
Mannose-Modified Chitosan Poly(lactic-co-glycolic acid) Microspheres Act as a Mannose Receptor-Mediated Delivery System Enhancing the Immune Response

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Synthesis of Mannose-Modified Chitosan PLGA Microspheres
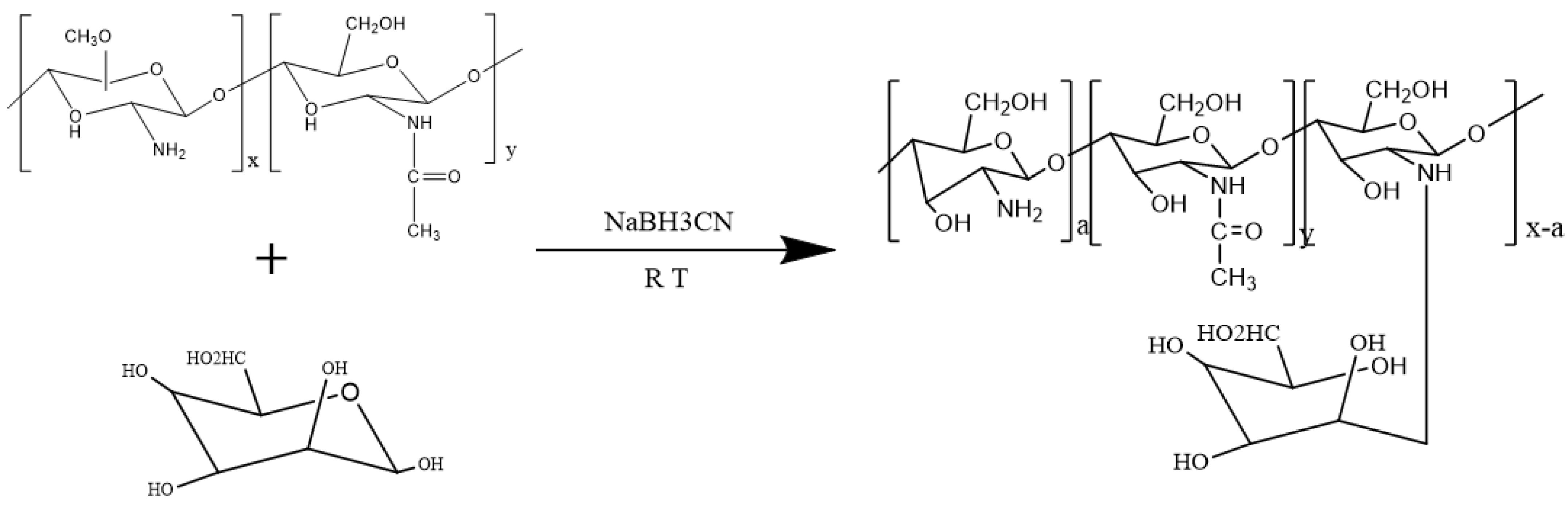
2.2.1. Synthesis of Mannose-Modified Chitosan
2.2.2. Physicochemical Property Analysis of MAN-CS
2.2.3. Synthesis of PLGA Microspheres
2.2.4. Determination of Drug Loading on Microspheres
2.2.5. Orthogonal Experiment Optimization
2.2.6. Synthesis of Mannose-Modified Chitosan PLGA Microspheres
2.2.7. Microsphere Surface Morphology, Particle Size, and Zeta Potential
2.3. In Vitro Experiments
2.3.1. Cytotoxicity of MAN-CS-OVA-PLGA-MPs on DCs
2.3.2. Phagocytosis of Different Microspheres by DC Cells
2.3.3. The Effect of MAN-CS-OVA-PLGA-MPs on the Delivery of OVA
2.4. In Vivo Experiments
2.4.1. Mice Grouping and Immunization
2.4.2. Immunophenotype of Spleen Lymphocytes in Mice
2.4.3. The Maturation of DCs in Mouse Spleen
