
The Prophylactic Effects of Glutamine on Muscle Protein Synthesis and Degradation in Rats with Ethanol-Induced Liver Damage

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Protocol
2.3. Measurements and Analytical Procedures
2.3.1. Assessment of Liver Damage
Liver Function Index
Plasma Ammonia Concentration
Liver Histological Assessment
Hepatic Cytokine Levels
Protein Expression of Cytochrome P450 2E1 (CYP2E1)
Toll-Like Receptor 4 (TLR4) Signaling Pathway
2.3.2. Assessment of Small-Intestinal Damage
Ratio of Urine Lactulose (L) to Mannitol (M)
Plasma Endotoxin Concentration
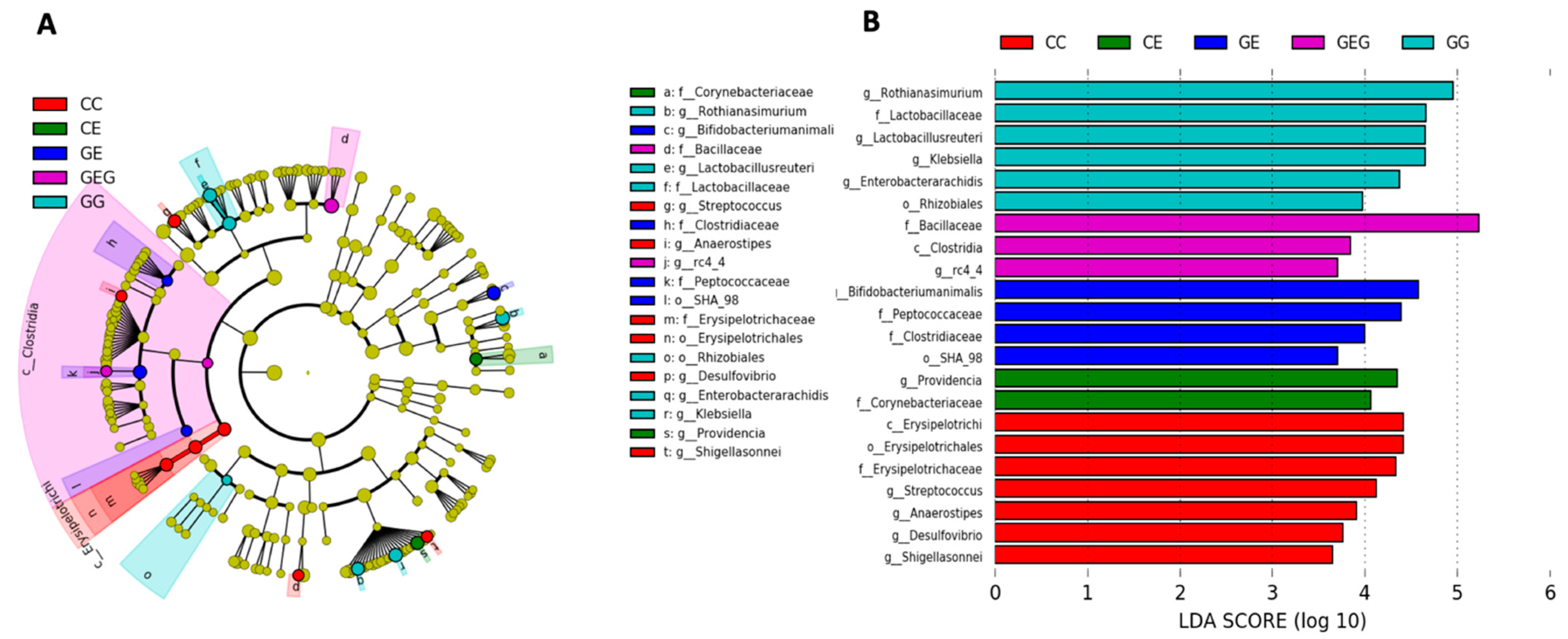
Intestinal Bacterial Composition
2.3.3. Assessment of Muscle Damage
Forelimb Grip Strength Test
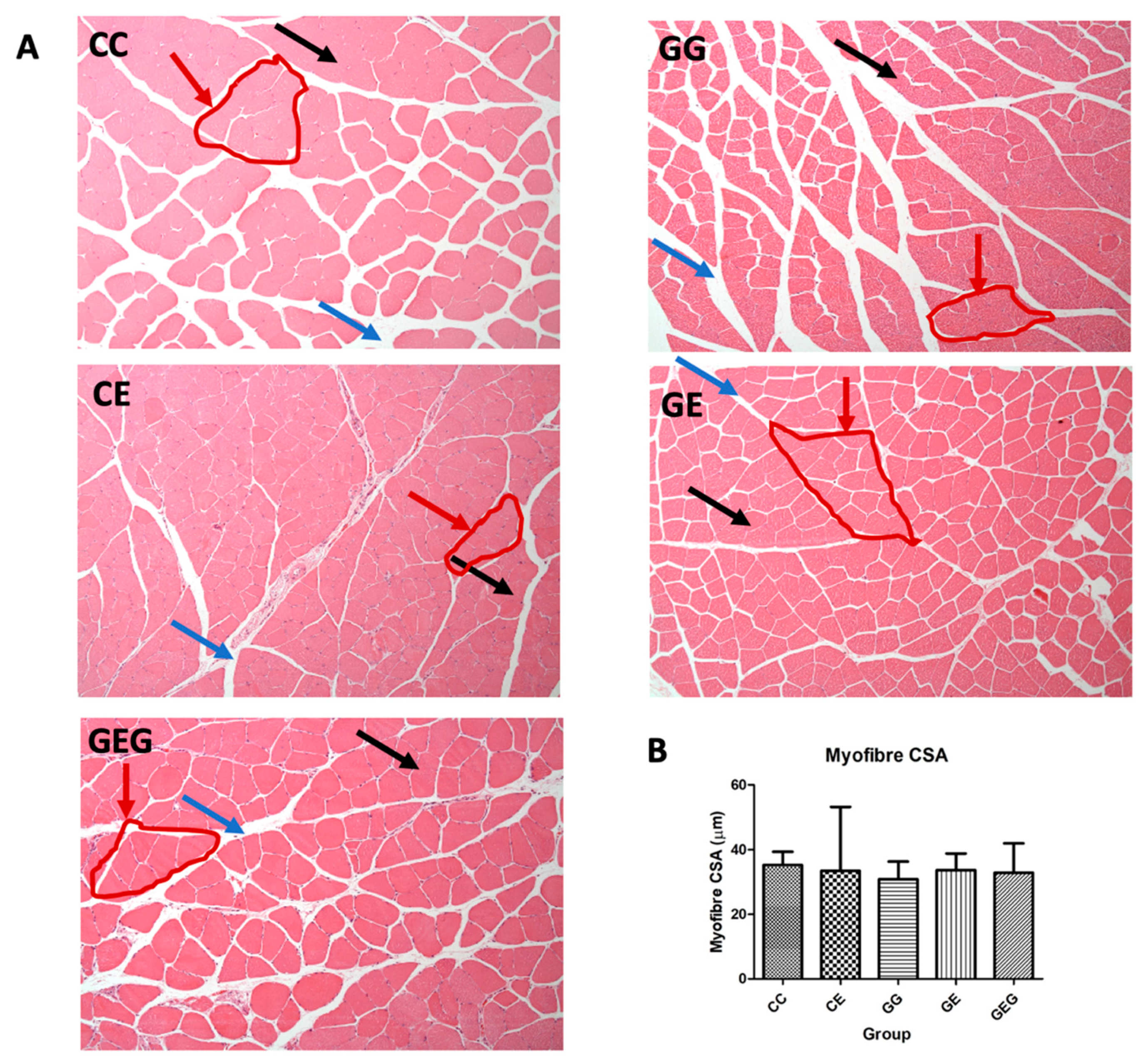
Muscle Histological Assessment
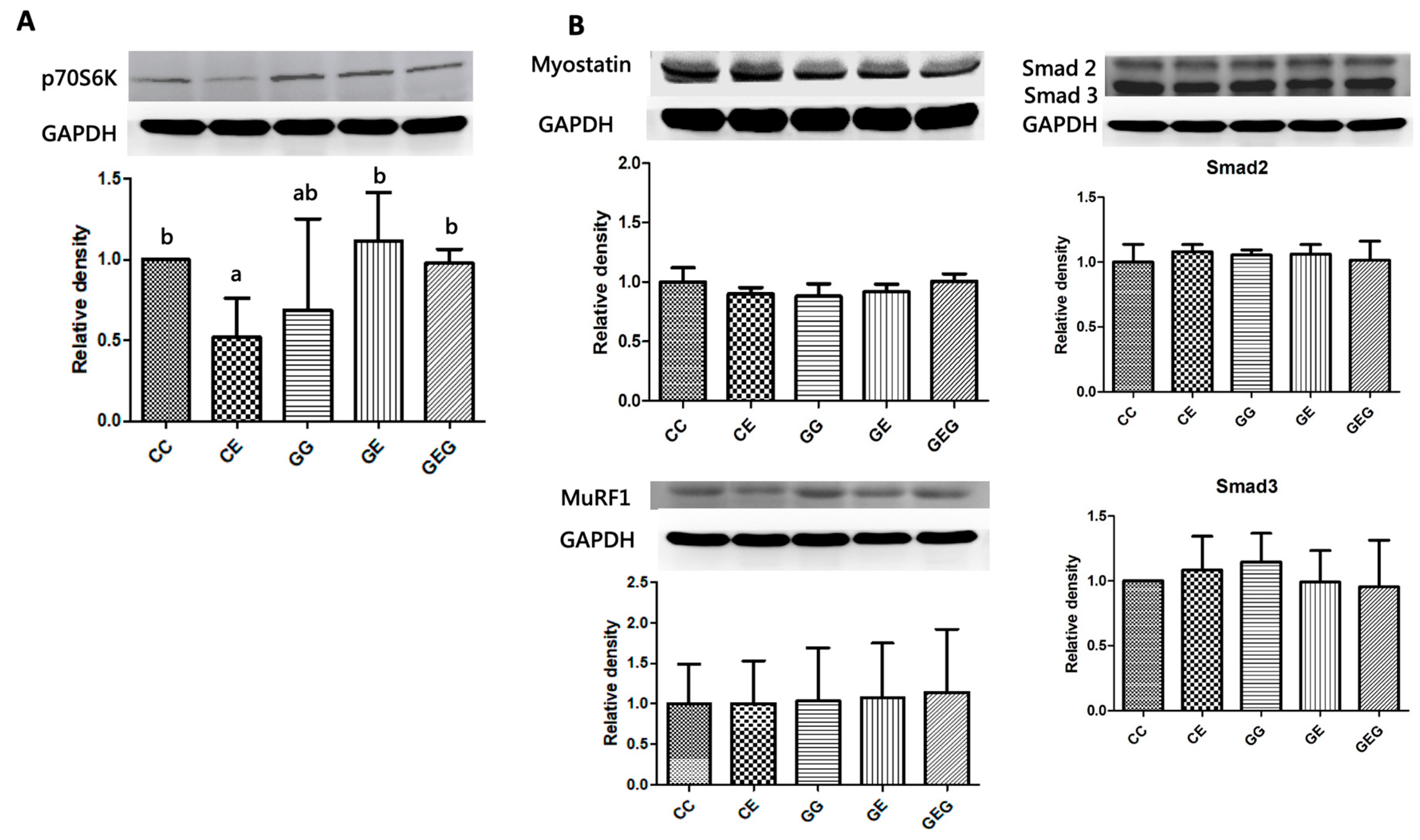
Muscle Protein Synthesis Markers
Muscle Protein Degradation Markers
2.3.4. Amino Acid Composition
2.3.5. Statistical Analysis
3. Results
