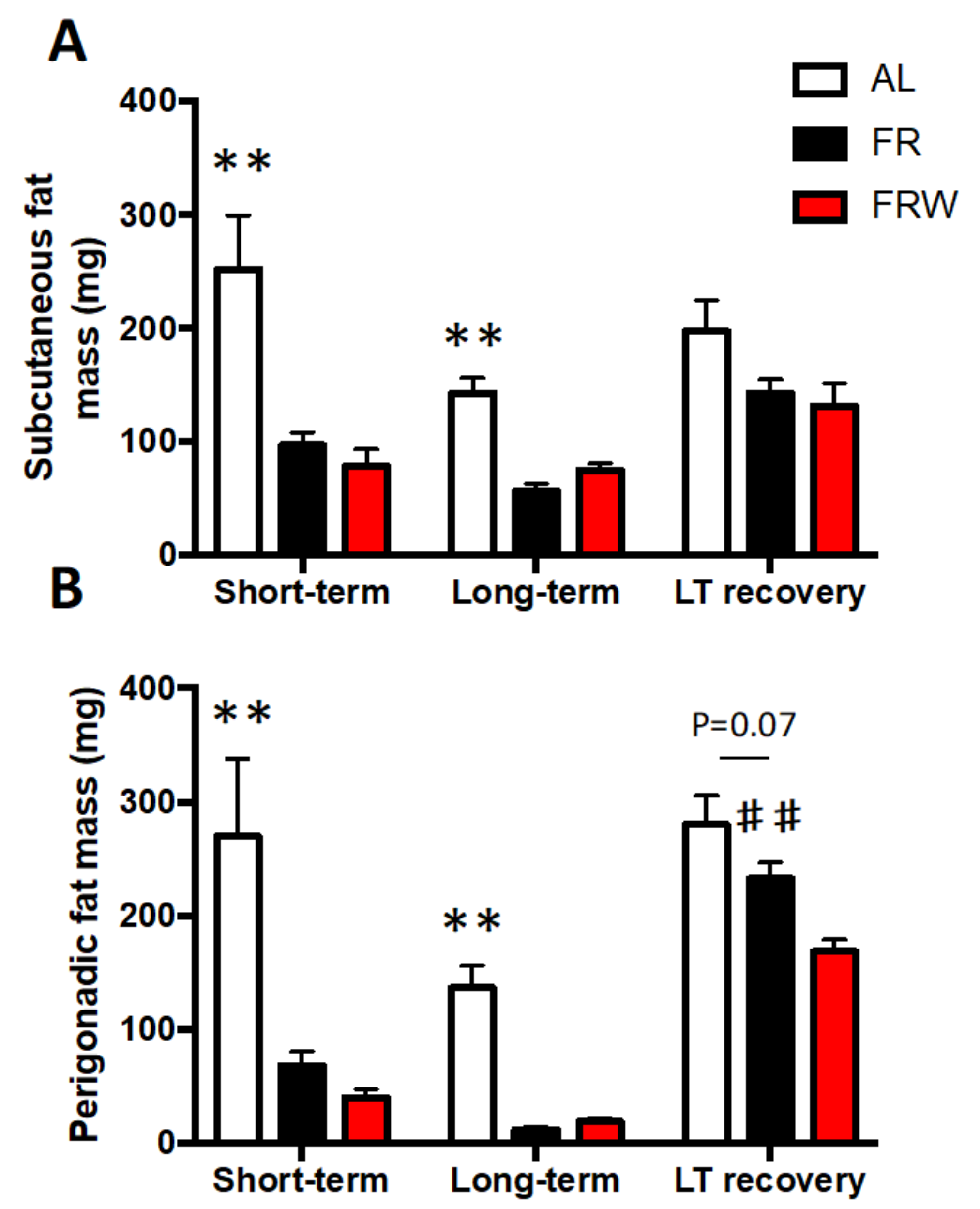
3.2.1. Body Weight, Food Intake, Physical Activity and Fat Mass Evolution
3.2.2. Ghrelin and Leptin Assays
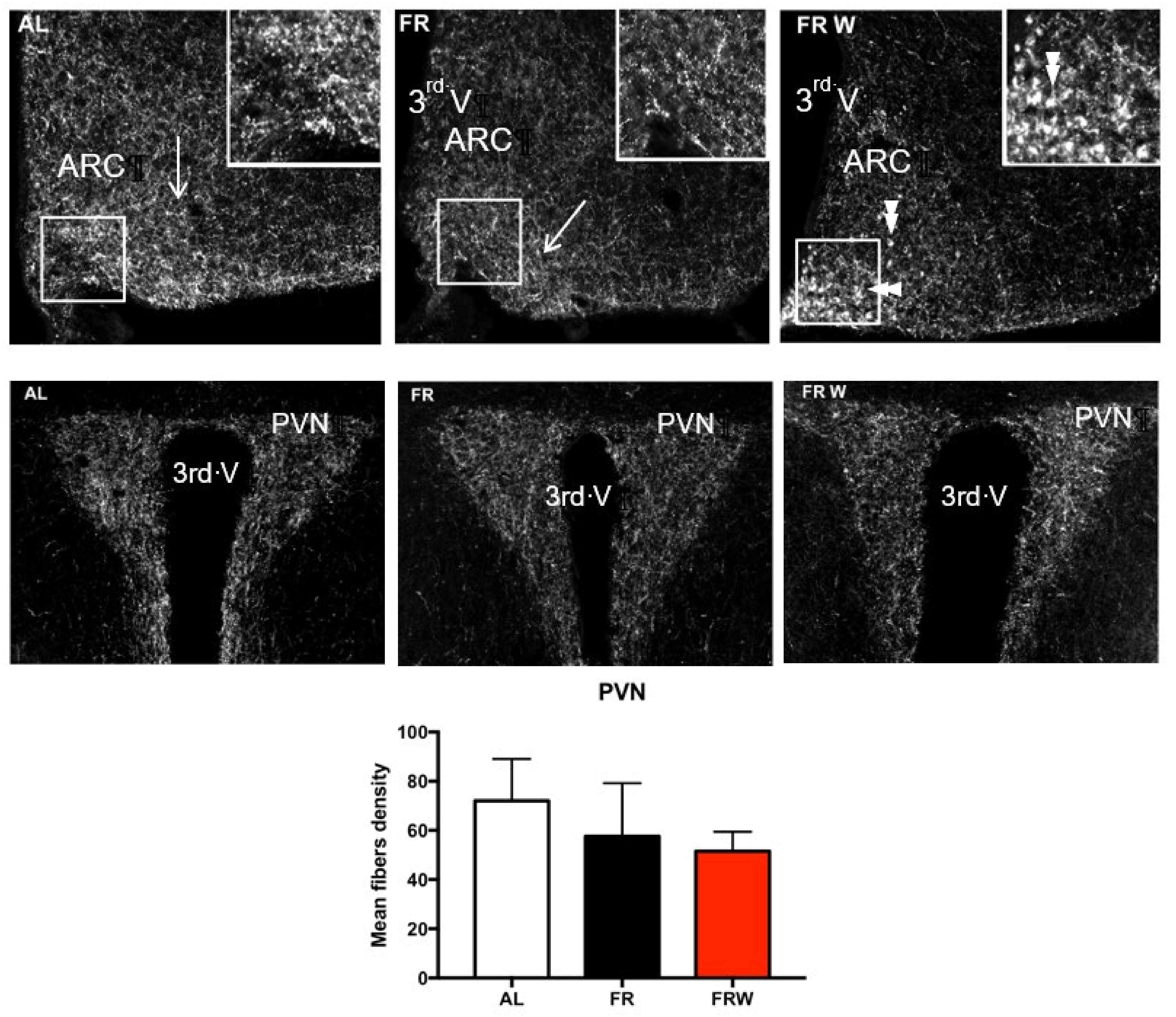
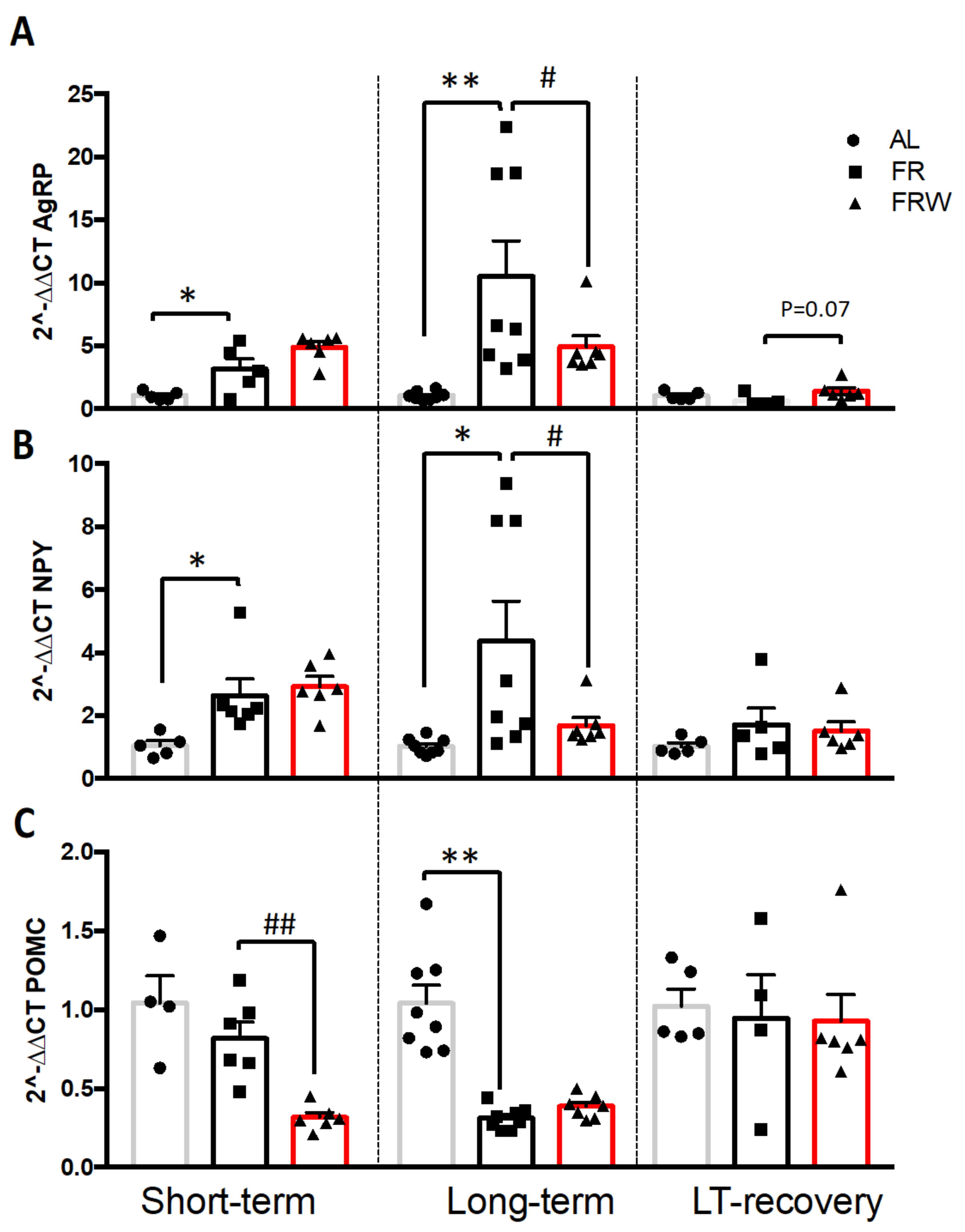
3.2.3. Immunocytochemical Detection of AgRP
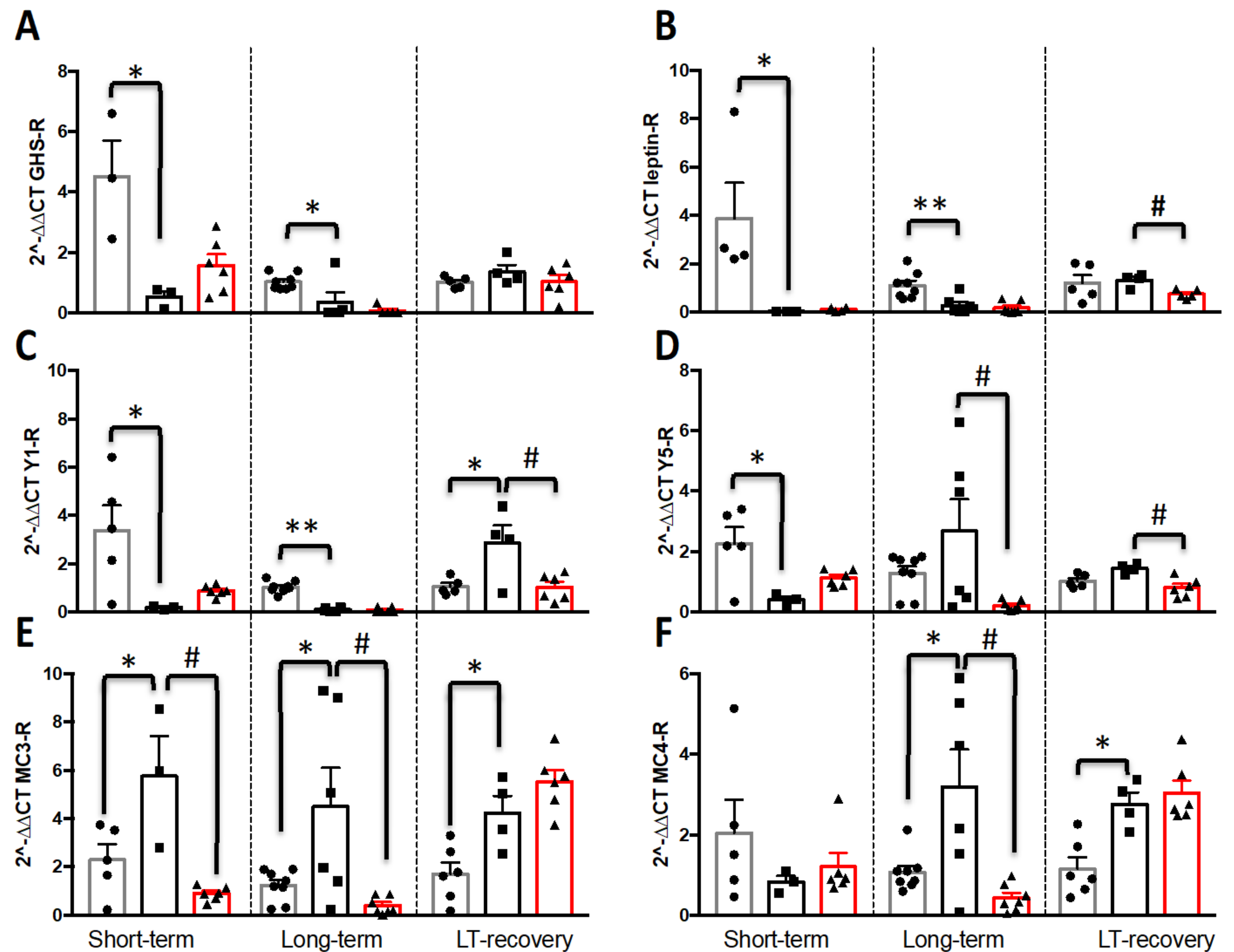
3.2.4. mRNA Expression of AgRP/NPY and Melanocortin Systems in the Hypothalamus
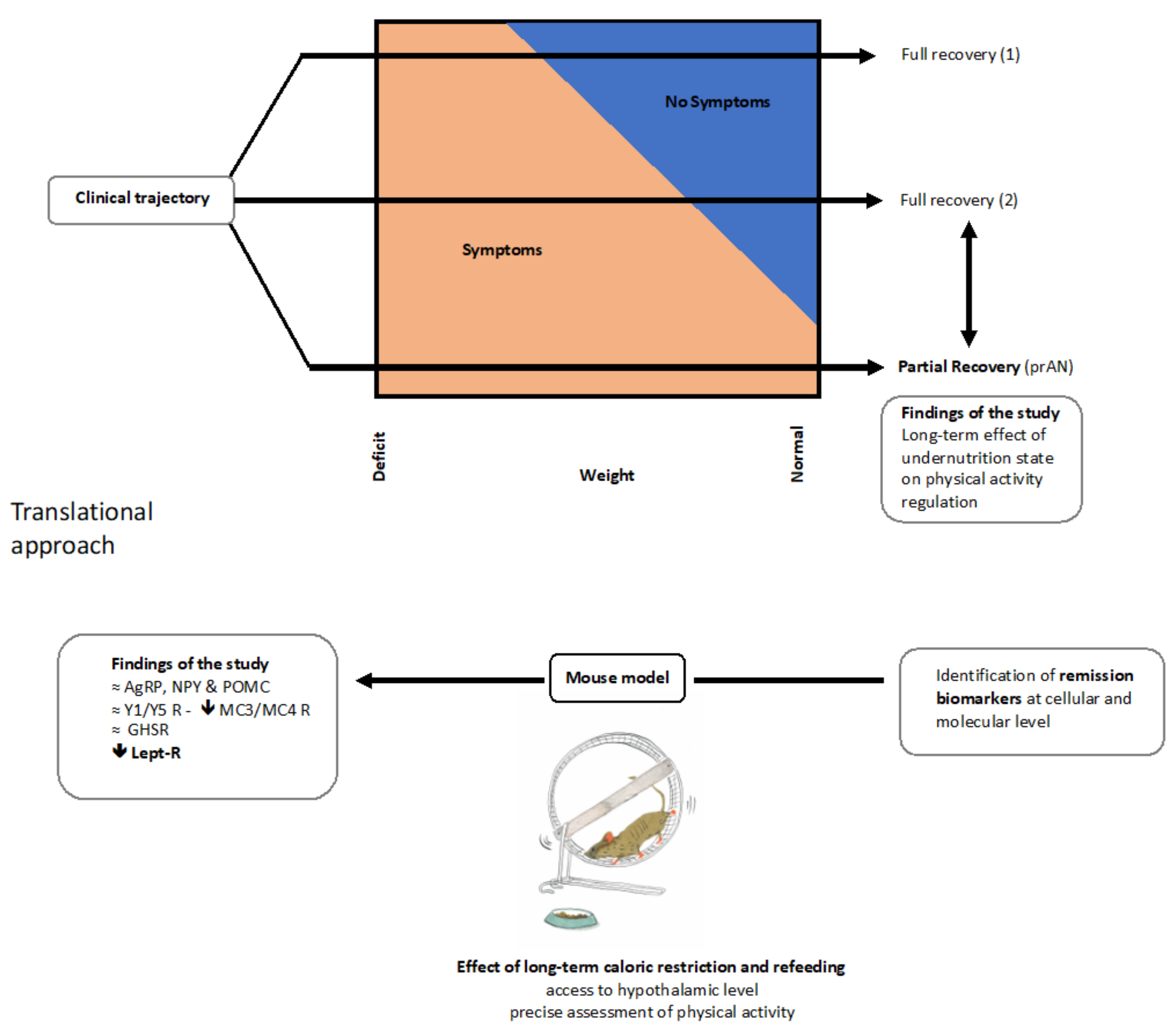
4. Discussion
4.1. Physical Activity in Acute AN and prAN
4.2. Impact of Moderate Physical Activity during Chronic Food Restriction on Recovery
4.3. Nutritional Recovery Differentially Impacts Hypothalamic Activity
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duncan, L.; Yilmaz, Z.; Gaspar, H.; Walters, R.; Goldstein, J.; Anttila, V.; Bulik-Sullivan, B.; Ripke, S.; Eating Disorders Working Group of the Psychiatric Genomics Consortium; Thornton, L.; et al. Significant Locus and Metabolic Genetic Correlations Revealed in Genome-Wide Association Study of Anorexia Nervosa. Am. J. Psychiatry 2017, 174, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Schorr, M.; Miller, K.K. The endocrine manifestations of anorexia nervosa: Mechanisms and management. Nat. Rev. Endocrinol. 2017, 13, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.J.; Yilmaz, Z.; Thornton, L.M.; Hübel, C.; Coleman, J.R.I.; Gaspar, H.A.; Bryois, J.; Hinney, A.; Leppä, V.M.; Mattheisen, M.; et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat. Genet. 2019, 51, 1207–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duriez, P.; Ramoz, N.; Gorwood, P.; Viltart, O.; Tolle, V. A Metabolic Perspective on Reward Abnormalities in Anorexia Nervosa. Trends Endocrinol. Metab. 2019, 30, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Viltart, O.; Duriez, P.; Tolle, V. Metabolic and neuroendocrine adaptations to undernutrition in anorexia nervosa: From a clinical to a basic research point of view. Horm. Mol. Biol. Clin. Investig. 2018, 36. [Google Scholar] [CrossRef]
- Holtkamp, K.; Hebebrand, J.; Herpertz-Dahlmann, B. The contribution of anxiety and food restriction on physical activity levels in acute anorexia nervosa. Int. J. Eat. Disord. 2004, 36, 163–171. [Google Scholar] [CrossRef]
- Duclos, M.; Ouerdani, A.; Mormede, P.; Konsman, J.P. Food restriction-induced hyperactivity: Addiction or adaptation to famine? Psychoneuroendocrinology 2013, 38, 884–897. [Google Scholar] [CrossRef]
- Pleplé, A.; Lalanne, C.; Huas, C.; Mattar, L.; Hanachi, M.; Flament, M.F.; Carchon, I.; Jouen, F.; Berthoz, S.; Godart, N. Nutritional status and anxious and depressive symptoms in anorexia nervosa: A prospective study. Sci. Rep. 2021, 11, 771. [Google Scholar] [CrossRef]
- Van Furth, E.F.; van der Meer, A.; Cowan, K. Top 10 research priorities for eating disorders. Lancet Psychiatry 2016, 3, 706–707. [Google Scholar] [CrossRef]
- Strober, M.; Freeman, R.; Morrell, W. The Long-Term Course of Severe Anorexia Nervosa in Adolescents: Survival Analysis of Recovery, Relapse, and Outcome Predictors over 10–15 Years in a Prospective Study. Int. J. Eat. Disord. 1997, 22, 339–360. [Google Scholar] [CrossRef]
