
Effects of Short-Term Phosphate Loading on Aerobic Capacity under Acute Hypoxia in Cyclists: A Randomized, Placebo-Controlled, Crossover Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
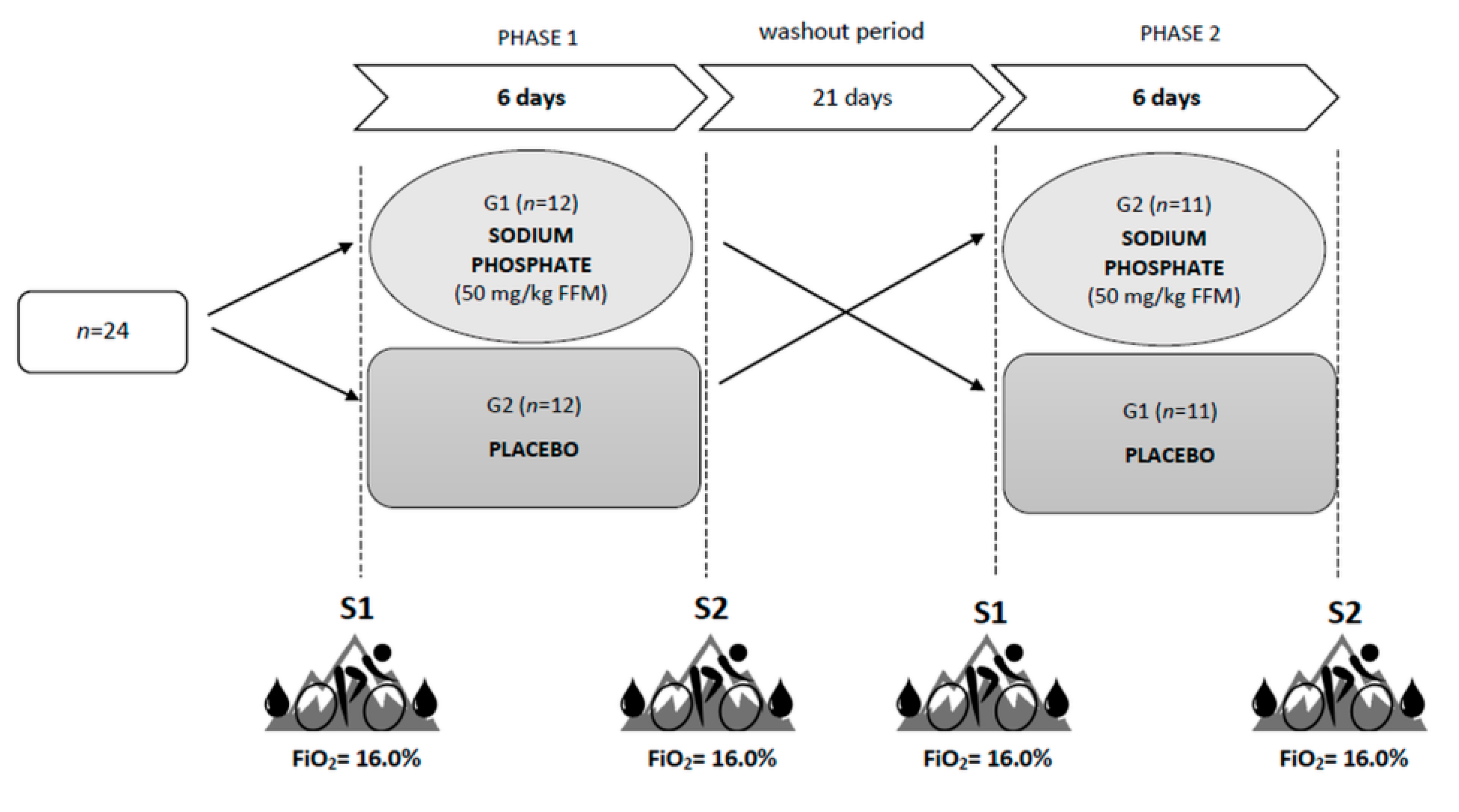
2.2. Study Design
2.3. Incremental Exercise Test
2.4. Determination of 2,3-DPG and HIF-1α
2.5. Supplementation with Sodium Phosphate
2.6. Statistical Analysis
3. Results
3.1. Exercise Performance and Cardiorespiratory Variables
3.2. Acid-Base Balance and Blood Gas Analysis
3.3. 2,3-Diphosphoglycerate (2,3-DPG) and Hypoxia-Inducible Factor 1 alpha (HIF-1α)
3.4. Calcium–Phosphate Balance
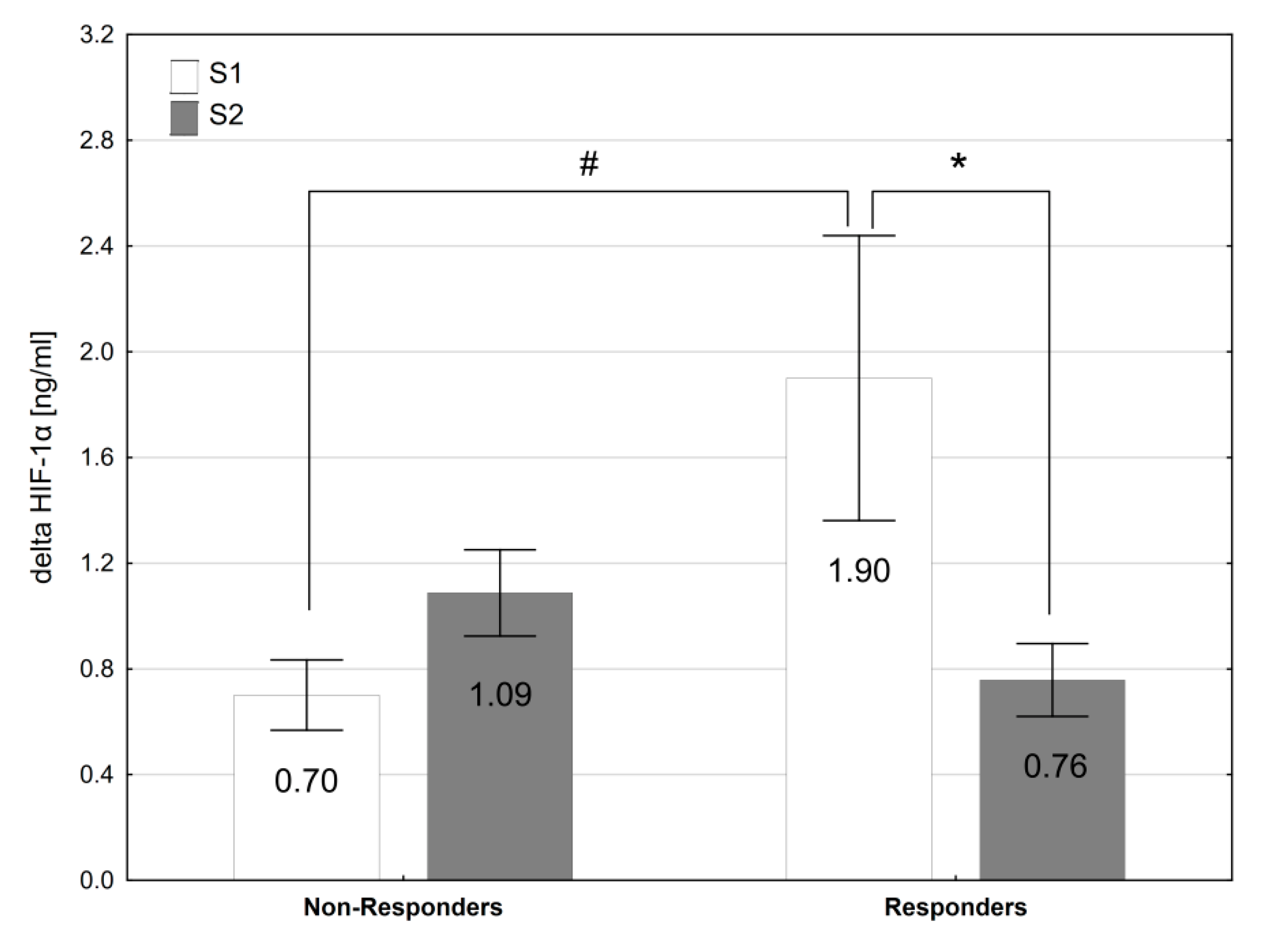
3.5. Responders vs. Non-Responders
4. Discussion
5. Practical Applications and Future Research
6. Study Limitations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peltonen, J.E.; Rusko, H.K.; Rantamaki, J.; Sweins, K.; Nittymaki, S.; Vitasalo, J.T. Effects of oxygen fraction in inspired air on force production and electromyogram activity during ergometer rowing. Eur. J. Appl. Physiol. 1997, 76, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Fulco, C.S.; Rock, P.B.; Cymerman, A. Maximal and submaximal exercise performance at altitude. Aviat. Space Environ. Med. 1998, 69, 793–801. [Google Scholar] [PubMed]
- Chapman, R.F.; Stager, J.M.; Tanner, D.A.; Stray-Gundersen, J.; Levine, B.D. Impairment of 3000-m run time at altitude is influenced by arterial oxyhemoglobin saturation. Med. Sci. Sports Exerc. 2011, 43, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Płoszczyca, K.; Czuba, M.; Chalimoniuk, M.; Gajda, R.; Baranowski, M. Red blood cell 2,3-diphosphoglycerate decreases in response to a 30 km time trial under hypoxia in cyclists. Front. Physiol. 2021, 12, 670977. [Google Scholar] [CrossRef]
- Wehrlin, J.P.; Hallen, J. Linear decrease in VO2max and performance with increasing altitude in endurance athletes. Eur. J. Appl. Physiol. 2006, 96, 404–412. [Google Scholar] [CrossRef]
- Dempsey, J.A.; Wagner, P.D. Exercise-induced arterial hypoxemia. J. Appl. Physiol. 1999, 87, 1997–2006. [Google Scholar] [CrossRef] [PubMed]
- Amann, M.; Eldridge, M.W.; Lovering, A.T.; Stickland, M.K.; Pegelow, D.F.; Dempsey, J.A. Arterial oxygenation influences central motor output and exercise performance via effects on peripheral locomotor muscle fatigue in humans. J. Physiol. 2006, 575, 937–952. [Google Scholar] [CrossRef]