3.1. Food Intake, Ethanol Consumption, Final BW, Relative Liver Weight, and Muscle Weights
3.2. Assessment of Liver Damage
3.2.1. Plasma AST and ALT Activities and Ammonia Level
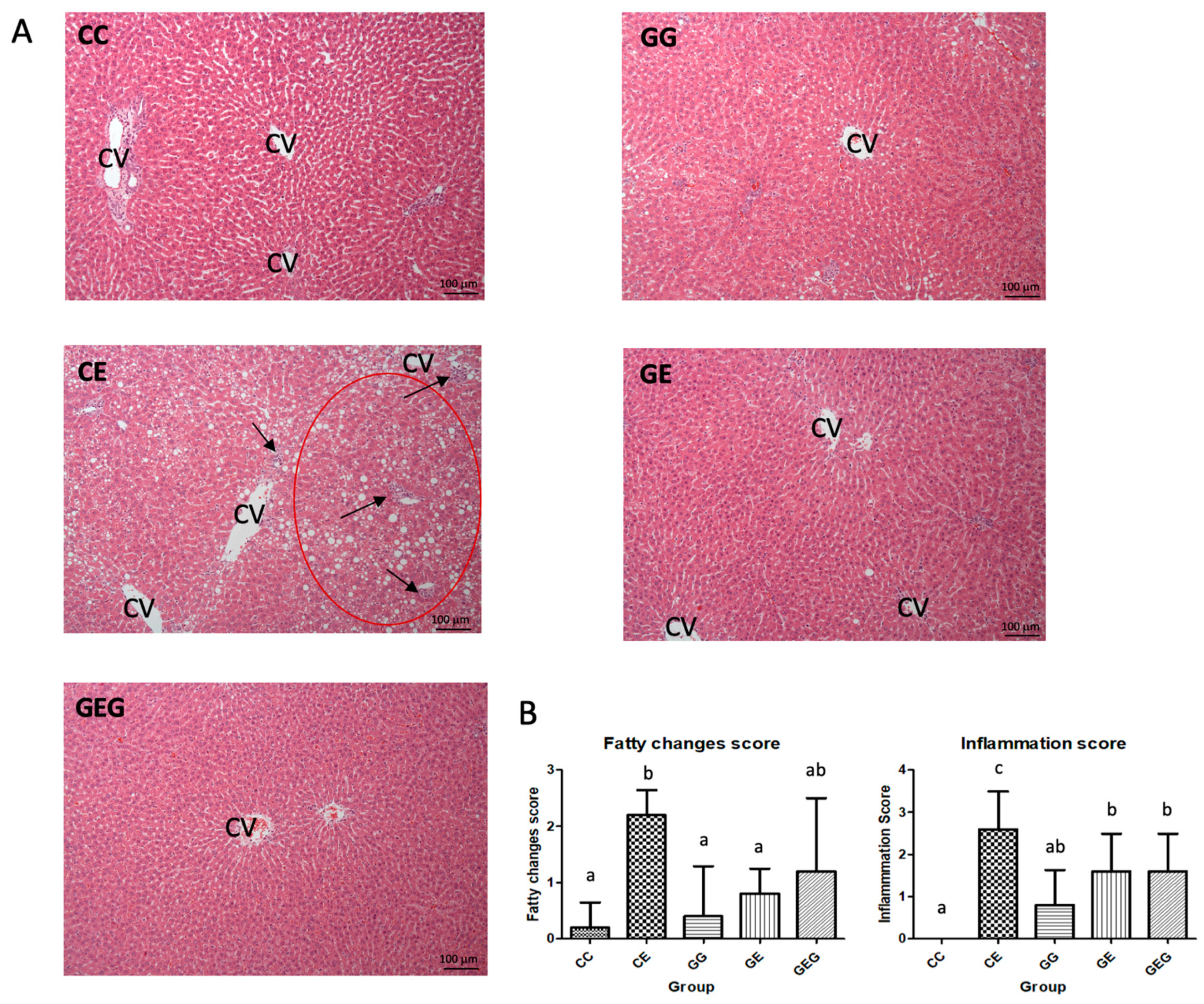
3.2.2. Hepatic Histopathology Scores and Cytokine Levels
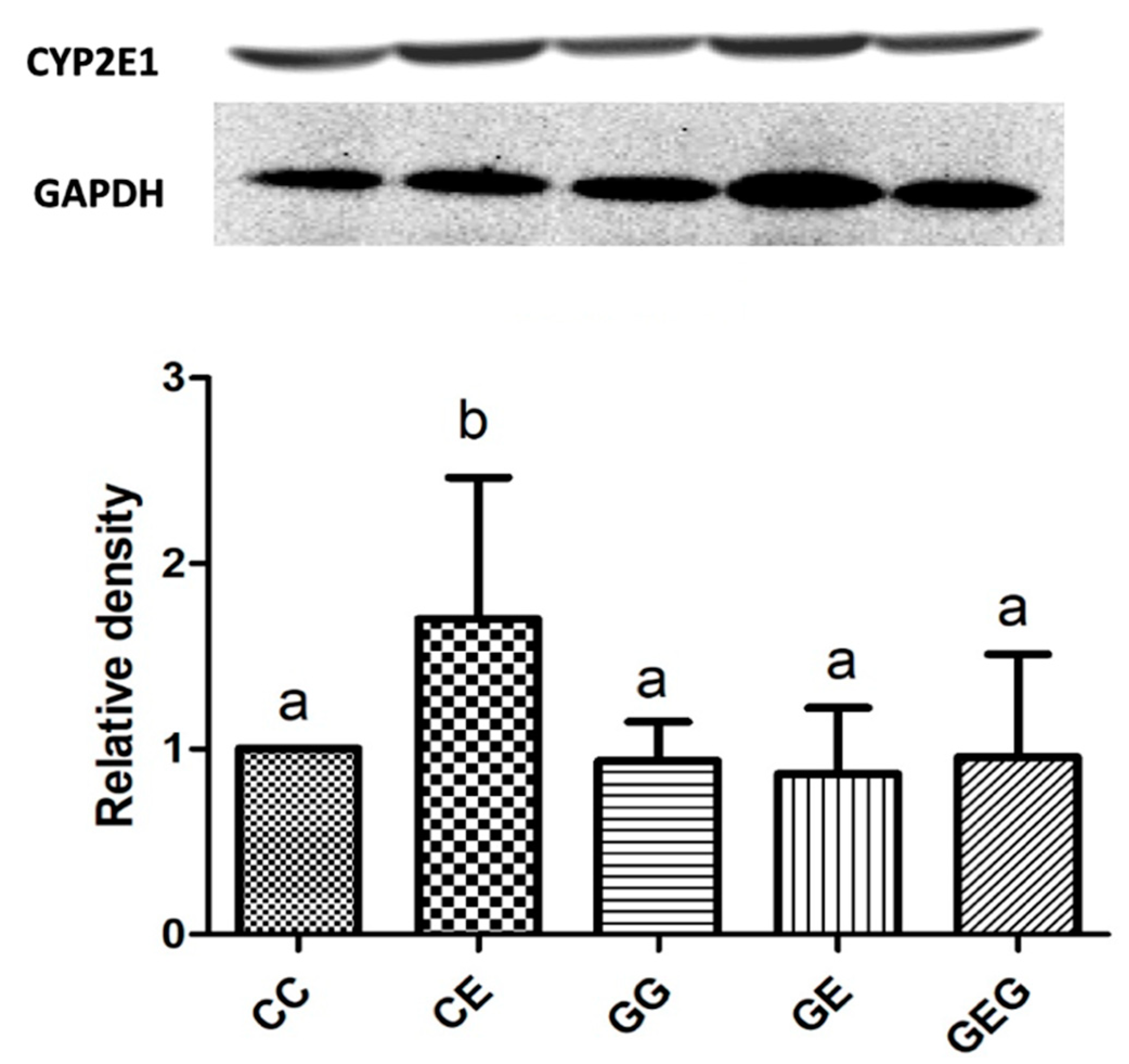
3.2.3. Hepatic CYP2E1
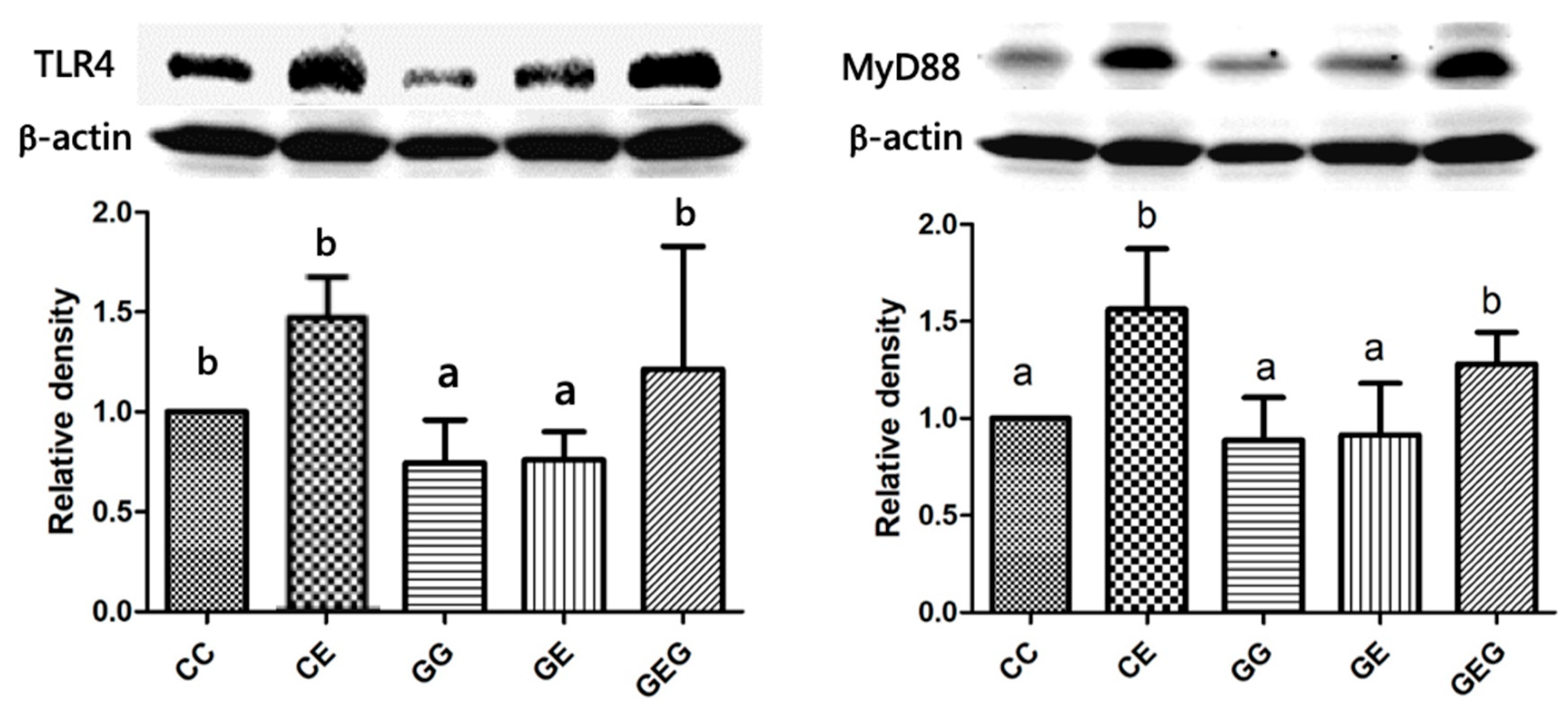
3.2.4. TLR4-Signaling Pathway
3.3. Assessment of Gut Damage
3.3.1. Intestinal Permeability
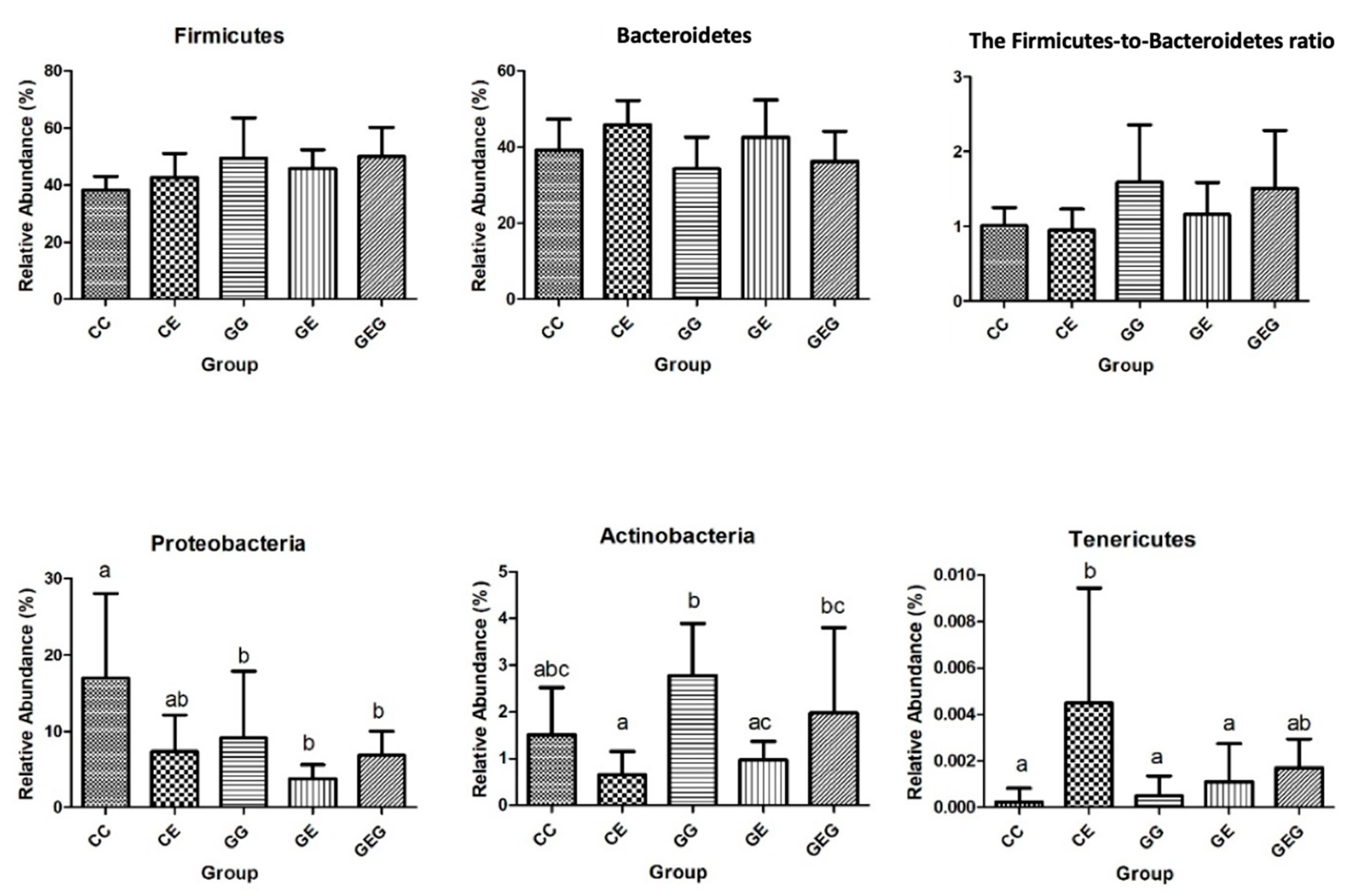