- Treasure, J.; Zipfel, S.; Micali, N.; Wade, T.; Stice, E.; Claudino, A.; Schmidt, U.; Frank, G.K.; Bulik, C.M.; Wentz, E. Anorexia nervosa. Nat. Rev. Dis. Primers 2015, 1, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Muratore, A.F.; Attia, E. Current Therapeutic Approaches to Anorexia Nervosa: State of the Art. Clin. Ther. 2021, 43, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Eddy, K.T.; Dorer, D.J.; Franko, D.L.; Tahilani, K.; Thompson-Brenner, H.; Herzog, D.B. Diagnostic Crossover in Anorexia Nervosa and Bulimia Nervosa: Implications for DSM-V. Am. J. Psychiatry 2008, 165, 245–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, Y.; Arai, T.; Okuda, Y.; Yamashita, K. The Regulation of obese(ob) Gene Expression by Intracellular Fatty Acid Concentration in Adipocytes. Intern. Med. 1999, 38, 213–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fried, S.K.; Ricci, M.R.; Russell, C.D.; Laferrère, B. Regulation of Leptin Production in Humans. J. Nutr. 2000, 130, 3127S–3131S. [Google Scholar] [CrossRef]
- Peelman, F.; Zabeau, L.; Moharana, K.; Savvides, S.N.; Tavernier, J. 20 YEARS OF LEPTIN: Insights into signaling assemblies of the leptin receptor. J. Endocrinol. 2014, 223, T9–T23. [Google Scholar] [CrossRef]
- Fernandes, M.F.A.; Matthys, D.; Hryhorczuk, C.; Sharma, S.; Mogra, S.; Alquier, T.; Fulton, S. Leptin Suppresses the Rewarding Effects of Running via STAT3 Signaling in Dopamine Neurons. Cell Metab. 2015, 22, 741–749. [Google Scholar] [CrossRef] [Green Version]
- Exner, C.; Hebebrand, J.; Remschmidt, H.; Wewetzer, C.; Ziegler, A.; Herpertz, S.; Schweiger, U.; Blum, W.F.; Preibisch, G.; Heldmaier, G.; et al. Leptin suppresses semi-starvation induced hyperactivity in rats: Implications for anorexia nervosa. Mol. Psychiatry 2000, 5, 476–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Date, Y.; Kojima, M.; Hosoda, H.; Sawaguchi, A.; Mondal, M.S.; Suganuma, T.; Matsukura, S.; Kangawa, K.; Nakazato, M. Ghrelin, a Novel Growth Hormone-Releasing Acylated Peptide, Is Synthesized in a Distinct Endocrine Cell Type in the Gastrointestinal Tracts of Rats and Humans. Endocrinology 2000, 141, 4255–4261. [Google Scholar] [CrossRef]
- Müller, T.; Nogueiras, R.; Andermann, M.; Andrews, Z.B.; Anker, S.; Argente, J.; Batterham, R.; Benoit, S.; Bowers, C.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef]
- Méquinion, M.; Langlet, F.; Zgheib, S.; Dickson, S.; Dehouck, B.; Chauveau, C.; Viltart, O. Ghrelin: Central and Peripheral Implications in Anorexia Nervosa. Front. Endocrinol. 2013, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Labarthe, A.; Fiquet, O.; Hassouna, R.; Zizzari, P.; Lanfumey, L.; Ramoz, N.; Grouselle, D.; Epelbaum, J.; Tolle, V. Ghrelin-Derived Peptides: A Link between Appetite/Reward, GH Axis, and Psychiatric Disorders? Front. Endocrinol. 2014, 5, 163. [Google Scholar] [CrossRef] [Green Version]
- Ralevski, E.; Shanabrough, M.; Newcomb, J.; Gandelman, E.; Hayden, R.; Horvath, T.L.; Petrakis, I. Ghrelin is Related to Personality Differences in Reward Sensitivity and Impulsivity. Alcohol Alcohol. 2018, 53, 52–56. [Google Scholar] [CrossRef]
- Bake, T.; Edvardsson, C.E.; Cummings, C.J.; Dickson, S.L. Ghrelin’s effects on food motivation in rats are not limited to palatable foods. J. Neuroendocrinol. 2019, 31, e12665. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, H.; Kojima, M.; Matsuo, H.; Kangawa, K. Ghrelin and Des-acyl Ghrelin: Two Major Forms of Rat Ghrelin Peptide in Gastrointestinal Tissue. Biochem. Biophys. Res. Commun. 2000, 279, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, A.; Inui, A.; Fujimiya, M.; Sakamaki, R.; Shinfuku, N.; Ueta, Y.; Meguid, M.M.; Kasuga, M. Stomach regulates energy balance via acylated ghrelin and desacyl ghrelin. Gut 2005, 54, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Gortan Cappellari, G.; Barazzoni, R. Ghrelin forms in the modulation of energy balance and metabolism. Eat. Weight. Disord. 2019, 24, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Dardennes, R.; Tolle, V.; Lavoisy, G.; Grouselle, D.; Alanbar, N.; Duriez, P.; Gorwood, P.; Ramoz, N.; Epelbaum, J. Lower leptin level at discharge in acute anorexia nervosa is associated with early weight-loss. Eur. Eat. Disord. Rev. 2021, 29, 634–644. [Google Scholar] [CrossRef]