- Clark, S.A.; Bourdon, P.C.; Schmidt, W.; Singh, B.; Cable, G.; Onus, K.J.; Woolford, S.M.; Stanef, T.; Gore, C.J.; Aughey, R.J. The effect of acute simulated moderate altitude on power, performance and pacing strategies in well-trained cyclists. Eur. J. Appl. Physiol. 2007, 102, 45–55. [Google Scholar] [CrossRef]
- Saugy, J.J.; Rupp, T.; Faiss, R.; Lamon, A.; Bourdillon, N.; Millet, G.P. Cycling time trial is more altered in hypobaric than normobaric hypoxia. Med. Sci. Sports Exerc. 2016, 648, 680–688. [Google Scholar] [CrossRef]
- Chapman, R.F. The individual response to training and competition at altitude. Br. J. Sports Med. 2013, 47, i40–i44. [Google Scholar] [CrossRef]
- Gore, C.J.; Hahn, A.G.; Scroop, G.C.; Watson, D.B.; Norton, K.I.; Wood, R.J.; Campbell, D.P.; Emonson, D.L. Increased arterial desaturation in trained cyclists during maximal exercise at 580 m altitude. J. Appl. Physiol. 1996, 80, 2204–2210. [Google Scholar] [CrossRef]
- Gore, C.J.; Little, S.C.; Hahn, A.G.; Scroop, G.C.; Norton, K.I.; Bourdon, P.C.; Woolford, S.M.; Buckley, J.D.; Stanef, T.; Campbell, D.P.; et al. Reduced performance of male and female athletes at 580 m altitude. Eur. J. Appl. Physiol. 1997, 75, 136–143. [Google Scholar] [CrossRef]
- Gore, C.J.; McSharry, P.E.; Hewett, A.J.; Saunders, P.U. Preparation for football competition at moderate to high altitude. Scan. J. Med. Sci. Sports 2008, 18, 85–95. [Google Scholar] [CrossRef]
- Chapman, R.F.; Laymon, A.S.; Levine, B.D. Timing of arrival and pre-acclimatization strategies for the endurance athlete competing at moderate to high altitudes. High Alt. Med. Biol. 2013, 14, 319–324. [Google Scholar] [CrossRef]
- Hahn, A.G.; Gore, C.J. The effect of altitude on cycling performance: A challenge to traditional concepts. Sports Med. 2001, 31, 533–557. [Google Scholar] [CrossRef]
- Schuler, B.; Thomsen, J.J.; Gassmann, M.; Lundby, C. Timing the arrival at 2340 m altitude for aerobic performance. Scand. J. Med. Sci. Sports 2007, 17, 588–594. [Google Scholar] [CrossRef]
- Michalczyk, M.; Czuba, M.; Zydek, G.; Zając, A.; Langfort, J. Dietary Recommendations for Cyclists during Altitude Training. Nutrients 2016, 8, 377. [Google Scholar] [CrossRef] [Green Version]
- Burtscher, M.; Niedermeier, M.; Burtscher, J.; Pesta, D.; Suchy, J.; Strasser, B. Preparation for Endurance Competitions at Altitude: Physiological, Psychological, Dietary and Coaching Aspects. A Narrative Review. Front. Physiol. 2018, 9, 1504. [Google Scholar] [CrossRef]