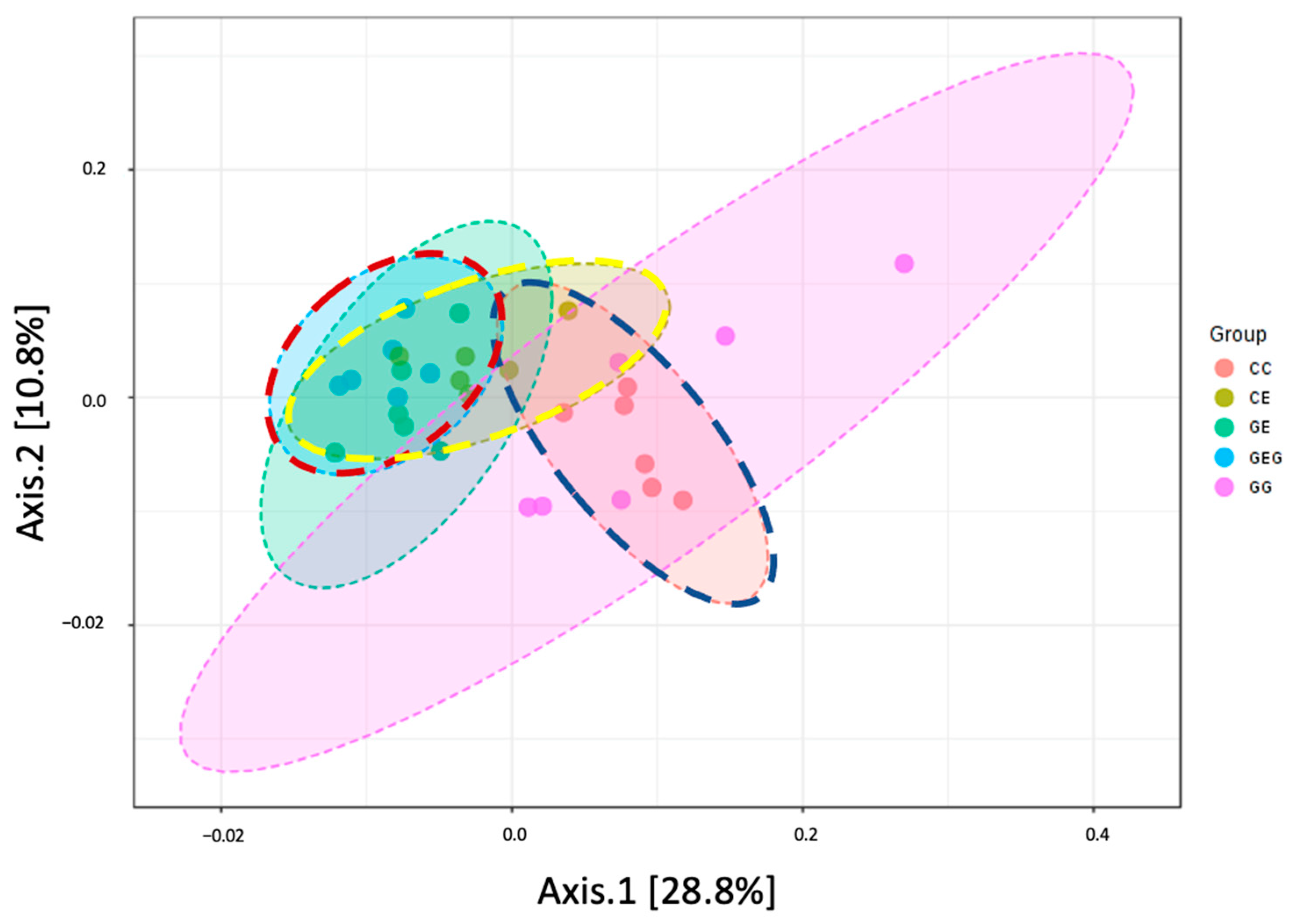
3.3.2. Fecal Bacterial Composition
3.4. Assessment of Muscle Damage
3.4.1. Forelimb Grip Strength Test
3.4.2. Muscle Histological Assessment
3.4.3. Muscle Protein Synthesis Markers
3.4.4. Muscle Protein Degradation Markers
3.5. Amino Acid Composition
4. Discussion
4.1. BW and Liver Weight
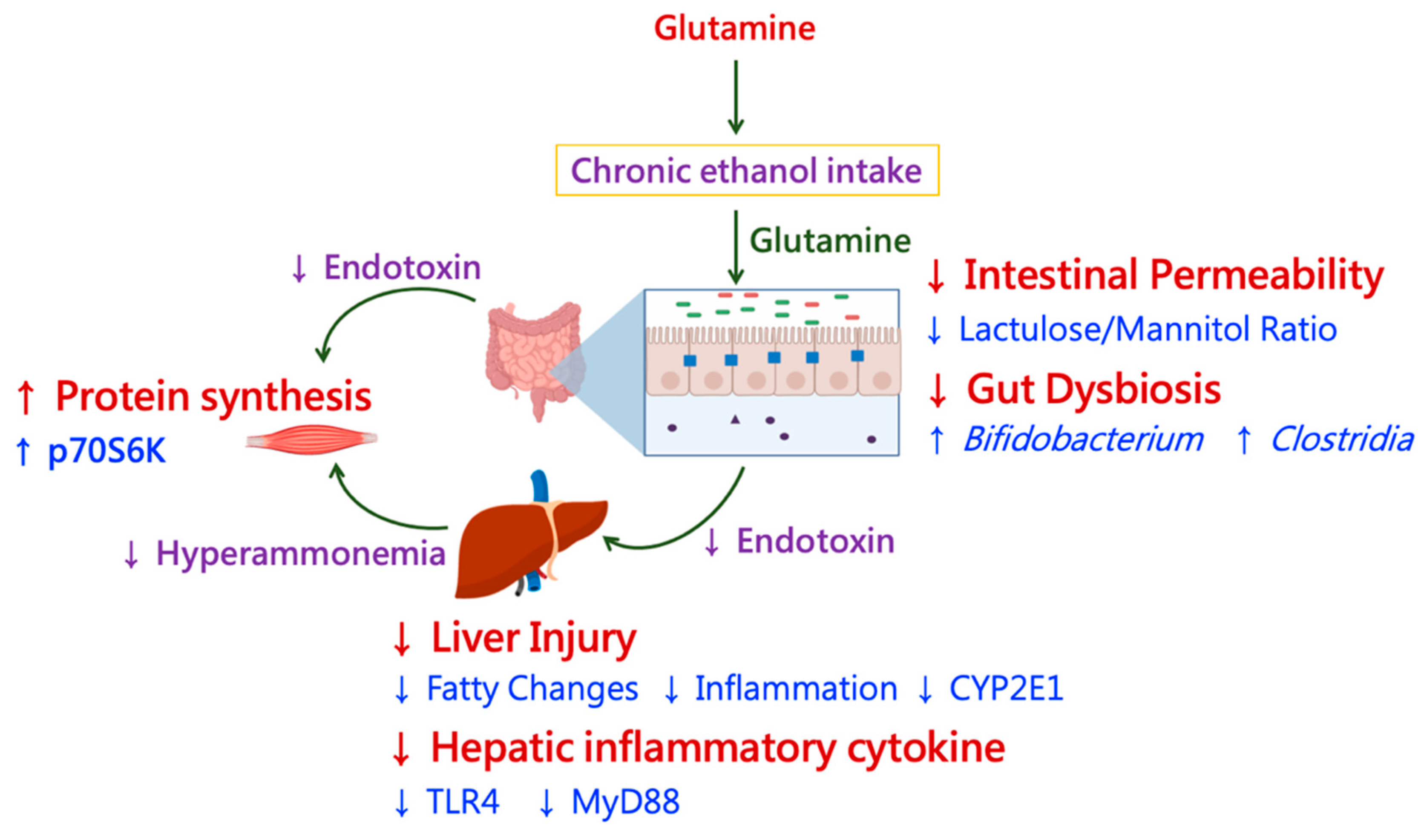
4.2. Ethanol, Glutamine, and Liver Damage
4.3. Ethanol, Glutamine, and Gut Damage
4.4. Ethanol, Glutamine, and Muscular Protein Synthesis and Degradation
4.5. Glutamine and Gut–Muscle Axis with Chronic Ethanol Intake
4.6. Glutamine and the Liver–Muscle Axis with Chronic Ethanol Intake
4.7. The Study Limitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Rehm, J.; Samokhvalov, A.V.; Shield, K.D. Global burden of alcoholic liver diseases. J. Hepatol. 2013, 59, 160–168. [Google Scholar] [CrossRef] [Green Version]
- 2019 Cause of Death Statistics in Taiwan; Ministry of Health and Welfare: Taipei, Taiwan, 2020.