- Milos, G.; Antel, J.; Kaufmann, L.-K.; Barth, N.; Koller, A.; Tan, S.; Wiesing, U.; Hinney, A.; Libuda, L.; Wabitsch, M.; et al. Short-term metreleptin treatment of patients with anorexia nervosa: Rapid on-set of beneficial cognitive, emotional, and behavioral effects. Transl. Psychiatry 2020, 10, 303. [Google Scholar] [CrossRef]
- Seidel, M.; Markmann Jensen, S.; Healy, D.; Dureja, A.; Watson, H.J.; Holst, B.; Bulik, C.M.; Sjögren, J.M. A Systematic Review and Meta-Analysis Finds Increased Blood Levels of All Forms of Ghrelin in Both Restricting and Binge-Eating/Purging Subtypes of Anorexia Nervosa. Nutrients 2021, 13, 709. [Google Scholar] [CrossRef]
- Duriez, P.; Robichon, L.; Dardennes, R.; Lavoisy, G.; Grouselle, D.; Epelbaum, J.; Ramoz, N.; Gorwood, P.; Tolle, V.; Viltart, O. Unexpected Association of Desacyl-Ghrelin with Physical Activity and Chronic Food Restriction: A Translational Study on Anorexia Nervosa. J. Clin. Med. 2020, 9, 2782. [Google Scholar] [CrossRef]
- Meyer, C.; Taranis, L.; Touyz, S. Excessive exercise in the eating disorders: A need for less activity from patients and more from researchers. Eur. Eat. Disord. Rev. 2008, 16, 81–83. [Google Scholar] [CrossRef]
- Krashes, M.J.; Koda, S.; Ye, C.; Rogan, S.C.; Adams, A.C.; Cusher, D.S.; Maratos-Flier, E.; Roth, B.L.; Lowell, B.B. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J. Clin. Investig. 2011, 121, 1424–1428. [Google Scholar] [CrossRef] [Green Version]
- Betley, J.N.; Cao, Z.F.H.; Ritola, K.D.; Sternson, S.M. Parallel, Redundant Circuit Organization for Homeostatic Control of Feeding Behavior. Cell 2013, 155, 1337–1350. [Google Scholar] [CrossRef] [Green Version]
- Zhan, C.; Zhou, J.; Feng, Q.; Zhang, J.-E.; Lin, S.; Bao, J.; Wu, P.; Luo, M. Acute and Long-Term Suppression of Feeding Behavior by POMC Neurons in the Brainstem and Hypothalamus, Respectively. J. Neurosci. 2013, 33, 3624–3632. [Google Scholar] [CrossRef]
- Bunner, W.; Landry, T.; Laing, B.T.; Li, P.; Rao, Z.; Yuan, Y.; Huang, H. ARCAgRP/NPY Neuron Activity Is Required for Acute Exercise-Induced Food Intake in Un-Trained Mice. Front. Physiol. 2020, 11, 411. [Google Scholar] [CrossRef]
- Williams, K.W.; Elmquist, J.K. From neuroanatomy to behavior: Central integration of peripheral signals regulating feeding behavior. Nat. Neurosci. 2012, 15, 1350–1355. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, M.O.; Bober, J.; Ferreira, J.G.; Tellez, L.A.; Mineur, Y.S.; Souza, D.O.; Gao, X.-B.; Picciotto, M.R.; Araújo, I.; Liu, Z.-W.; et al. AgRP neurons regulate development of dopamine neuronal plasticity and nonfood-associated behaviors. Nat. Neurosci. 2012, 15, 1108–1110. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, M.O.; Zimmer, M.R.; Bober, J.; Horvath, T.L. Hypothalamic Agrp Neurons Drive Stereotypic Behaviors beyond Feeding. Cell 2015, 160, 1222–1232. [Google Scholar] [CrossRef] [Green Version]