2.4.4. Determination of OVA-Specific IgG and IgG Isotype in Serum of Mice
2.4.5. Detection of Cytokines in Serum of Immunized Mice by ELISA
2.5. Statistical Analysis
3. Results
3.1. Physicochemical Characteristics
3.1.1. Identification of Mannose-Modified Chitosan
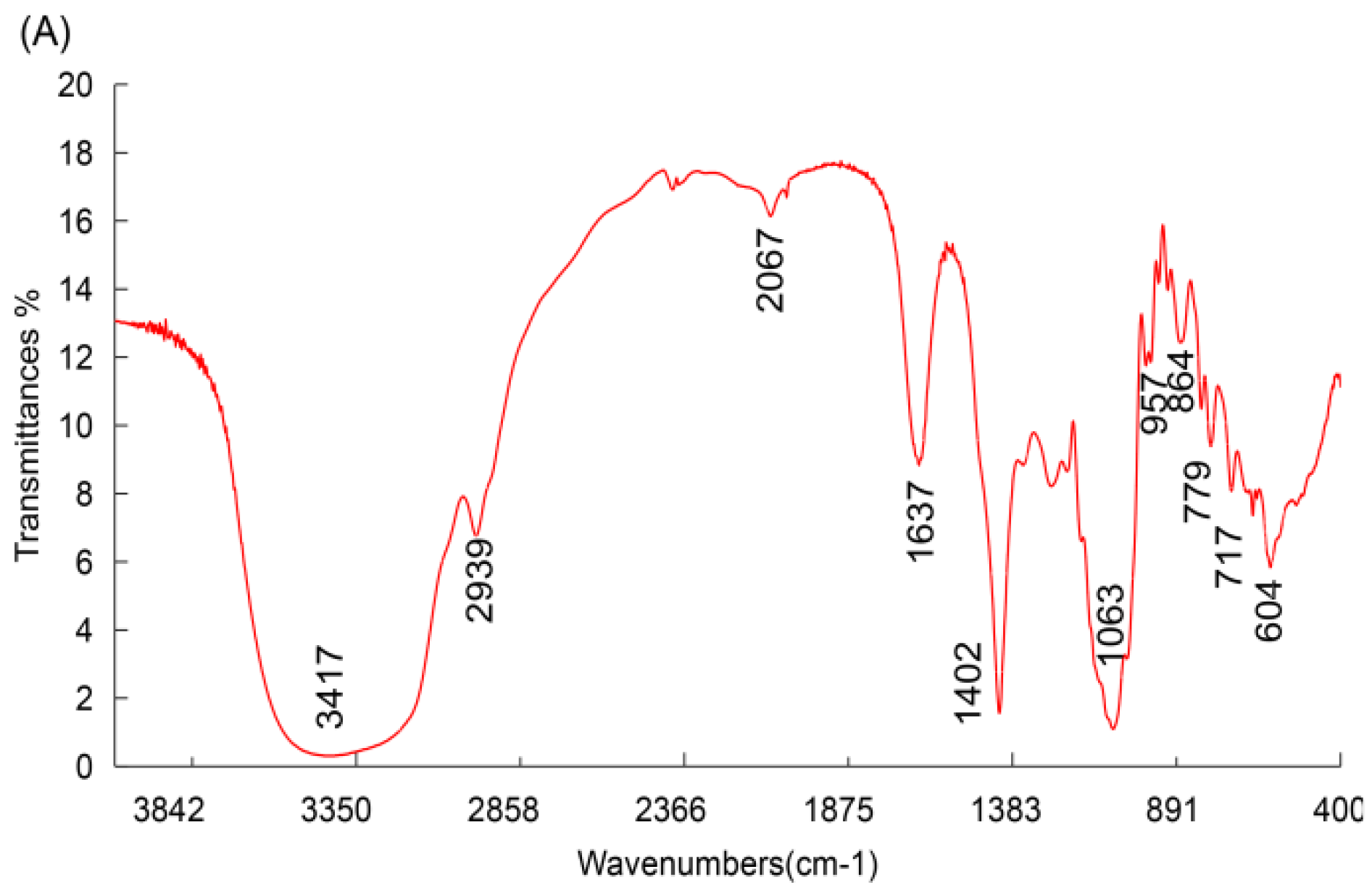
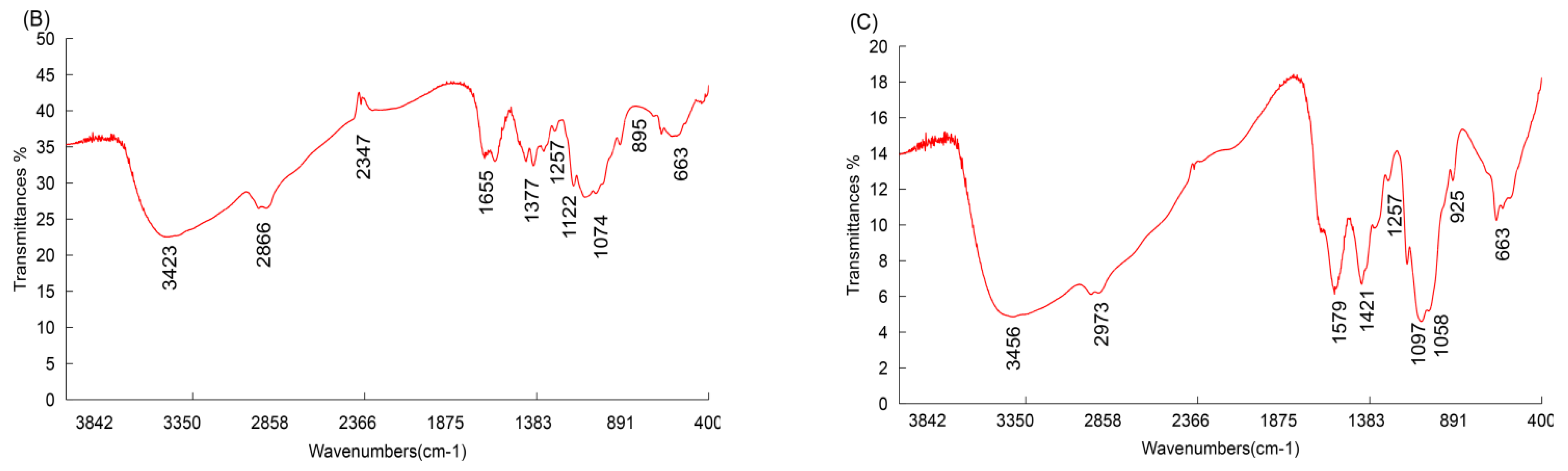
3.1.2. FT-IR Spectroscopy Analysis
3.2. Orthogonal Design of PLGA Microspheres
3.3. Morphology of the Microspheres Obtained by SEM
3.4. The Particle Size and Electromotive Force of Three Types of PLGA Microspheres
3.5. In Vitro Experiments
3.5.1. Cytotoxicity of MAN-CS-OVA-PLGA-MPs towards DCs
3.5.2. Effects of MAN-CS-OVA-PLGA-MPs on the Phagocytic Activity of DCs
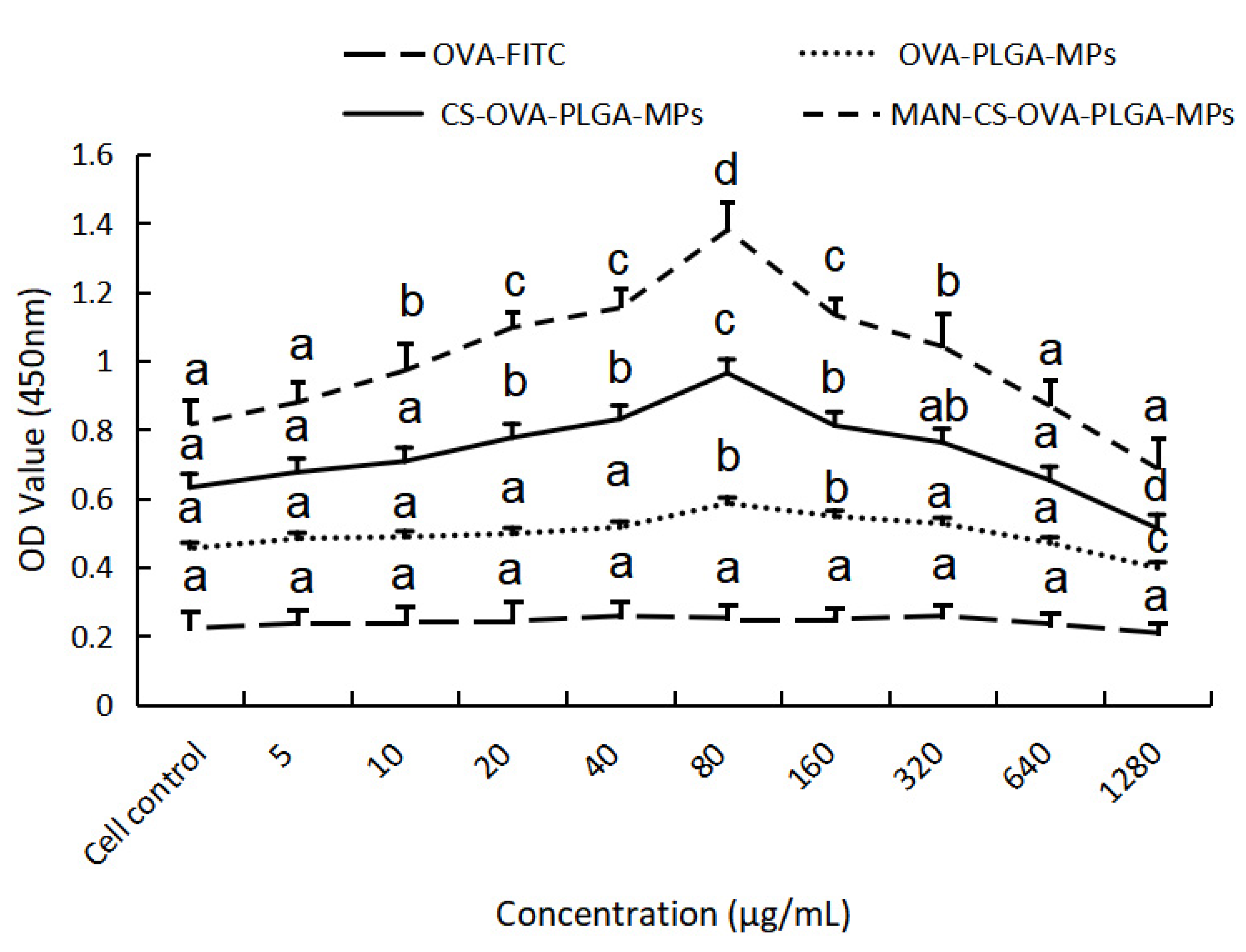
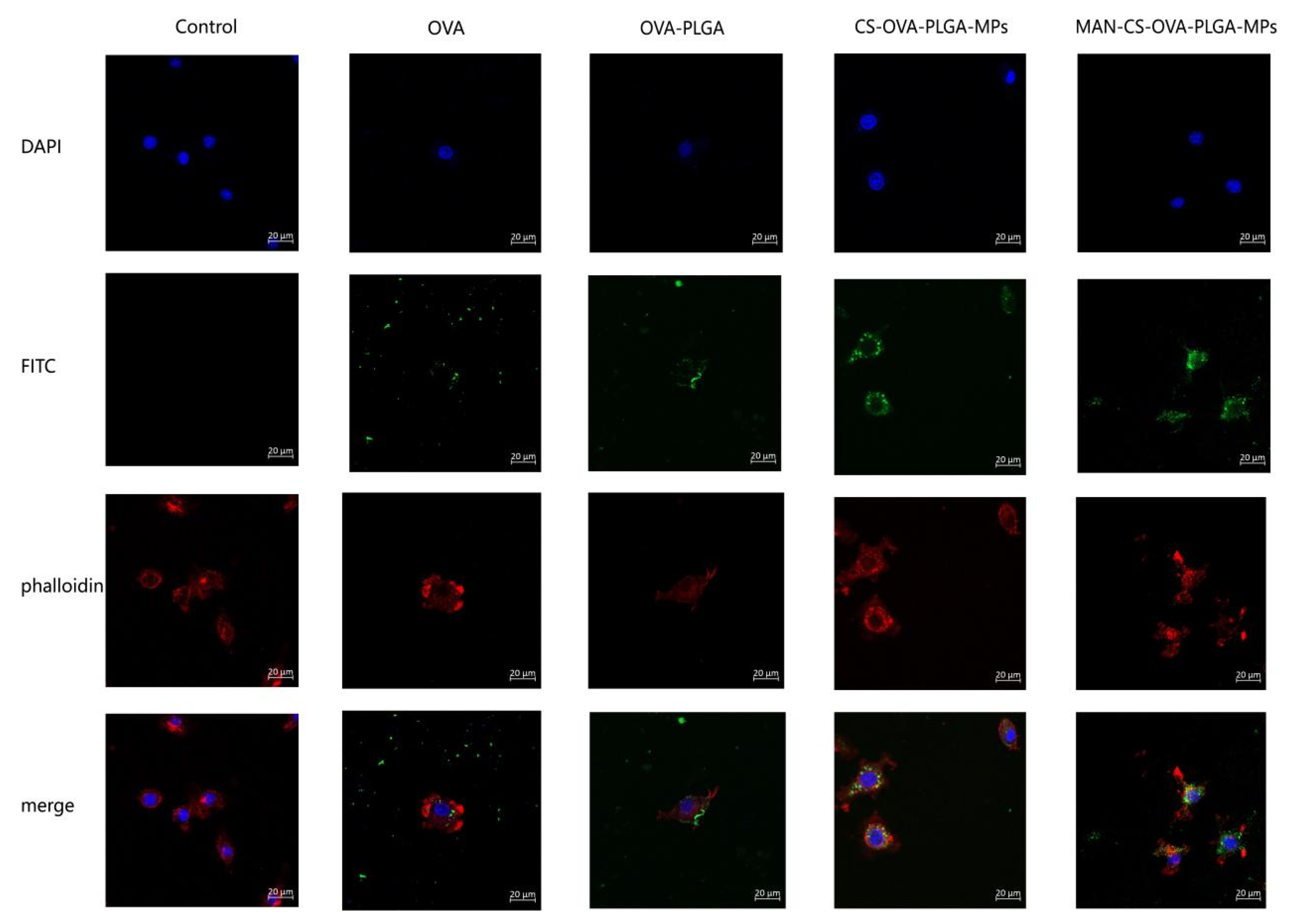
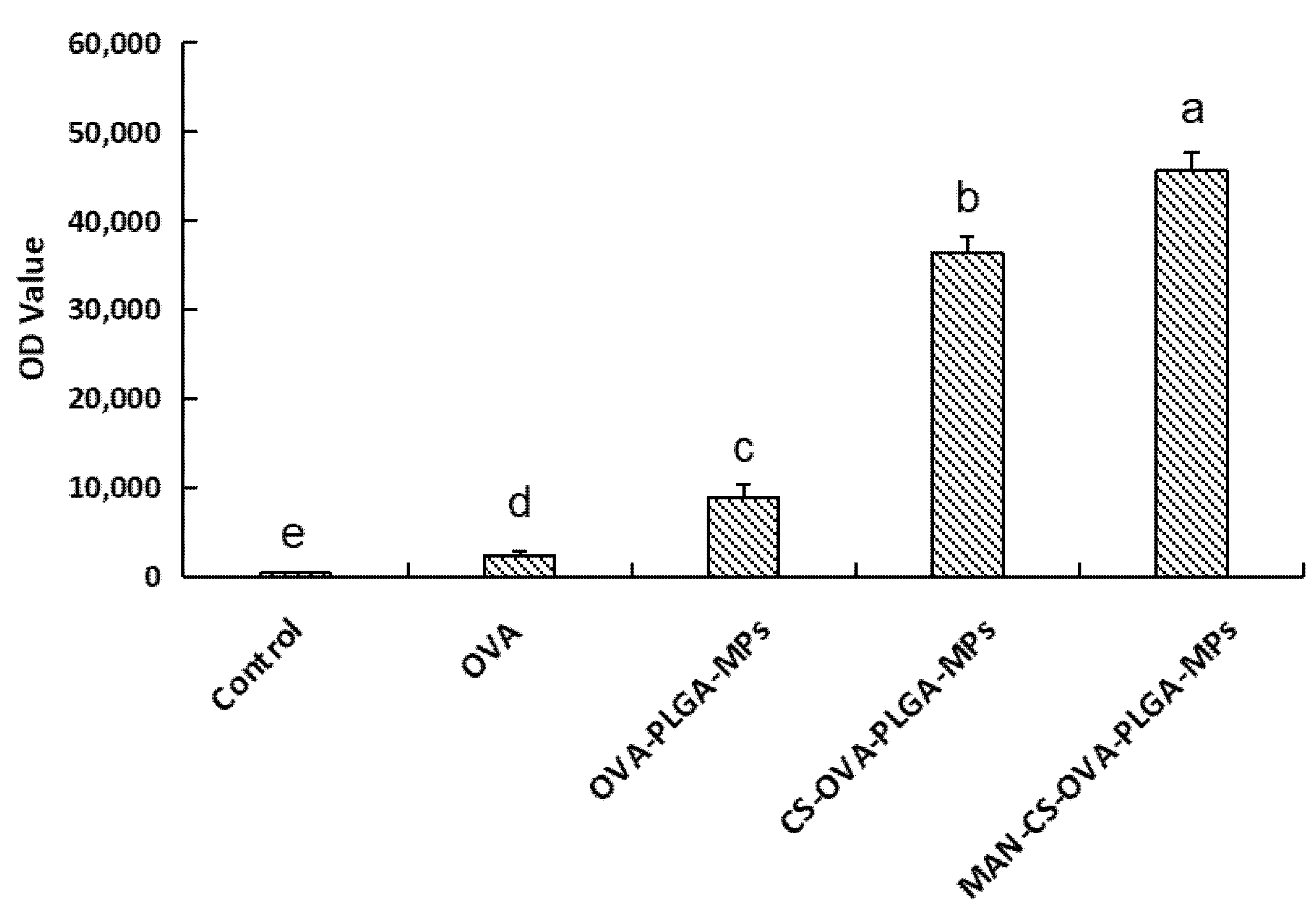
3.5.3. The Delivery Effect of MAN-CS-OVA-PLGA-MPs on OVA
3.6. In Vivo Experiments
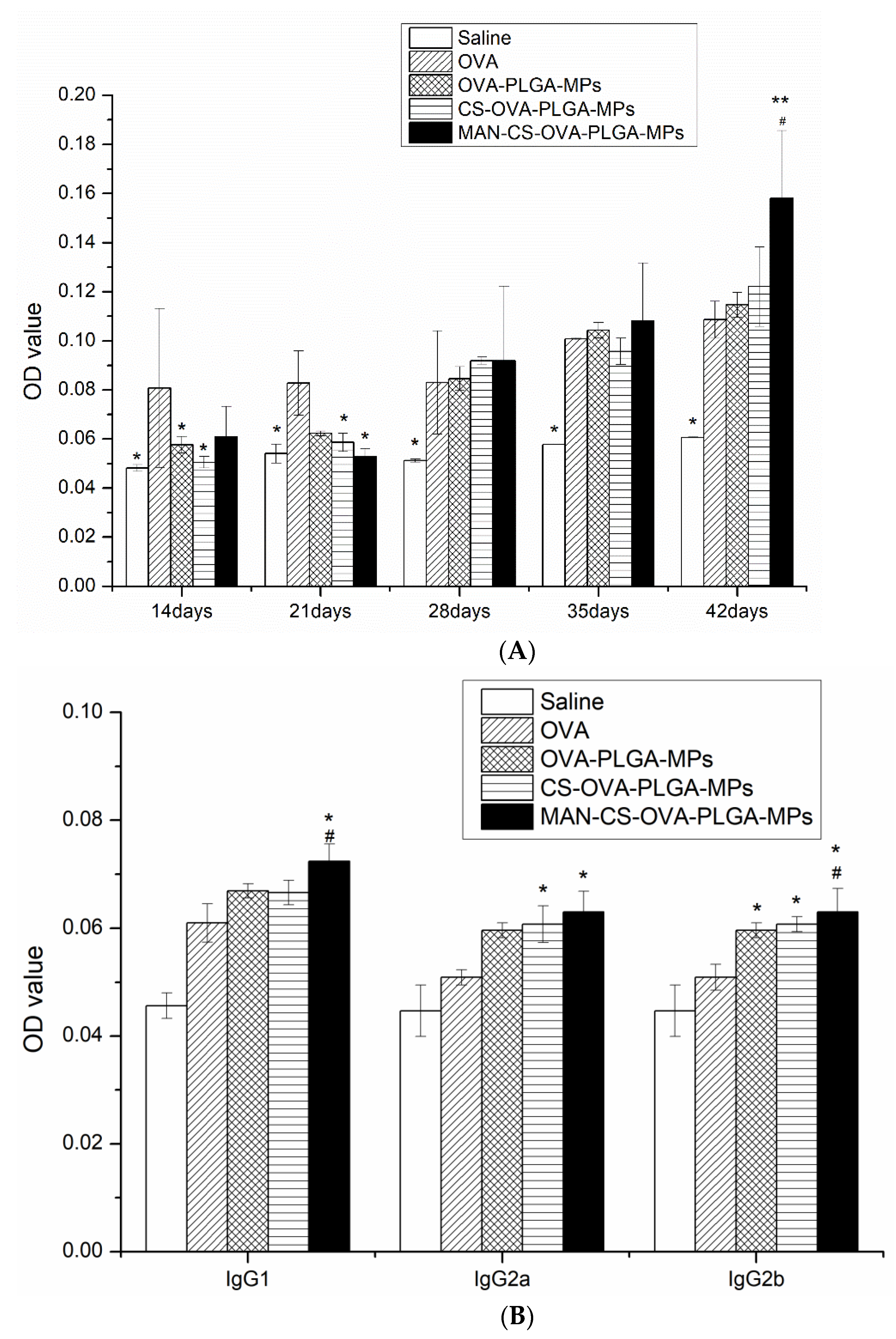
3.6.1. Effect of MAN-CS-OVA-PLGA-MPs on OVA-Specific IgG and IgG Isotype Antibody Titers of Murine Serum