- Dasarathy, S. Myostatin and beyond in cirrhosis: All roads lead to sarcopenia. J. Cachex-Sarcopenia Muscle 2017, 8, 864–869. [Google Scholar] [CrossRef]
- Dasarathy, S. Nutrition and alcoholic liver disease: Effects of alcoholism o0n nutrition, effects of nutrition on alcoholic liver disease, and nutritional therapies for alcoholic liver disease. Clin. Liver Dis. 2016, 20, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Dasarathy, S. Consilience in sarcopenia of cirrhosis. J. Cachex-Sarcopenia Muscle 2012, 3, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Thapaliya, S.; Runkana, A.; McMullen, M.R.; Nagy, L.E.; McDonald, C.; Prasad, S.V.N.; Dasarathy, S. Alcohol-induced autophagy con-tributes to loss in skeletal muscle mass. Autophagy 2014, 10, 677–690. [Google Scholar] [CrossRef]
- Szabo, G.; Petrasek, J. Gut–liver axis and sterile signals in the development of alcoholic liver disease. Alcohol Alcohol. 2017, 52, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Tirapegui, J.; Cruzat, V.F. Glutamine and Skeletal Muscle. In Glutamine in Clinical Nutrition; Rajendram, R., Preedy, V.R., Patel, V.B., Eds.; Springer: New York, NY, USA, 2015; pp. 499–511. [Google Scholar]
- Kuhn, K.S.; Schuhmann, K.; Stehle, P.; Darmaun, M.; Fürst, P. Determination of glutamine in muscle protein facilitates accurate assessment of proteolysis and de novo synthesis-derived endogenous glutamine production. Am. J. Clin. Nutr. 1999, 70, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.W.; Goldberg, A.L. The metabolic fates of amino acids and theformation of glutamine in skeletal muscle. J. Biol. Chem. 1978, 253, 3685–3693. [Google Scholar] [CrossRef]
- Wang, X.; Pierre, J.F.; Heneghan, A.F.; Busch, R.A.; Kudsk, K.A. Glutamine improves innate immunity and prevents bacterial en-teroinvasion during parenteral nutrition. JPEN J. Parenter Enteral Nutr. 2015, 39, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Wu, Z.; Ji, Y.; Sun, K.; Dai, Z.; Wu, G. L-Glutamine Enhances Tight Junction Integrity by Activating CaMK Kinase 2–AMP-Activated Protein Kinase Signaling in Intestinal Porcine Epithelial Cells. J. Nutr. 2016, 146, 501–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermon, S.; Castell, L.M.; Calder, P.; Bishop, N.C.; Blomstrand, E.; Mooren, F.C.; Krüger, K.; Kavazis, A.N.; Quindry, J.C.; Senchina, D.; et al. Consensus Statement Immunonutrition and Exercise. Exerc. Immunol. Rev. 2017, 23, 8–50. [Google Scholar] [PubMed]
- Lacey, J.M.; Wilmore, D.W. Is Glutamine a Conditionally Essential Amino Acid? Nutr. Rev. 2009, 48, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Castell, L.M. Glutamine Supplementation In Vitro and In Vivo, in Exercise and in Immunodepression. Sports Med. 2003, 33, 323–345. [Google Scholar] [CrossRef] [PubMed]
- Fläring, U.; Rooyackers, O.; Wernerman, J.; Hammarqvist, F. Glutamine attenuates post-traumatic glutathione depletion in human muscle. Clin. Sci. 2003, 104, 275–282. [Google Scholar] [CrossRef]
- Rohde, T.; A MacLean, D.; Hartkopp, A.; Pedersen, B.K. The immune system and serum glutamine during a triathlon. Eur. J. Appl. Physiol. Occup. Physiol. 1996, 74, 428–434. [Google Scholar] [CrossRef]
- Parry-Billings, M.; Calder, P.; Newsholme, E.; Evans, J. Does glutamine contribute to immunosuppression after major burns? Lancet 1990, 336, 523–525. [Google Scholar] [CrossRef]
- Lambertucci, A.C.; Lambertucci, R.H.; Hirabara, S.M.; Curi, R.; Moriscot, A.S.; Alba-Loureiro, T.C.; Guimarães-Ferreira, L.; Levada-Pires, A.C.; Vasconcelos, D.A.; Sellitti, D.F.; et al. Glutamine supplementation stimulates protein-synthetic and inhibits proteindegradative signaling pathways in skeletal muscle of diabetic rats. PLoS ONE 2012, 7, e50390. [Google Scholar] [CrossRef] [Green Version]
- Brunt, E.M.; Janney, C.G.; Di Bisceglie, A.M.; Neuschwander-Tetri, B.A.; Bacon, B.R. Nonalcoholic steatohepatitis: A proposal for grading and staging the histological lesions. Am. J. Gastroenterol. 1999, 94, 2467–2474. [Google Scholar] [CrossRef]
- Lu, N.S.; Chiu, W.C.; Chen, Y.L.; Peng, H.C.; Shirakawa, H.; Yang, S.C. Fish oil up-regulates hepatic autophagy in rats with chronic ethanol consumption. J. Nutr. Biochem. 2020, 77, 108314. [Google Scholar] [CrossRef]
- Uesugi, T.; Froh, M.; Arteel, G.E.; Bradford, B.U.; Wheeler, M.D.; Gäbele, E.; Isayama, F.; Thurman, R.G. Role of Lipopolysaccha-ride-Binding Protein in Early Alcohol-Induced Liver Injury in Mice. J. Immunol. 2002, 168, 2963–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-L.; Peng, H.-C.; Tan, S.-W.; Tsai, C.-Y.; Huang, Y.-H.; Wu, H.-Y.; Yang, S.-C. Amelioration of ethanol-induced liver injury in rats by nanogold flakes. Alcohol 2013, 47, 467–472. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Choudhary, S.; Holmes, E.W.; Yong, S.; Banan, A.; Jakate, S.; Fields, J.Z. Preventing gut leakiness by oats supple-mentation ameliorates alcohol-induced liver damage in rats. J. Pharmacol. Exp. Ther. 2001, 299, 442–448. [Google Scholar]
- Glass, D.J. Skeletal muscle hypertrophy and atrophy signaling pathways. Int. J. Biochem. Cell Biol. 2005, 37, 1974–1984. [Google Scholar] [CrossRef]
- Yuan, M.; Breitkopf, S.B.; Yang, X.; Asara, J.M. A positive/negative ion–switching, targeted mass spectrometry–based metabo-lomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872. [Google Scholar] [CrossRef] [Green Version]
- Mezey, E. Interaction Between Alcohol and Nutrition in the Pathogenesis of Alcoholic Liver Disease. Semin. Liver Dis. 1991, 11, 340–348. [Google Scholar] [CrossRef]
- Israel, Y.; Britton, R.S.; Orrego, H. Liver cell enlargement induced by chronic alcohol consumption: Studies on its causes and consequences. Clin. Biochem. 1982, 15, 189–192. [Google Scholar] [CrossRef]
- Peng, H.C.; Chen, Y.L.; Chen, J.R.; Yang, S.S.; Huang, K.H.; Wu, Y.C.; Lin, Y.H.; Yang, S.C. Effects of glutamine administration on in-flammatory responses in chronic ethanol-fed rats. J. Nutr. Biochem. 2011, 22, 282–288. [Google Scholar] [CrossRef]