- Denis, R.G.P.; Joly-Amado, A.; Webber, E.; Langlet, F.; Schaeffer, M.; Padilla, S.L.; Cansell, C.; Dehouck, B.; Castel, J.; Delbès, A.-S.; et al. Palatability Can Drive Feeding Independent of AgRP Neurons. Cell Metab. 2017, 25, 975. [Google Scholar] [CrossRef] [Green Version]
- Miletta, M.C.; Iyilikci, O.; Shanabrough, M.; Šestan-Peša, M.; Cammisa, A.; Zeiss, C.J.; Dietrich, M.O.; Horvath, T.L. AgRP neurons control compulsive exercise and survival in an activity-based anorexia model. Nat. Metab. 2020, 2, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Gao, Y.; Alhadeff, A.L.; Castorena, C.M.; Huang, Y.; Lieu, L.; Afrin, S.; Sun, J.; Betley, J.N.; Guo, H.; et al. Cellular and synaptic reorganization of arcuate NPY/AgRP and POMC neurons after exercise. Mol. Metab. 2018, 18, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Li, H.E.; Rossi, M.A.; Watson, G.D.R.; Moore, H.G.; Cai, M.T.; Kim, N.; Vokt, K.A.; Lu, D.; Bartholomew, R.A.; Hughes, R.N.; et al. Hypothalamic-Extended Amygdala Circuit Regulates Temporal Discounting. J. Neurosci. 2021, 41, 1928–1940. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hou, Y.; Zhang, J.; Sui, G.; Du, X.; Licinio, J.; Wong, M.-L.; Yang, Y. AGRP neurons modulate fasting-induced anxiolytic effects. Transl. Psychiatry 2019, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, M.; Langlet, F.; Lafont, C.; Molino, F.; Hodson, D.J.; Roux, T.; Lamarque, L.; Verdié, P.; Bourrier, E.; Dehouck, B.; et al. Rapid sensing of circulating ghrelin by hypothalamic appetite-modifying neurons. Proc. Natl. Acad. Sci. USA 2013, 110, 1512–1517. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Liu, C.; Uchida, A.; Chuang, J.-C.; Walker, A.; Liu, T.; Osborne-Lawrence, S.; Mason, B.L.; Mosher, C.; Berglund, E.D.; et al. Arcuate AgRP neurons mediate orexigenic and glucoregulatory actions of ghrelin. Mol. Metab. 2014, 3, 64–72. [Google Scholar] [CrossRef]
- Florent, V.; Baroncini, M.; Jissendi-Tchofo, P.; Lopes, R.; Vanhoutte, M.; Rasika, S.; Pruvo, J.-P.; Vignau, J.; Verdun, S.; Johansen, J.E.; et al. Hypothalamic Structural and Functional Imbalances in Anorexia Nervosa. Neuroendocrinology 2020, 110, 552–562. [Google Scholar] [CrossRef]
- Méquinion, M.; Caron, E.; Zgheib, S.; Stievenard, A.; Zizzari, P.; Tolle, V.; Cortet, B.; Lucas, S.; Prevot, V.; Chauveau, C.; et al. Physical activity: Benefit or weakness in metabolic adaptations in a mouse model of chronic food restriction? Am. J. Physiol. Endocrinol. Metab. 2015, 308, E241–E255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méquinion, M.; Chauveau, C.; Viltart, O. The Use of Animal Models to Decipher Physiological and Neurobiological Alterations of Anorexia Nervosa Patients. Front. Endocrinol. 2015, 6, 68. [Google Scholar] [CrossRef] [Green Version]
- Scharner, S.; Stengel, A. Animal Models for Anorexia Nervosa—A Systematic Review. Front. Hum. Neurosci. 2020, 14, 596381. [Google Scholar] [CrossRef] [PubMed]
- Duriez, P.; Eddarkaoui, S.; Blum, D.; Dickson, S.L.; Gorwood, P.; Tolle, V.; Viltart, O. Does physical activity associated with chronic food restriction alleviate anxiety like behaviour, in female mice? Horm. Behav. 2020, 124, 104807. [Google Scholar] [CrossRef]
- Verhagen, L.A.W.; Egecioglu, E.; Luijendijk, M.C.M.; Hillebrand, J.J.G.; Adan, R.A.H.; Dickson, S.L. Acute and chronic suppression of the central ghrelin signaling system reveals a role in food anticipatory activity. Eur. Neuropsychopharmacol. 2011, 21, 384–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, TX, USA, 2013; ISBN 978-0-89042-555-8. [Google Scholar]