- Buck, C.L.; Wallman, K.E.; Dawson, B.; Guelfi, K.J. Sodium phosphate as an ergogenic aid. Sports Med. 2013, 43, 425–435. [Google Scholar] [CrossRef]
- Cade, R.; Conte, M.; Zauner, C.; Mars, D.; Peterson, J.; Lunne, D.; Hommen, N.; Packer, D. Effects of phosphate loading on 2,3-diphosphoglycerate and maximal oxygen uptake. Med. Sci. Sports Exerc. 1984, 16, 263–268. [Google Scholar] [CrossRef]
- Kreider, R.; Miller, G.W.; Williams, M.H.; Somma, C.T.; Nasser, T.A. Effects of phosphate loading on oxygen uptake, ventilatory anaerobic threshold, and run performance. Med. Sci. Sports Exerc. 1990, 22, 250–256. [Google Scholar] [PubMed]
- Stewart, I.; McNaughton, L.; Davies, P.; Tristram, S. Phosphate loading and the effects on VO2max in trained cyclists. Res. Q. Exerc. Sport 1990, 61, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Czuba, M.; Zajac, A.; Poprzęcki, S.; Cholewa, J. The influence of sodium phosphate supplementation on VO2max, Serum 2,3-diphosphoglycerate level and heart rate in off-road cyclists. J. Hum. Kinet. 2008, 19, 149–164. [Google Scholar] [CrossRef]
- Czuba, M.; Zajac, A.; Poprzęcki, S.; Cholewa, J.; Woska, S. Effects of sodium phosphate loading on aerobic power and capacity in off road cyclists. J. Sports Sci. Med. 2009, 8, 591–599. [Google Scholar]
- Brewer, C.P.; Dawson, B.; Wallman, K.E.; Guelfi, K.J. Effect of repeated sodium phosphate loading on cycling time-trial performance and VO2peak. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 187–194. [Google Scholar] [CrossRef]
- Bredle, D.L.; Stager, J.M.; Brechue, W.F.; Farber, M.O. Phosphate supplementation, cardiovascular function, and exercise performance in humans. J. Appl. Physiol. 1988, 65, 1821–1826. [Google Scholar] [CrossRef]
- West, J.S.; Ayton, T.; Wallman, K.E.; Guelfi, K.J. The effect of 6 days of sodium phosphate supplementation on appetite, energy intake, and aerobic capacity in trained men and women. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 422–429. [Google Scholar] [CrossRef] [Green Version]
- Buck, C.L.; Dawson, B.; Guelfi, K.; McNaughton, L.; Wallman, K.E. Sodium phosphate supplementation and time trial performance in female cyclists. J. Sports Sci. Med. 2014, 13, 469–475. [Google Scholar]
- Brown, J.A.; Glaister, M. The Effects of sodium phosphate supplementation on physiological responses to submaximal exercise and 20 km cycling time-trial performance. J. Diet. Suppl. 2019, 16, 564–575. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, D.H.; Smith, A.E.; Kendall, K.L.; Stout, J.R. Phosphate supplementation: An update. Strength Cond. J. 2010, 32, 53–56. [Google Scholar] [CrossRef]