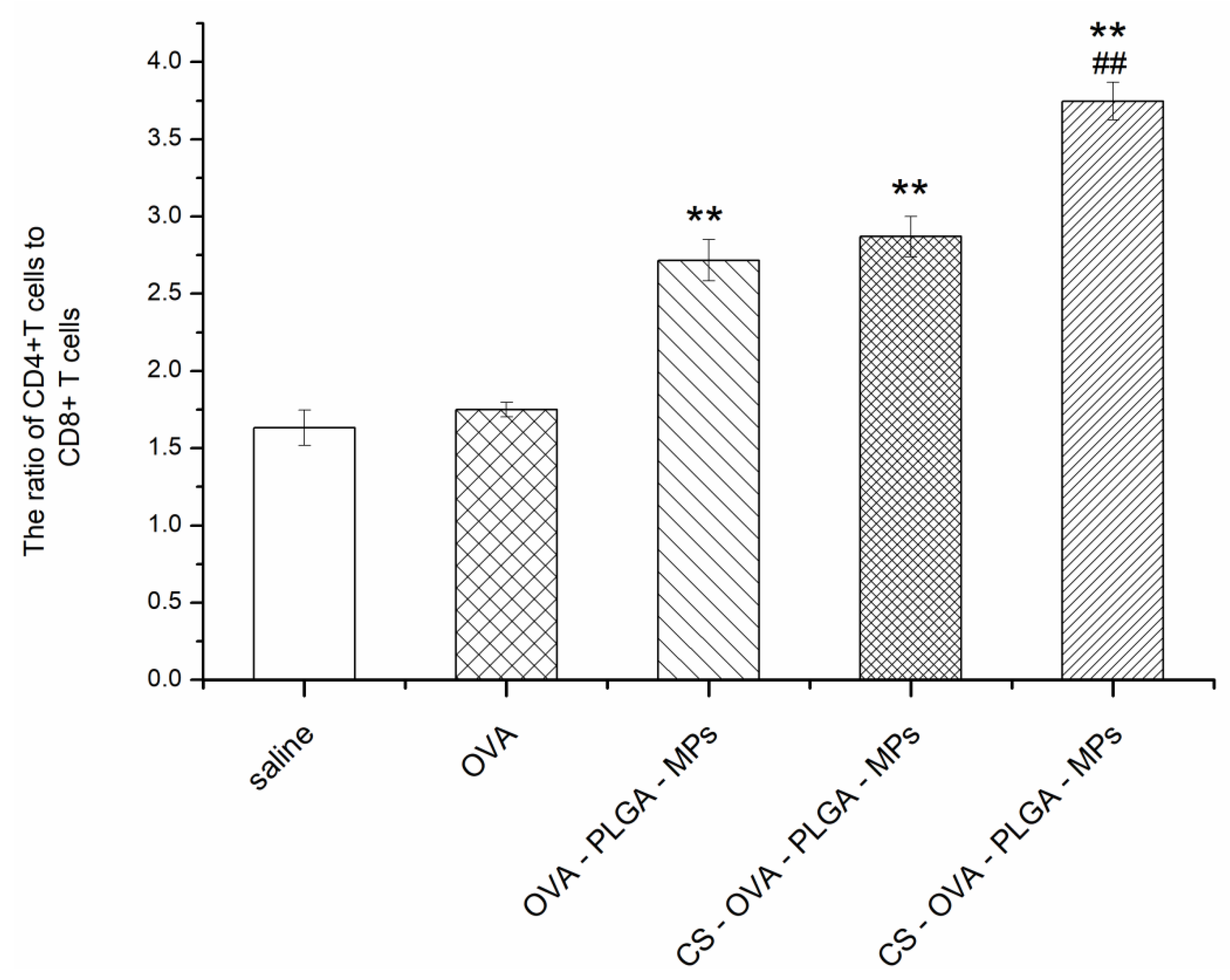
3.6.2. Effect of MAN-CS-OVA-PLGA-MPs on the Immunophenotype of Spleen Lymphocytes in Mice
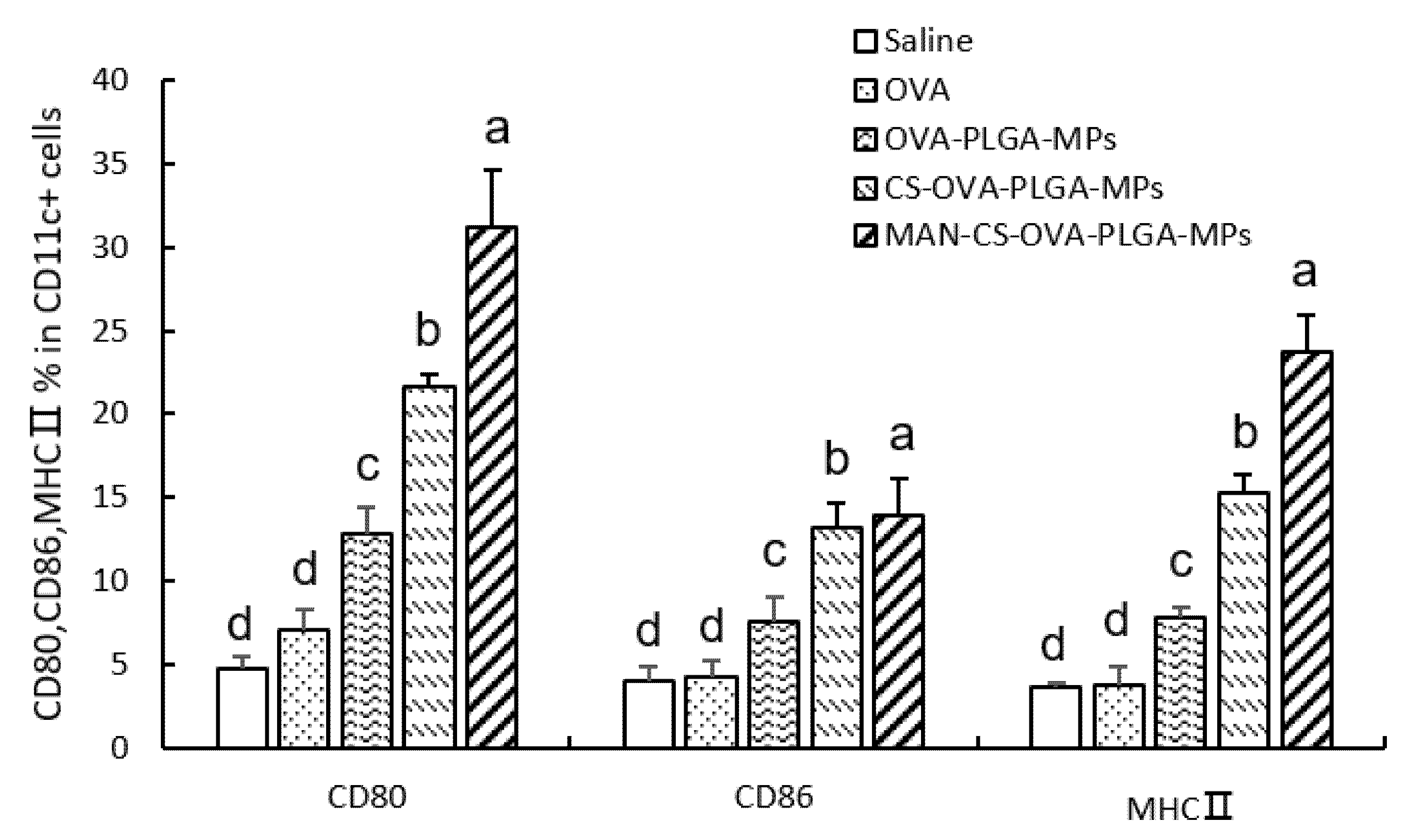
3.6.3. Effect of MAN-CS-OVA-PLGA-MPs on the Expression of DC Surface Molecules in the Spleen
3.6.4. Effect of MAN-CS-OVA-PLGA-MPs on the Concentrations of Cytokines in the Serum of Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hannah, B.M.; Joanna, L.T.; Mats, A.; Ed, C.L. Immunomodulatory properties of chitosan polymers. Biomaterials 2018, 184, 1–9. [Google Scholar]
- Muxika, A.; Etxabide, A.; Uranga, J.; Guerrero, P.; de la Caba, K. Chitosan as a bioactive polymer: Processing, properties and applications. Int. J. Biol. Macromol. 2017, 105, 1358–1368. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, Y.; Guan, L.; Zhang, Y.; Dang, Q.; Dong, P.; Li, J.; Liang, X.G. Preparation of astaxanthin-loaded DNA/chitosan nanoparticles for improved cellular uptake and antioxidation capability. Food