- Garner, D.M. Eating Disorder Inventory—2nd Manual; Psychological Assessment Resources: Odessa, FL, USA, 1991. [Google Scholar]
- Zigmond, A.S.; Snaith, R.P. The Hospital Anxiety and Depression Scale. Acta Psychiatr. Scand. 1983, 67, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, W.K.; Price, L.H.; Rasmussen, S.A.; Mazure, C.; Fleischmann, R.L.; Hill, C.L.; Heninger, G.R.; Charney, D.S. The Yale-Brown Obsessive Compulsive Scale. I. Development, Use, and Reliability. Arch. Gen. Psychiatry 1989, 46, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate–phenol–chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Ehrlich, S.; Tam, F.I. Editorial: Refeeding in Anorexia Nervosa: Quo Vadis? J. Am. Acad. Child. Adolesc. Psychiatry 2021, 60, 566–567. [Google Scholar] [CrossRef]
- Redgrave, G.W.; Schreyer, C.C.; Coughlin, J.W.; Fischer, L.K.; Pletch, A.; Guarda, A.S. Discharge Body Mass Index, Not Illness Chronicity, Predicts 6-Month Weight Outcome in Patients Hospitalized With Anorexia Nervosa. Front. Psychiatry 2021, 12, 641861. [Google Scholar] [CrossRef]
- Giles, E.D.; Hagman, J.; Pan, Z.; MacLean, P.S.; Higgins, J.A. Weight restoration on a high carbohydrate refeeding diet promotes rapid weight regain and hepatic lipid accumulation in female anorexic rats. Nutr. Metab. 2016, 13, 18. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, K.; Herpertz-Dahlmann, B.; Hebebrand, K.; Mika, C.; Kratzsch, J.; Hebebrand, J. Physical Activity and Restlessness Correlate with Leptin Levels in Patients with Adolescent Anorexia Nervosa. Biol. Psychiatry 2006, 60, 311–313. [Google Scholar] [CrossRef]
- Van Elburg, A.A.; Kas, M.J.H.; Hillebrand, J.J.G.; Eijkemans, R.J.C.; Van Engeland, H. The impact of hyperactivity and leptin on recovery from anorexia nervosa. J. Neural Transm. 2007, 114, 1233–1237. [Google Scholar] [CrossRef] [Green Version]
- Broberger, C.; Johansen, J.; Johansson, C.; Schalling, M.; Hokfelt, T. The neuropeptide Y/agouti gene-related protein (AGRP) brain circuitry in normal, anorectic, and monosodium glutamate-treated mice. Proc. Natl. Acad. Sci. USA 1998, 95, 15043–15048. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, I.; Lindfors, C.; Fetissov, S.O.; Hökfelt, T.; Johansen, J.E. Aberrant agouti-related protein system in the hypothalamus of theanx/anx mouse is associated with activation of microglia. J. Comp. Neurol. 2008, 507, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- De Rijke, C.E.; Hillebrand, J.J.G.; Verhagen, L.A.W.; Roeling, T.A.P.; Adan, R.A.H. Hypothalamic neuropeptide expression following chronic food restriction in sedentary and wheel-running rats. J. Mol. Endocrinol. 2005, 35, 381–390. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Wilson, B.D.; Yang, Y.K.; Kerns, J.A.; Chen, Y.; Gantz, I.; Barsh, G.S. Antagonism of Central Melanocortin Receptors in Vitro and in Vivo by Agouti-Related Protein. Science 1997, 278, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Kopp, J.; Xu, Z.-Q.; Zhang, X.; Pedrazzini, T.; Herzog, H.; Kresse, A.; Wong, H.; Walsh, J.H.; Hökfelt, T. Expression of the neuropeptide Y Y1 receptor in the CNS of rat and of wild-type and Y1 receptor knock-out mice. Focus on immunohistochemical localization. Neuroscience 2002, 111, 443–532. [Google Scholar] [CrossRef]
- Morin, S.M.; Gehlert, D.R. Distribution of NPY Y5-Like Immunoreactivity in the Rat Brain. J. Mol. Neurosci. 2006, 29, 109–114. [Google Scholar] [CrossRef]