- Currell, K.; Moore, D.R.; Peeling, P.; Burke, L.M.; Stear, S.J.; Castell, L.M. A–Z of nutritional supplements: Dietary supplements, sports nutrition foods and ergogenic aids for health and performance—Part 28. Br. J. Sports Med. 2012, 46, 75–76. [Google Scholar] [CrossRef] [Green Version]
- Lenfant, C.; Torrance, J.; English, E.; Finch, C.A.; Reynafarje, C.; Ramos, J.; Faura, J. Effect of altitude on oxygen binding by hemoglobin and on organic phosphate levels. J. Clin. Investig. 1968, 47, 2652–2656. [Google Scholar] [CrossRef] [Green Version]
- Mairbäurl, H.; Schobersberger, W.; Humpeler, E.; Hasibeder, W.; Fischer, W.; Raas, E. Beneficial effects of exercising at moderate altitude on red cell oxygen transport and on exercise performance. Pflug. Arch. 1986, 406, 594–599. [Google Scholar] [CrossRef]
- Koistinen, P.O.; Rusko, H.; Irjala, K.; Rajamäki, A.; Penttinen, K.; Sarparanta, V.P.; Karpakka, J.; Leppäluoto, J. EPO, red cells, and serum transferrin receptor in continuous and intermittent hypoxia. Med. Sci. Sports Exerc. 2000, 32, 800–804. [Google Scholar] [CrossRef]
- Moore, L.G.; Brewer, G.J. Beneficial effect of rightward hemoglobin-oxygen dissociation curve shift for short term high altitude adaptation. J. Lab. Clin. Med. 1981, 98, 145–154. [Google Scholar]
- Jain, S.C.; Singh, M.V.; Rawal, S.B.; Sharma, V.M.; Divekar, H.M.; Tyagi, A.K.; Panwar, M.R.; Swamy, Y.V. Effect of phosphate supplementation on oxygen delivery at high altitude. Int. J. Biometeorol. 1987, 31, 249–257. [Google Scholar] [CrossRef]
- Moore, L.G.; Brewer, G.J.; Oelshlegel, F.J.; Rose, A.M. Pharmacological Stimulation of Red Blood Cell Metabolism for High Altitude Preadaptation. In Oxygen Transport to Tissue. Advances in Experimental Medicine and Biology; Bruley, D.F., Bicher, H.I., Eds.; Springer: New York, NY, USA, 1973; pp. 693–698. [Google Scholar]
- Płoszczyca, K.; Gajda, R.; Czuba, M. The Effects of Sodium Phosphate Supplementation on the Cardiorespiratory System and Gross Efficiency during Exercise under Hypoxia in Male Cyclists: A Randomized, Placebo-Controlled, Cross-Over Study. Nutrients 2021, 13, 3556. [Google Scholar] [CrossRef]
- Urbaniak, G.C.; Plous, S. Research Randomizer (Version 4.0) [Computer Software]. Available online: http://www.randomizer.org/ (accessed on 14 September 2020).