- Chiu, W.C.; Huang, Y.L.; Chen, Y.L.; Peng, H.C.; Liao, W.H.; Chuang, H.L.; Chen, J.R.; Yang, S.C. Synbiotics reduce ethanol-induced he-patic steatosis and inflammation by improving intestinal permeability and microbiota in rats. Food Funct. 2015, 6, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, S.; Lim, A.; Catalano, D.; Mandrekar, P. Human Binge Alcohol Intake Inhibits TLR4–MyD88 and TLR4–TRIF Re-sponses but Not the TLR3-TRIF Pathway: HspA1A and PP1 Play Selective Regulatory Roles. J. Immunol. 2018, 200, 2291–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, Y.S.; Zhang, B.; Loomba, R.; Seki, E. TLR2 and TLR9 contribute to alcohol-mediated liver injury through induction of CXCL1 and neutrophil infiltration. Am. J. Physiol. Liver Physiol. 2015, 309, G30–G41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.-L.; Li, X.-L.; Xi, P.-B.; Zhang, J.; Wu, G.; Zhu, W.-Y. l-Glutamine regulates amino acid utilization by intestinal bacteria. Amino Acids 2012, 45, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Peng, H.C.; Chien, Y.W.; Chen, Y.L.; Lu, N.S.; Yang, S.C. Effects of fish oil on lipid metabolism and its molecular biolog-ical regulators in chronic ethanol-fed rats. Nutrients 2018, 10, 2018. [Google Scholar] [CrossRef] [Green Version]
- Bull-Otterson, L.; Feng, W.; Kirpich, I.; Wang, Y.; Qin, X.; Liu, Y.; Gobejishvili, L.; Joshi-Barve, S.; Ayvaz, T.; Petrosino, J.; et al. Metagenomic analyses of alcohol induced pathogenic alterations in the intestinal microbi-ome and the effect of Lactobacillus rhamnosus GG treatment. PLoS ONE 2013, 8, e53028. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Farhadi, A.; Jakate, S.M.; Tang, Y.; Shaikh, M.; Keshavarzian, A. Lactobacillus GG treatment ameliorates alco-hol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol 2009, 43, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Shao, T.; Zhao, C.; Li, F.; Gu, Z.; Liu, L.; Zhang, L.; Wang, Y.; He, L.; Liu, Y.; Liu, Q. Intestinal HIF-1α deletion exacerbates alcoholic liv-er disease by inducing intestinal dysbiosis and barrier dysfunction. J. Hepatol. 2018, 69, 886–895. [Google Scholar] [CrossRef]
- Wang, Y.; Kirpich, I.; Liu, Y.; Ma, Z.; Barve, S.; McClain, C.J.; Feng, W. Lactobacillus rhamnosus GG Treatment Potentiates Intestinal Hypoxia-Inducible Factor, Promotes Intestinal Integrity and Ameliorates Alcohol-Induced Liver Injury. Am. J. Pathol. 2011, 179, 2866–2875. [Google Scholar] [CrossRef] [PubMed]
- Bhagwandeen, B.; Apte, M.; Manwarring, L.; Dickeson, J. Endotoxin induced hepatic necrosis in rats on an alcohol diet. J. Pathol. 1987, 152, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, L.; Crabb, D.W. Alcoholic liver disease. Curr. Opin. Gastroenterol. 2001, 17, 211–220. [Google Scholar] [CrossRef]
- Bjorkholm, B.; Olling, S.; Larsson, P.; Hagberg, L. An outbreak of diphtheria among Swedish alcoholics. Infection 1987, 1, 354–358. [Google Scholar] [CrossRef]
- Hartmann, P.; Seebauer, C.T.; Schnabl, B. Alcoholic Liver Disease: The Gut Microbiome and Liver Cross Talk. Alcohol. Clin. Exp. Res. 2015, 39, 763–775. [Google Scholar] [CrossRef] [Green Version]
- Acharya, C.; Bajaj, J.S. Gut Microbiota and Complications of Liver Disease. Gastroenterol. Clin. N. Am. 2017, 46, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Basuroy, S.; Sheth, P.; Mansbach, C.M.; Rao, R.K. Acetaldehyde disrupts tight junctions and adherens junctions in human colonic mucosa: Protection by EGF and l-glutamine. Am. J. Physiol. Liver Physiol. 2005, 289, G367–G375. [Google Scholar] [CrossRef]
- Seth, A.; Basuroy, S.; Sheth, P.; Rao, R.K. l-Glutamine ameliorates acetaldehyde-induced increase in paracellular permeability in Caco-2 cell monolayer. Am. J. Physiol. Liver Physiol. 2004, 287, G510–G517. [Google Scholar] [CrossRef]
- Sukhotnik, I.; Agam, M.; Shamir, R.; Shehadeh, N.; Lurie, M.; Coran, A.G.; Shiloni, E.; Mogilner, J. Oral glutamine prevents gut mu-cosal injury and improves mucosal recovery following lipopolysaccharide endotoxemia in a rat. J. Surg. Res. 2007, 143, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, K.K.; Shukla, P.K.; Mir, H.; Manda, B.; Gangwar, R.; Yadav, N.; McMullen, M.; Nagy, L.E.; Rao, R. Glutamine supplementa-tion attenuates ethanol-induced disruption of apical junctional complexes in colonic epithelium and ameliorates gut barrier dysfunction and fatty liver in mice. J. Nutr. Biochem. 2016, 27, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Lin, Z.; Zeng, Y.; Lin, X.; Zhang, Y. Probiotic and glutamine treatments attenuate alcoholic liver disease in a rat mod-el. Exp. Ther. Med. 2019, 18, 4733–4739. [Google Scholar] [PubMed]
- Sarin, S.K.; Pande, A.; Schnabl, B. Microbiome as a therapeutic target in alcohol-related liver disease. J. Hepatol. 2019, 70, 260–272. [Google Scholar] [CrossRef] [Green Version]
- Moens, F.; Verce, M.; De Vuyst, L. Lactate- and acetate-based cross-feeding interactions between selected strains of lactobacilli, bifidobacteria and colon bacteria in the presence of inulin-type fructans. Int. J. Food Microbiol. 2017, 241, 225–236. [Google Scholar] [CrossRef]
- Ramakrishna, B. Role of the gut microbiota in human nutrition and metabolism. J. Gastroenterol. Hepatol. 2013, 28, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Shenkman, B.S.; Belova, S.P.; Zinovyeva, O.; Samkhaeva, N.D.; Mirzoev, T.; Vilchinskaya, N.A.; Altaeva, E.G.; Turtikova, O.V.; Kostrominova, T.Y.; Nemirovskaya, T.L. Effect of Chronic Alcohol Abuse on Anabolic and Catabolic Signaling Pathways in Human Skeletal Muscle. Alcohol. Clin. Exp. Res. 2018, 42, 41–52. [Google Scholar] [CrossRef]
- Shenkman, B.S.; Zinovyeva, O.; Belova, S.P.; Mirzoev, T.M.; Vilchinskaya, N.A.; Vikhlyantsev, I.M.; Ulanova, A.D.; Turtikova, O.V.; Samkhaeva, N.D.; Parfenov, V.; et al. Cellular and molecular signatures of alcohol-induced myopathy in women. Am. J. Physiol. Metab. 2019, 316, E967–E976. [Google Scholar] [CrossRef]
- Steiner, J.L.; Gordon, B.S.; Lang, C.H. Moderate alcohol consumption does not impair overload-induced muscle hypertrophy and protein synthesis. Physiol. Rep. 2015, 3, e12333. [Google Scholar] [CrossRef] [Green Version]
- Steiner, J.L.; Lang, C.H. Alcohol intoxication following muscle contraction in mice decreases muscle protein synthesis but not mTOR signal transduction. Alcohol. Clin. Exp. Res. 2015, 39, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korzick, D.H.; Sharda, D.R.; Pruznak, A.M.; Lang, C.H. Aging accentuates alcohol-induced decrease in protein synthesis in gas-trocnemius. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R887–R898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, J.L.; Lang, C.H. Dysregulation of skeletal muscle protein metabolism by alcohol. Am. J. Physiol. Metab. 2015, 308, E699–E712. [Google Scholar] [CrossRef] [Green Version]