- Mercer, R.E.; Chee, M.J.S.; Colmers, W.F. The role of NPY in hypothalamic mediated food intake. Front. Neuroendocr. 2011, 32, 398–415. [Google Scholar] [CrossRef]
- Acuna-Goycolea, C.; Tamamaki, N.; Yanagawa, Y.; Obata, K.; van den Pol, A.N. Mechanisms of Neuropeptide Y, Peptide YY, and Pancreatic Polypeptide Inhibition of Identified Green Fluorescent Protein-Expressing GABA Neurons in the Hypothalamic Neuroendocrine Arcuate Nucleus. J. Neurosci. 2005, 25, 7406–7419. [Google Scholar] [CrossRef] [Green Version]
- Galas, L.; Tonon, M.-C.; Beaujean, D.; Fredriksson, R.; Larhammar, D.; Lihrmann, I.; Jégou, S.; Fournier, A.; Chartrel, N.; Vaudry, H. Neuropeptide Y Inhibits Spontaneous Alpha-Melanocyte-Stimulating Hormone (Alpha-MSH) Release via a Y(5) Receptor and Suppresses Thyrotropin-Releasing Hormone-Induced Alpha-MSH Secretion via a Y(1) Receptor in Frog Melanotrope Cells. Endocrinology 2002, 143, 1686–1694. [Google Scholar] [CrossRef]
- Méquinion, M.; Le Thuc, O.; Zgheib, S.; Alexandre, D.; Chartrel, N.; Rovère, C.; Hardouin, P.; Viltart, O.; Chauveau, C. Long-Term Energy Deficit in Mice Causes Long-Lasting Hypothalamic Alterations after Recovery. Neuroendocrinology 2017, 105, 372–383. [Google Scholar] [CrossRef]
- Iyengar, S.; Li, D.L.; Simmons, R.M. Characterization of neuropeptide Y-induced feeding in mice: Do Y1-Y6 receptor subtypes mediate feeding? J. Pharmacol. Exp. Ther. 1999, 289, 1031–1040. [Google Scholar] [PubMed]
- Nguyen, A.D.; Mitchell, N.F.; Lin, S.; Macia, L.; Yulyaningsih, E.; Baldock, P.A.; Enriquez, R.F.; Zhang, L.; Shi, Y.-C.; Zolotukhin, S.; et al. Y1 and Y5 Receptors Are Both Required for the Regulation of Food Intake and Energy Homeostasis in Mice. PLoS ONE 2012, 7, e40191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Hersch, J.; Bodell, L.P.; Schebendach, J.; Hildebrandt, T.; Walsh, B.T.; Mayer, L.E.S. The association between leptin and weight maintenance outcome in anorexia nervosa. Int. J. Eat. Disord. 2021, 54, 527–534. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Reference | Forward Primer | Reverse Primer |
|---|---|---|---|
| AgRP | U89486.1 | 5′-CCCAGAGTTCCCAGGTCTAAGTCT-3′ | 5′-CACCTCCGCCAAAGCTTCT-3′ |
| NPY | NM_023456.2 | 5′-CCGCTCTGCGACACTACAT-3′ | 5′-TGTCTCAGGGCTGGATCTCT-3′ |
| POMC | BC061215.1 | 5′-AGTGCCAGGACCTCACCA-3′ | 5′-CAGCGAGAGGTCGAGTTTG-3′ |
| MC3R | NM_008561 | 5′-TCCGATGCTGCCTAACCTCT-3′ | 5′-GGATGTTTTCCATCAGACTGACG-3′ |
| MC4R | NM_016977 | 5′-CCCGGACGGAGGATGCTAT-3′ | 5′-TCGCCACGATCACTAGAATGT-3′ |
| Y1R | NM_010934 | 5′-TGATCTCCACCTGCGTCAAC-3′ | 5′-ATGGCTATGGTCTCGTAGTCAT-3′ |
| Y2R | NM_008731 | 5′-GCCAGGGCACACTACTCCTA-3′ | 5′-CTACCCCTAGCAAGATGATGGA-3′ |
| Y5R | NM_016708.3 | 5′-CATCTCAAGCAGAAGCGACC-3′ | 5′-CTCCATACTAGAGTCCTCGGG-3′ |
| Lept-R | NM_010704.2 | 5′-GTCTTCGGGGATGTGAATGTC-3′ | 5′-ACCTAAGGGTGGATCGGGTTT-3′ |
| GHSR | NM_177330 | 5′-TGGAGATCGCGCAGATCAG-3′ | 5′CCGGGAACTCTCATCCTTCAG-3′ |
| GADPH | GU214026 | 5′-GAACATCATCCCTGCATCC-3′ | 5′-CCAGTGAGCTTCCCGTTCA-3′ |
| AN (n = 225) | prAN (n = 41) | AN vs. prAN | ||||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | U | p | |
| Age | 27.8 | 9.76 | 27.2 | 8.43 | 4558 | 0.904 |
| BMI | 15.1 | 1.19 | 20.5 | 1.81 | 0 | <0.001 |
| BMI min | 13.4 | 3.07 | 15 | 1.34 | 1956 | <0.001 |
| Illness duration | 8.9 | 8.8 | 10.3 | 8.6 | 3274 | 0.199 |
| EDI | ||||||
| Drive for thinness | 10.5 | 6.95 | 13.8 | 6.33 | 3030 | 0.004 |
| Bulimia | 3.65 | 5.35 | 5.92 | 6.37 | 3290 | 0.017 |
| Body dissatisfaction | 12.1 | 7.16 | 16.9 | 8.05 | 2750 | <0.001 |