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Edvardsen, E.; Hem, E.; Anderssen, S.A. End criteria for reaching maximal oxygen uptake must be strict and adjusted to sex and age: A cross-sectional study. PLoS ONE 2014, 9, e85276. [Google Scholar]
- Kuipers, H.; Verstappen, F.T.J.; Keizer, H.A.; Guerten, P.; van Kranenburg, G. Variability of aerobic performance in the laboratory and its physiological correlates. Int. J. Sports Med. 1985, 6, 197–201. [Google Scholar] [CrossRef]
- Cheng, B.; Kuipers, H.; Snyder, A.C.; Keizer, H.A.; Jeukendrup, A.; Hesselink, M. A New Approach for the determination of ventilatory and lactate thresholds. Int. J. Sports Med. 1992, 13, 518–522. [Google Scholar] [CrossRef]
- Czuba, M.; Zajac, A.; Cholewa, J.; Poprzęcki, S.; Waśkiewicz, Z.; Mikołajec, K. Lactate threshold (D-Max Method) and maximal lactate steady state in cyclists. J. Hum. Kinet. 2009, 21, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Płoszczyca, K.; Jazic, D.; Piotrowicz, Z.; Chalimoniuk, M.; Langfort, J.; Czuba, M. Comparison of maximal lactate steady state with anaerobic threshold determined by various methods based on graded exercise test with 3-min stages in elite cyclists. BMC Sports Sci. Med. Rehabil. 2020, 12, 70. [Google Scholar] [CrossRef]
- Van Beaumont, W. Evaluation of hemoconcentration from hematocrit measurements. J. Appl. Physiol. 1972, 32, 712–713. [Google Scholar] [CrossRef]
- Tremblay, M.S.; Galloway, S.D.; Sexsmith, J.R. Ergogenic effects of phosphate loading: Physiological fact or methodological fiction? Can. J. Appl. Physiol. 1994, 19, 1–11. [Google Scholar] [CrossRef]
- Galloway, S.D.R.; Tremblay, M.S.; Sexsmith, J.R.; Roberts, C.J. The effects of acute phosphate supplementation in subjects of different aerobic fitness levels. Eur. J. Appl. Physiol. Occup. Physiol. 1996, 72, 224–230. [Google Scholar] [CrossRef]
- Hopkins, W.; Marshall, S.; Batterham, A.; Hanin, J. Progressive statistics for studies in sports medicine and exercise science. Med. Sci. Sports Exerc. 2009, 41, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, W.G.; Schabort, E.J.; Hawley, J.A. Reliability of Power in Physical Performance Tests. Sports Med. 2001, 31, 211–234. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.G.; Hawley, J.A.; Burke, L.M. Design and analysis of research on sport performance enhancement. Med. Sci. Sports Exerc. 1999, 31, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Paton, C.D.; Hopkins, W.G. Variation in performance of elite cyclists from race to race. Eur. J. Sport Sci. 2006, 6, 25–31. [Google Scholar] [CrossRef]
- Benesch, R.; Benesch, R.E. The effect of organic phosphates from the human erythrocyte on the allosteric properties of hemoglobin. Biochem. Biophys. Res. Commun. 1967, 26, 162–167. [Google Scholar] [CrossRef]
- Duhm, J.; Gerlach, E. On the mechanisms of the hypoxia-induced increase of 2,3-diphosphoglycerate in erythrocytes. Pflug. Arch. 1971, 326, 254–269. [Google Scholar] [CrossRef]
- Lichtman, M.A.; Miller, D.R.; Cohen, J.; Waterhouse, C. Reduced red cell glycolysis, 2,3-diphosphoglycerate and adenosine triphosphate concentration, and increased hemoglobin-oxygen affinity caused by hypophosphatemia. Ann. Intern. Med. 1971, 74, 562–568. [Google Scholar] [CrossRef]