- Gumucio, J.P.; Sugg, K.B.; Mendias, C.L. TGF-β Superfamily Signaling in Muscle and Tendon Adaptation to Resistance Exercise. Exerc. Sport Sci. Rev. 2015, 43, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stitt, T.N.; Drujan, D.; Clarke, B.A.; Panaro, F.; Timofeyva, Y.; Kline, W.O.; Gonzalez, M.; Yancopoulos, G.D.; Glass, D.J. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcrip-tion factors. Mol. Cell 2004, 14, 395–403. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glass, D.J. Signalling pathways that mediate skeletal muscle hypertrophy and atrophy. Nat. Cell Biol. 2003, 5, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Nathan, J.A.; Goldberg, A.L. Muscle wasting in disease: Molecular mechanisms and promising therapies. Nat. Rev. Drug Discov. 2015, 14, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Haba, Y.; Fujimura, T.; Oyama, K.; Kinoshita, J.; Miyashita, T.; Fushida, S.; Harada, S.; Ohta, T. Effect of Oral Branched-Chain Amino Acids and Glutamine Supplementation on Skeletal Muscle Atrophy After Total Gastrectomy in Rat Model. J. Surg. Res. 2019, 243, 281–288. [Google Scholar] [CrossRef]
- Morales, M.G.; Olguín, H.; Di Capua, G.; Brandan, E.; Simon, F.; Cabello-Verrugio, C. Endotoxin-induced skeletal muscle wasting is prevented by angiotensin-(1–7) through a p38 MAPK-dependent mechanism. Clin. Sci. 2015, 129, 461–476. [Google Scholar] [CrossRef]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabo-lism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef]
- Dasarathy, S.; Hatzoglou, M. Hyperammonemia and proteostasis in cirrhosis. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Holecek, M. Evidence of a vicious cycle in glutamine synthesis and breakdown in pathogenesis of hepatic encephalopa-thy-therapeutic perspectives. Metab. Brain Dis. 2014, 29, 9–17. [Google Scholar] [CrossRef] [Green Version]
- HoleČek, M. Branched-chain amino acids in health and disease: Metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. 2018, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holeček, M. Branched-chain amino acid supplementation in treatment of liver cirrhosis: Updated views on how to attenuate their harmful effects on cataplerosis and ammonia formation. Nutrition 2017, 41, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.-L.; Lai, Y.-N.; Shang, H.-F.; Lin, M.-T.; Chen, W.-J. Effects of glutamine supplementation on innate immune response in rats with gut-derived sepsis. Br. J. Nutr. 2004, 91, 423–429. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient 2 | Diet Type | |||
|---|---|---|---|---|
| Control | Glutamine 3 | Ethanol | Ethanol + Glutamine 3 | |
| (g/L (1000 kcal)) | ||||
| Casein | 41.4 | 31.03 | 41.4 | 31.03 |
| Maltodextrin | 115.2 | 117.1 | 25.6 | 27.5 |
| ICN: AIN-76 vitamins | 2.5 | 2.5 | 2.5 | 2.5 |
| ICN: AIN-76 minerals | 2.6 | 2.6 | 2.6 | 2.6 |
| L-Cysteine | 0.5 | 0.5 | 0.5 | 0.5 |
| DL-Methionine | 0.3 | 0.3 | 0.3 | 0.3 |
| Olive oil | 28.4 | 28.4 | 28.4 | 28.4 |
| Corn oil | 8.5 | 8.5 | 8.5 | 8.5 |
| Safflower oil | 2.7 | 2.7 | 2.7 | 2.7 |
| Choline bitartrate | 0.53 | 0.53 | 0.53 | 0.53 |
| Fiber | 10 | 10 | 10 | 10 |
| Xanthan gum | 3 | 3 | 3 | 3 |
| Ethanol | - | - | 50 | 50 |
| Glutamine | - | 8.437 | - | 8.437 |
| Total nitrogen | 6.47 | 6.47 | 6.47 | 6.47 |
| Group | Final Body Weight (g) | Relative Liver Weight 3 (%) | Quadriceps Weight (g) | Gastrocnemius Weight (g) |
|---|---|---|---|---|
| CC | 422.9 ± 7.5 a | 2.4 ± 0.2 a | 2.5 ± 0.4 | 3.5 ± 0.4 |
| CE | 389.1 ± 34.9 bc | 2.9 ± 0.2 b | 2.7 ± 0.5 | 3.2 ± 0.5 |
| GG | 407.1 ± 16.1 ab | 2.3 ± 0.2 a | 2.4 ± 0.6 | 3.3 ± 0.6 |
| GE | 375.8 ± 23.7 c | 2.8 ± 0.2 b | 2.2 ± 0.4 | 3.3 ± 0.4 |
| GEG | 381.3 ± 19.2 c | 2.7 ± 0.2 b | 2.3 ± 0.5 | 3.0 ± 0.3 |
| Group | AST (U/L) | ALT (U/L) | Ammonia (μg/dL) |
|---|---|---|---|
| CC | 73.9 ± 7.4 a | 43.3 ± 6.8 a | 175.1 ± 22.1 a |
| CE | 130.4 ± 33.5 c | 113.4 ± 65.5 b | 221.7 ± 85.0 b |
| GG | 74.3 ± 4.8 a | 39.6 ± 7.1 a | 152.1 ± 22.0 ac |
| GE | 94.8 ± 22.5 b | 65.6 ± 19.3 a | 99.4 ± 33.3 cd |
| GEG | 100.6 ± 12.6 b | 54.1 ± 14.7 a | 70.3 ± 19.8 d |
| Groups | IL-1β (pg/mg Protein) | IL-6 (pg/mg Protein) | IL-10 (pg/mg Protein) | TNF-α (pg/mg Protein) |
|---|---|---|---|---|
| CC | 29.0 ± 4.9 a | 111.6 ± 26.0 a | 48.3 ± 6.6 a | 19.7 ± 2.0 a |
| CE | 53.1 ± 3.6 b | 149.3 ± 18.1 b | 72.6 ± 15.7 b | 32.8 ± 4.2 b |
| GG | 28.7 ± 3.1 a | 114.6 ± 24.7 a | 49.6 ± 13.2 a | 20.8 ± 3.6 a |
| GE | 28.9 ± 6.1 a | 106.7 ± 33.2 a | 50.8 ± 15.9 a | 16.9 ± 4.6 a |
| GEG | 28.2 ± 6.5 a | 104.2 ± 31.9 c | 60.4 ± 12.0 ac | 18.4 ± 3.8 a |
| Group | Ratio of Lactulose to Mannitol | Endotoxin (EU/mL) |
|---|---|---|
| CC | 0.9 ± 0.0 a | 22.7 ± 3.3 a |
| CE | 3.0 ± 0.0 c | 29.7 ± 5.3 b |
| GG | 2.1 ± 0.0 b | 24.1 ± 4.5 a |
| GE | 2.6 ± 0.0 d | 23.4 ± 4.2 a |
| GEG | 1.0 ± 0.0 e | 20.7 ± 3.3 a |
| Group | Shannon | Simpson | ACE | Chao1 |
|---|---|---|---|---|
| CC | 3.1 ± 0.2 | 0.9 ± 0.0 | 518.4 ± 24.9 | 520.3 ± 21.7 |
| CE | 3.2 ± 0.3 | 0.8 ± 0.0 | 545.2 ± 17.7 | 553.7 ± 23.7 |
| GG | 3.3 ± 0.3 | 0.9 ± 0.0 | 514.2 ± 61.4 | 529.9 ± 67.7 |
| GE | 3.3 ± 0.5 | 0.8 ± 0.1 | 565.5 ± 14.2 | 568.4 ± 15.5 |
| GEG | 3.5 ± 0.2 | 0.9 ± 0.0 | 553.0 ± 24.8 | 561.9 ± 21.6 |
| Group | Grip Strength (g) | Difference in Grip Strength (g) | Difference in Grip Strength (g/kg BW) | |
|---|---|---|---|---|
| Initial | Final | |||
| CC | 1287.7 ± 206.8 | 1799.6 ± 254.8 | 511.9 ± 314.0 | 1217.9 ± 756.4 |
| CE | 1237.8 ± 135.2 | 1637.2 ± 282.1 | 399.4 ± 369.0 | 1021.0 ± 909.6 |
| GG | 1439.3 ± 137.2 | 1891.0 ± 127.0 | 451.7 ± 188.3 | 1115.1 ± 473.2 |
| GE | 1283.1 ± 146.6 | 1778.0 ± 240.2 | 494.9 ± 252.7 | 1306.3 ± 666.6 |
| GEG | 1438.7 ± 146.0 | 1766.0 ± 231.1 | 327.3 ± 339.3 | 867.0 ± 897.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Q.; Chen, Y.-H.; Pratama, S.A.; Chen, Y.-L.; Shirakawa, H.; Peng, H.-C.; Yang, S.-C. The Prophylactic Effects of Glutamine on Muscle Protein Synthesis and Degradation in Rats with Ethanol-Induced Liver Damage. Nutrients 2021, 13, 2788. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082788
Xiao Q, Chen Y-H, Pratama SA, Chen Y-L, Shirakawa H, Peng H-C, Yang S-C. The Prophylactic Effects of Glutamine on Muscle Protein Synthesis and Degradation in Rats with Ethanol-Induced Liver Damage. Nutrients. 2021; 13(8):2788. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082788
Chicago/Turabian StyleXiao, Qian, Yi-Hsiu Chen, Satwika Arya Pratama, Ya-Ling Chen, Hitoshi Shirakawa, Hsiang-Chi Peng, and Suh-Ching Yang. 2021. "The Prophylactic Effects of Glutamine on Muscle Protein Synthesis and Degradation in Rats with Ethanol-Induced Liver Damage" Nutrients 13, no. 8: 2788. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082788