| Ineffectiveness | 11.6 | 7.57 | 12.7 | 6.52 | 3890 | 0.383 |
| Perfectionism | 7.52 | 4.76 | 9.15 | 4.51 | 3407 | 0.044 |
| Interpersonal distrust | 6.29 | 4.59 | 6 | 4.56 | 4016 | 0.564 |
| Interoceptive awareness | 10.5 | 6.9 | 13.7 | 7.05 | 3143 | 0.008 |
| Maturity fears | 6.87 | 5.95 | 7.35 | 6.22 | 4040 | 0.604 |
| Ascetism | 7.73 | 4.95 | 8.68 | 4.49 | 3666 | 0.161 |
| Emotional dysregulation | 5.44 | 5.49 | 8.25 | 6.72 | 3185 | 0.011 |
| Social insecurity | 7.84 | 5.23 | 8.4 | 4.26 | 3904 | 0.402 |
| Total | 89.62 | 46.57 | 110.97 | 41.14 | 3133 | 0.007 |
| HADS Anxiety | 12.5 | 4.33 | 13.9 | 3.45 | 2969 | 0.069 |
| HADS Depression | 8.59 | 4.53 | 8.29 | 4.34 | 3485 | 0.663 |
| YBS Compulsion | 7.84 | 5.37 | 8.74 | 4.27 | 3410 | 0.525 |
| YBS Obsession | 8.93 | 5.01 | 10.6 | 4.16 | 3061 | 0.117 |
| Ferritin (ng/mL) | 81 | 75.1 | 30.8 | 17 | 845 | <0.001 |
| Albumin (g/L) | 47.7 | 6.21 | 43.7 | 8.73 | 1670 | <0.001 |
| AN (n = 266) vs. AN with Cardiac Monitoring (n = 68) | aAN (n = 225) vs. aAN with Cardiac Monitoring (n = 55) | prAN (n = 41) vs. prAN with Cardiac Monitoring (n = 13) | ||||
|---|---|---|---|---|---|---|
| U | p | U | p | U | P | |
| Age | 7656 | 0.108 | 5030 | 0.050 | 218 | 0.559 |
| BMI | 7786 | 0.110 | 5256 | 0.124 | 209 | 0.248 |
| BMI min | 6640 | 0.371 | 4258 | 0.338 | 265 | 0.984 |
| Illness duration | 6721 | 0.032 | 4517 | 0.034 | 220 | 0.759 |
| EDI | ||||||
| Drive for thinness | 7989 | 0.908 | 5456 | 0.910 | 224 | 0.727 |
| Bulimia | 7936 | 0.889 | 5408 | 0.825 | 232 | 0.860 |
| Body dissatisfaction | 7483 | 0.373 | 4968 | 0.270 | 238 | 0.965 |
| Ineffectiveness | 7927 | 0.834 | 5408 | 0.834 | 237 | 0.957 |
| Perfectionism | 7698 | 0.574 | 5149 | 0.462 | 215 | 0.586 |
| Interpersonal distrust | 6987 | 0.098 | 4841 | 0.172 | 195 | 0.326 |
| Interoceptive Awareness | 7784 | 0.668 | 5447 | 0.896 | 157 | 0.071 |
| Maturity fears | 7664 | 0.539 | 5170 | 0.488 | 240 | 1.000 |
| Ascetism | 7816 | 0.704 | 5500 | 0.981 | 191 | 0.290 |
| Emotional Dysregulation | 7170 | 0.169 | 5120 | 0.425 | 176 | 0.167 |
| Social insecurity | 7430 | 0.331 | 4986 | 0.285 | 232 | 0.861 |
| Total | 7552 | 0.460 | 5200 | 0.560 | 211 | 0.529 |
| HADS Anxiety | 6356 | 0.104 | 4672 | 0.512 | 125 | 0.019 |
| HADS Depression | 7043 | 0.634 | 4890 | 0.865 | 192 | 0.418 |
| YBS Compulsion | 7126 | 0.697 | 4948 | 0.922 | 200 | 0.531 |
| YBS Obsession | 6751 | 0.310 | 4610 | 0.397 | 207 | 0.629 |
| BMI min | Current BMI | Illness Duration | Bradycardia | Physiological | Moderate | High Intensity | ||
|---|---|---|---|---|---|---|---|---|
| BMI min | Rho | — | ||||||
| p-value | — | |||||||
| Current BMI | Rho | 0.538 | — | |||||
| p-value | <0.001 | — | ||||||
| Illness duration | Rho | −0.131 | −0.040 | — | ||||
| p-value | 0.293 | 0.746 | — | |||||
| Bradycardia | Rho | 0.079 | −0.143 | −0.298 | — | |||
| p-value | 0.531 | 0.251 | 0.015 | — | ||||
| Physiological | Rho | 0.168 | 0.322 | 0.158 | −0.642 | — | ||
| p-value | 0.181 | 0.008 | 0.206 | <0.001 | — | |||
| Moderate | Rho | −0.234 | −0.032 | 0.277 | −0.733 | 0.343 | — | |
| p-value | 0.061 | 0.800 | 0.024 | <0.001 | 0.005 | — | ||
| High intensity | Rho | −0.271 | −0.084 | 0.213 | −0.557 | −0.032 | 0.669 | — |
| p-value | 0.029 | 0.502 | 0.087 | <0.001 | 0.797 | <0.001 | — |
| BMI min | Current BMI | Illness Duration | Brady-Cardia | Physiological | Moderate | High Intensity | ||
|---|---|---|---|---|---|---|---|---|
| BMI min | Rho | — | ||||||
| p-value | — | |||||||