Chem. 2017, 227, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, P.M.; Ignacimuthu, S.; RajivGandhi, M.; Shajahan, A.; Ganesan, P. Comparative studies of tripolyphosphate and glutaraldehyde cross-linked chitosan-botanical pesticide nanoparticles and their agricultural applications. Int. J. Biol. Macromol. 2017, 104, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Saad, Q.; Muhammad, Z.; Shariq, N.; Khurshid, Z.; Shah, A.; Husain, S.; Shehriar, H.; Ihtesham, R. Electrospinning of Chitosan-Based Solutions for Tissue Engineering and Regenerative Medicine. Int. J. Mol. Sci. 2018, 19, 407. [Google Scholar]
- Zheng, Y.; Yan, S.; Shi, B. Mechanism of antioxidative stress and anti-inflammatory action of chitosan and its derivatives. J. Anim. Nutr. 2018, 30, 1633–1638. [Google Scholar]
- Shi, G.N.; Zhang, C.N.; Xu, R.; Niu, J.F.; Song, H.J.; Zhang, X.Y.; Wang, W.W.; Wang, Y.M.; Li, C.; Wei, X.Q.; et al. Enhanced antitumor immunity by targeting dendritic cells with tumor cell lysate-loaded chitosan nanoparticles vaccine. Biomaterials 2016, 113, 191–202. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, W.; Liu, F.; Zheng, M.; Zheng, D.; Zhang, T. Intranasal immunization with live attenuated influenza vaccine plus chitosan as an adjuvant protects mice against homologous and heterologous virus challenge. Arch. Virol. 2012, 157, 1451–1461. [Google Scholar] [CrossRef]
- Tao, W.; Zheng, H.Q.; Fu, T. N-(2-hydroxy) propyl-3-trimethylammonium chitosan chloride: An immune-enhancing adjuvant for hepatitis E virus recombinant polypeptide vaccine in mice. Hum. Vaccine Immunother. 2017, 13, 1818–1822. [Google Scholar] [CrossRef] [Green Version]
- Farris, E.; Brown, D.; Ramer-Tait, A.; Pannier, A. Chitosan-zein nano-in-microparticles capable of mediating in vivo transgene expression following oral delivery. J. Control. Release 2017, 249, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wu, N.; Yue, H. Safety evaluation of chitosan quaternary ammonium salt microspheres as adjuvant for injection vaccine. J. Process Eng. 2016, 16, 647–653. [Google Scholar]
- Prajapati, S.; Jain, A.; Jain, A.; Jain, S. Biodegradable polymers and constructs: A novel approach in drug delivery. Eur. Polym. J. 2019, 120, 109191. [Google Scholar] [CrossRef]
- Ghitman, J.; Biru, E.; Stan, R.; Iov, H. Review of hybrid PLGA nanoparticles: Future of smart drug delivery and theranostics medicine. Mater. Des. 2020, 193, 108805. [Google Scholar] [CrossRef]
- Wu, X.Y.; Li, Y.C.; Raza, F.; Wang, X.R.; Zhang, S.L.; Rong, R.N.; Qiu, M.F.; Su, J. Red Blood Cell Membrane-Camouflaged Tedizolid Phosphate-Loaded PLGA Nanoparticles for Bacterial-Infection Therapy. Pharmaceutics 2021, 13, 99. [Google Scholar] [CrossRef]
- Cheng, S.; Nethi, S.K.; Al-Kofahi, M.; Prabha, S. Pharmacokinetic—Pharmacodynamic Modeling of Tumor Targeted Drug Delivery Using Nano-Engineered Mesenchymal Stem Cells. Pharmaceutics 2021, 13, 92. [Google Scholar] [CrossRef]