- Kreider, R.; Miller, G.W.; Schenck, D.; Cortes, C.W.; Miriel, V.; Somma, C.T.; Rowland, P.; Turner, C.; Hill, D. Effects of phosphate loading on metabolic and myocardial responses to maximal and endurance exercise. Int. J. Sport Nutr. 1992, 2, 20–47. [Google Scholar] [CrossRef]
- Goss, F.; Robertson, R.; Riechman, S.; Zoeller, R.; Dabayebeh, I.D.; Moyna, N.; Boer, N.; Peoples, J.; Metz, K. Effect of potassium phosphate supplementation on perceptual and physiological responses to maximal graded exercise. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 53–62. [Google Scholar] [CrossRef]
- Mairbäurl, H.; Schobersberger, W.; Hasibeder, W.; Schwaberger, G.; Gaesser, G.; Tanaka, K.R. Regulation of red cell 2,3-DPG and Hb-O2-affinity during acute exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1986, 55, 174–180. [Google Scholar] [CrossRef]
- Farber, O.M.; Sullivan, T.Y.; Fineberg, N.; Carlone, S.; Manfredi, F. Effect of decreased O2 affinity of hemoglobin on work performance during exercise in healthy humans. J. Lab. Clin. Med. 1984, 104, 166–175. [Google Scholar]
- Kreider, R.B. Phosphate supplementation in exercise and sport. In Macroelements, Water and Electrolytes in Sport Nutrition; Driskell, J.A., Wolinsky, I., Eds.; CRC Press: Boca Raton, FL, USA, 1999; p. 29201346. [Google Scholar]
- Levine, B.D.; Stray-Gundersen, J.; Mehta, R.D. Effect of altitude on football performance. Scand. J. Med. Sci. Sports 2008, 18, 76–84. [Google Scholar] [CrossRef]
- Swenson, E.R. Hypoxia and Its Acid-Base Consequences: From Mountains to Malignancy. Adv. Exp. Med. Biol. 2016, 903, 301–323. [Google Scholar]
- Sumi, D.; Kasai, N.; Ito, H.; Goto, K. The Effects of Endurance Exercise in Hypoxia on Acid-Base Balance, Potassium Kinetics, and Exogenous Glucose Oxidation. Front. Physiol. 2019, 10, 504. [Google Scholar] [CrossRef]
- Dempsey, J.A.; Forster, H.V. Mediation of Ventilatory Adaptations. Physiol. Rev. 1982, 62, 262–346. [Google Scholar] [CrossRef]
- Ge, R.-L.; Babb, T.G.; Sivieri, M.; Resaland, G.K.; Karlsen, T.; Stray-Gundersen, J.; Levine, B.D. Urine acid-base compensation at simulated moderate altitude. High Alt. Med. Biol. 2006, 7, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Lühker, O.; Berger, M.M.; Pohlmann, A.; Hotz, L.; Gruhlke, T.; Hochreiter, M. Changes in acid–base and ion balance during exercise in normoxia and normobaric hypoxia. Eur. J. Appl. Physiol. 2017, 117, 2251–2261. [Google Scholar] [CrossRef]
- Hauser, T.; Bartsch, D.; Baumgärtel, L.; Schulz, H. Reliability of maximal lactate-steady-state. Int. J. Sports Med. 2013, 34, 196–199. [Google Scholar] [CrossRef]
- Mann, T.; Lamberts, R.P.; Lambert, M.I. Methods of prescribing relative exercise intensity: Physiological and practical considerations. Sports Med. 2013, 43, 613–625. [Google Scholar] [CrossRef]
- Vella, C.A.; Robergs, R.A. A review of the stroke volume response to upright exercise in healthy subjects. Br. J. Sports Med. 2005, 39, 190–195. [Google Scholar] [CrossRef]
- Mazaheri, R.; Schmied, C.; Niederseer, D.; Guazzi, M. Cardiopulmonary Exercise Test Parameters in Athletic Population: A Review. J. Clin. Med. 2021, 10, 5073. [Google Scholar] [CrossRef]