| Current BMI | Rho | 0.620 | — | |||||
| p-value | <0.001 | — | ||||||
| Illness duration | Rho | −0.186 | −0.073 | — | ||||
| p-value | 0.182 | 0.600 | — | |||||
| Bradycardia | Rho | 0.121 | 0.081 | −0.423 | — | |||
| p-value | 0.394 | 0.566 | 0.002 | — | ||||
| Physiological | Rho | 0.050 | 0.173 | 0.195 | −0.670 | — | ||
| p-value | 0.725 | 0.217 | 0.161 | <0.001 | — | |||
| Moderate | Rho | −0.279 | −0.188 | 0.324 | −0.730 | 0.342 | — | |
| p-value | 0.045 | 0.177 | 0.018 | <0.001 | 0.013 | — | ||
| High intensity | Rho | −0.332 | −0.269 | 0.307 | −0.535 | −0.059 | 0.677 | — |
| p-value | 0.016 | 0.052 | 0.025 | <0.001 | 0.675 | <0.001 | — |
| BMI min | Current BMI | Illness Duration | Brady-cardia | Physiological | Moderate | High Intensity | ||
|---|---|---|---|---|---|---|---|---|
| BMI min | Rho | — | ||||||
| p-value | — | |||||||
| Current BMI | Rho | −0.234 | — | |||||
| p-value | 0.442 | — | ||||||
| Illness duration | Rho | 0.306 | −0.113 | — | ||||
| p-value | 0.310 | 0.714 | — | |||||
| Bradycardia | Rho | 0.479 | −0.126 | −0.415 | — | |||
| p-value | 0.098 | 0.683 | 0.158 | — | ||||
| Physiological | Rho | 0.193 | 0.297 | −0.116 | −0.462 | — | ||
| p-value | 0.528 | 0.325 | 0.707 | 0.115 | — | |||
| Moderate | Rho | −0.303 | −0.121 | 0.072 | −0.363 | 0.044 | — | |
| p-value | 0.315 | 0.696 | 0.816 | 0.224 | 0.892 | — | ||
| High intensity | Rho | −0.525 | −0.099 | −0.487 | −0.549 | −0.198 | 0.275 | — |
| p-value | 0.065 | 0.751 | 0.091 | 0.055 | 0.517 | 0.363 | — |
| Acyl-Ghrelin | Desacyl-Ghrelin | Leptin | |
|---|---|---|---|
| Short term | |||
| AL | 229+/−35.4 | 679+/−79.8 | 5724+/−866 |
| FR | 576+/−63.7 ** | 1583+/−160.5 * | 3548+/−248.9 * |
| FRW | 958+/−165.7 | 1763+/−252.7 | 3080+/−202.7 |
| Group effect (p) | <0.001 | <0.001 | 0.007 |
| Long term | |||
| AL | 166+/−42.5 | 1085+/−132.4 | 4668+/−419.8 |
| FR | 147+/−20.3 | 1459+/−108.4 | 3365+/−155.5 ** |
| FRW | 240+/−41.5 | 1534+/−132.2 | 3518+/−88.3 |
| Group effect (p) | 0.208 | 0.045 | 0.004 |
| Long term-Recovery | |||
| AL | 150+/−14.6 | 790+/−13.7 | 4594+/−380.7 |
| FR | 137+/−27.3 | 913+/−125.7 | 3639+/−95.4 * |
| FRW | 115+/−14 | 728+/−40.7 | 2915+/−100.9 |
| Group effect (p) | 0.633 | 0.433 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duriez, P.; Nilsson, I.A.K.; Le Thuc, O.; Alexandre, D.; Chartrel, N.; Rovere, C.; Chauveau, C.; Gorwood, P.; Tolle, V.; Viltart, O. Exploring the Mechanisms of Recovery in Anorexia Nervosa through a Translational Approach: From Original Ecological Measurements in Human to Brain Tissue Analyses in Mice. Nutrients 2021, 13, 2786. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082786
Duriez P, Nilsson IAK, Le Thuc O, Alexandre D, Chartrel N, Rovere C, Chauveau C, Gorwood P, Tolle V, Viltart O. Exploring the Mechanisms of Recovery in Anorexia Nervosa through a Translational Approach: From Original Ecological Measurements in Human to Brain Tissue Analyses in Mice. Nutrients. 2021; 13(8):2786. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082786
Chicago/Turabian StyleDuriez, Philibert, Ida A. K. Nilsson, Ophelia Le Thuc, David Alexandre, Nicolas Chartrel, Carole Rovere, Christophe Chauveau, Philip Gorwood, Virginie Tolle, and Odile Viltart. 2021. "Exploring the Mechanisms of Recovery in Anorexia Nervosa through a Translational Approach: From Original Ecological Measurements in Human to Brain Tissue Analyses in Mice" Nutrients 13, no. 8: 2786. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082786