- Skattebo, Ø.; Calbet, J.A.L.; Rud, B.; Capelli, C.; Hallén, J. Contribution of oxygen extraction fraction to maximal oxygen uptake in healthy young men. Acta Physiol. 2020, 230, e13486. [Google Scholar] [CrossRef]
- Stroka, D.M.; Burkhardt, T.; Desbaillets, I.; Wenger, R.H.; Neil, D.A.H.; Bauer, C.; Gassmann, M.; Candinas, D. HIF-1 is expressed in normoxic tissue and displays an organ specific regulation under systemic hypoxia. FASEB J. 2001, 15, 2445–2453. [Google Scholar] [CrossRef] [Green Version]
- Lemieux, P.; Birot, O. Altitude, Exercise, and Skeletal Muscle Angio-Adaptive Responses to Hypoxia: A Complex Story. Front. Physiol. 2021, 12, 735557. [Google Scholar] [CrossRef] [PubMed]
- Ameln, H.; Gustafsson, T.; Sundberg, C.J.; Okamoto, K.; Jansson, E.; Poellinger, L.; Makino, Y. Physiological activation of hypoxia inducible factor-1 in human skeletal muscle. FASEB J. 2005, 19, 1009–1011. [Google Scholar] [CrossRef]
- Van Thienen, R.; Masschelein, E.; D’Hulst, G.; Thomis, M.; Hespel, P. Twin Resemblance in Muscle HIF-1α Responses to Hypoxia and Exercise. Front. Physiol. 2017, 7, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Smet, S.; D’Hulst, G.; Poffé, C.; Van Thienen, R.; Berardi, E.; Hespel, P. High-intensity interval training in hypoxia does not affect muscle HIF responses to acute hypoxia in humans. Eur. J. Appl. Physiol. 2018, 118, 847–862. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, M.E.; Rundqvist, H. Skeletal muscle hypoxia-inducible factor-1 and exercise. Exp. Physiol. 2016, 101, 28–32. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | Sodium Phosphate | Placebo | ||
|---|---|---|---|---|
| Before (S1) | After (S2) | Before (S1) | After (S2) | |
| WRmax (W) | 310.7 ± 6.2 | 314.7 ± 6.3 | 303.5 ± 6.2 | 307.5 ± 6.3 |
| WRLT (W) | 227.3 ± 5.4 | 233.6 ** ± 5.7 | 228.2 ± 5.4 | 227.3 ± 5.7 |
| VO2max (L·min−1) | 3.49 ± 0.06 | 3.54 ± 0.06 | 3.53 ± 0.06 | 3.49 ± 0.06 |
| HRmax (bpm) | 178 ± 2.3 | 177 ± 2.3 | 178 ± 2.3 | 178 ± 2.3 |
| SVmax (ml) | 108.4 ± 2.2 | 108.7 ± 2.3 | 108.3 ± 2.2 | 110.1 ± 2.4 |
| Qmax (L·min−1) | 19.3 ± 0.3 | 19.2 ± 0.3 | 19.5 ± 0.3 | 19.5 ± 0.3 |
| VEmax (L·min−1) | 170.5 ± 3.1 | 169.6 ± 3.0 | 168.5 ± 3.1 | 171.5 ± 3.0 |
| VO2/HRmax (ml·bpm−1) | 19.6 ± 0.4 | 20.0 ± 0.4 | 19.9 ± 0.4 | 19.6 ± 0.4 |
| Variables | Sodium Phosphate | Placebo | ||
|---|---|---|---|---|
| Before (S1) | After (S2) | Before (S1) | After (S2) | |
| ∆ LA (mmol·L−1) | 11.78 ± 0.55 | 12.76 ± 0.50 | 12.09 ± 0.55 | 12.50 ± 0.50 |
| ∆ pH | −0.187 ± 0.013 | −0.208 ± 0.013 | −0.174 ± 0.013 | −0.197 ± 0.013 |
| ∆ BE (mmol·L−1) | −14.05 ± 0.61 | −14.65 ± 0.64 | −13.35 ± 0.61 | −14.30 ± 0.64 |
| ∆ HCO3 (mmol·L−1) | −12.49 ± 0.45 | −12.62 ± 0.46 | −11.96 ± 0.45 | −12.49 ± 0.46 |
| ∆ sO2 (%) | −5.13 ± 0.99 | −5.72 ± 0.74 | −5.12 ± 0.99 | −5.48 ± 0.74 |
| ∆ pO2 (mmHg) | −0.08 ± 1.57 | −0.10 ± 1.25 | −0.10 ± 1.53 | −0.55 ± 1.22 |
| ∆ pCO2 (mmHg) | −8.15 ± 0.62 | −7.02 ± 0.54 | −8.25 ± 0.62 | −7.75 ± 0.54 |
| ∆ p50 (mmHg) | 5.03 ± 0.45 | 5.42 ± 0.45 | 4.49 ± 0.45 | 5.22 ± 0.45 |
| Variables | Sodium Phosphate | Placebo | ||
|---|---|---|---|---|
| Before (S1) | After (S2) | Before (S1) | After (S2) | |
| Pi rest (mg·dL−1) | 3.10 ± 0.07 | 3.38 * ± 0.08 | 3.28 ± 0.07 | 3.30 ± 0.08 |
| Ca rest (mg·dL−1) | 9.54 ± 0.06 | 9.51 ± 0.06 | 9.52 ± 0.06 | 9.46 ± 0.06 |
| PTH rest (pg·mL−1) | 30.36 ± 1.97 | 31.25 ± 2.14 | 30.32 ± 1.97 | 30.49 ± 2.14 |
| 2,3-DPG rest (mmol·L er.−1) | 6.19 ± 0.51 | 7.29 ± 0.36 | 7.03 ± 0.54 | 6.45 ± 0.38 |
| 2,3-DPG max (mmol·L er.−1) | 8.61 # ± 1.17 | 9.80 # ± 0.86 | 11.86 ### ± 1.29 | 10.03 ### ± 0.85 |
| HIF-1α rest (ng·mL−1) | 1.66 ± 0.39 | 1.90 ± 0.42 | 2.12 ± 0.40 | 2.30 ± 0.43 |
| HIF-1α max (ng·mL−1) | 2.96 ### ± 0.57 | 2.82 ### ± 0.51 | 3.38 ### ± 0.59 | 3.87 ### ± 0.49 |
| Variables | Responders (n = 11) | Non-Responders (n = 11) | ||
|---|---|---|---|---|
| Before (S1) | After (S2) | Before (S1) | After (S2) | |
| WRmax (W) | 307.6 ± 9.2 | 316.1 *** ± 9.2 | 313.8 ± 9.2 | 313.3 ± 9.2 |
| WRLT (W) | 223.6 ± 8.4 | 236.4 *** ± 9.0 | 230.9 ± 8.4 | 230.9 ± 9.0 |
| VO2max (L·min−1) | 3.43 ± 0.09 | 3.57 ** ± 0.08 | 3.54 ± 0.09 | 3.51 ± 0.08 |
| HRmax (bpm) | 184 # ± 2.7 | 184 ## ± 2.5 | 172 ± 2.7 | 171 ± 2.5 |
| SVmax (mL) | 103.7 ± 2.9 | 103.6 ± 3.2 | 112.4 ± 2.7 | 112.9 ± 2.9 |
| Qmax (L·min−1) | 19.3 ± 0.4 | 19.2 ± 0.4 | 19.2 ± 0.5 | 19.2 ± 0.5 |
| VEmax (L·min−1) | 170.4 ± 4.6 | 170.0 ± 4.3 | 170.6 ± 4.6 | 169.2 ± 4.3 |
| VO2/HRmax (ml·bpm−1) | 18.7 # ± 0.5 | 19.4 ** ± 0.5 | 20.6 ± 0.5 | 20.6 ± 0.5 |
| Variables | Responders | Non-Responders | ||
|---|---|---|---|---|
| Before (S1) | After (S2) | Before (S1) | After (S2) | |
| Pi (mg·dL−1) | 3.07 ± 0.10 | 3.44 ± 0.11 | 3.14 ± 0.10 | 3.31 ± 0.11 |
| Ca (mg·dL−1) | 9.58 ± 0.08 | 9.56 ± 0.09 | 9.50 ± 0.08 | 9.47 ± 0.09 |
| PTH (pg·mL−1) | 27.67 ± 2.69 | 30.58 ± 3.09 | 32.80 ± 2.57 | 31.85 ± 2.95 |
| 2,3-DPG (mmol·L er.−1) | 6.54 ± 0.57 | 7.39 ± 0.58 | 5.83 ± 0.57 | 7.19 ± 0.58 |
| HIF-1α (ng·mL−1) | 1.57 ± 0.52 | 1.83 ± 0.66 | 1.76 ± 0.52 | 1.96 ± 0.66 |
| Variables | Responders | Non-Responders | ||
|---|---|---|---|---|
| Before (S1) | After (S2) | Before (S1) | After (S2) | |
| RBC (T·L−1) | 5.02 ± 0.08 | 5.01 ± 0.09 | 4.88 ± 0.07 | 4.80 ± 0.08 |
| Ret (‰) | 12.7 ± 0.96 | 12.8 ± 0.65 | 12.6 ± 0.86 | 12.0 ± 0.59 |
| [Hb] (g·dL−1) | 14.86 ± 0.21 | 14.97 ± 0.27 | 14.68 ± 0.19 | 14.45 ± 0.24 |
| Hct (%) | 43.6 ± 0.54 | 43.8 ± 0.69 | 42.8 ± 0.49 | 42.3 ± 0.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płoszczyca, K.; Chalimoniuk, M.; Przybylska, I.; Czuba, M. Effects of Short-Term Phosphate Loading on Aerobic Capacity under Acute Hypoxia in Cyclists: A Randomized, Placebo-Controlled, Crossover Study. Nutrients 2022, 14, 236. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14020236
Płoszczyca K, Chalimoniuk M, Przybylska I, Czuba M. Effects of Short-Term Phosphate Loading on Aerobic Capacity under Acute Hypoxia in Cyclists: A Randomized, Placebo-Controlled, Crossover Study. Nutrients. 2022; 14(2):236. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14020236
Chicago/Turabian StylePłoszczyca, Kamila, Małgorzata Chalimoniuk, Iwona Przybylska, and Miłosz Czuba. 2022. "Effects of Short-Term Phosphate Loading on Aerobic Capacity under Acute Hypoxia in Cyclists: A Randomized, Placebo-Controlled, Crossover Study" Nutrients 14, no. 2: 236. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14020236