- Duskey, J.T.; Baraldi, C.; Gamberini, M.C.; Ottonelli, I.; Ros, F.D.; Tosi, G.; Forni, F.; Vandelli, M.A.; Ruozi, B. Investigating Novel Syntheses of a Series of Unique Hybrid PLGA-Chitosan Polymers for Potential Therapeutic Delivery Applications. Polymers 2020, 12, 823. [Google Scholar] [CrossRef] [Green Version]
- George, A.; Shah, P.; Shrivastav, P. Natural biodegradable polymers-based nano-formulations for drug delivery: A review. Int. J. Pharm. 2019, 561, 244–264. [Google Scholar] [CrossRef]
- Lima, A.F.; Amado, I.R.; Pires, L.R. Poly(d, l-lactide-co-glycolide) (PLGA) Nanoparticles Loaded with Proteolipid Protein (PLP)—Exploring a New Administration Route. Polymers 2020, 12, 3063. [Google Scholar] [CrossRef]
- Khan, A.; Zhou, Z.W.; He, W.M.; Gao, K.; Khan, M.W.; Faisal, R.; Muhammad, H.; Sun, M. CXCR4-Receptor-Targeted Liposomes for the Treatment of Peritoneal Fibrosis. Mol. Pharm. 2019, 16, 2728–2741. [Google Scholar]
- Luo, L.; Zheng, S.; Huang, Y.; Qin, T.; Xing, J.; Niu, Y.; Bo, R.; Liu, Z.; Huang, Y.; Hu, Y.; et al. Preparation and characterization of Chinese yam polysaccharide PLGA nanoparticles and their immunological activity. Int. J. Pharm. 2016, 511, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Raza, F.; Zafar, H.; Zhang, S.L.; Kamal, Z.; Su, J.; Yuan, W.E.; Qiu, M.M. Recent Advances in Cell Membrane-Derived Biomimetic Nanotechnology for Cancer Immunotherapy. Adv. Healthc. Mater. 2021, 10, e2002081. [Google Scholar] [CrossRef]
- Phanthipha, R.; Peerapong, P.; Prapaporn, U. A mannose-specific C-type lectin from Fenneropenaeus merguiensis exhibited antimicrobial activity to mediate shrimp innate immunity. Mol. Immunol. 2017, 92, 87–98. [Google Scholar]
- Raza, F.; Zafar, H.; Zhu, Y.; Ren, Y.; Ullah, A.; Khan, A.U.; He, X.Y.; Han, H.; Aquib, M.; Boakye-Yiadom, K.O.; et al. Review on Recent Advances in Stabilizing Peptides/Proteins upon Fabrication in Hydrogels from Biodegradable Polymers. Pharmaceutics 2018, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L. Construction and Evaluation of Dendritic Cell Targeting Vector Mediated by Mannose Receptor; Suzhou University: Suzhou, China, 2009; pp. 123–146. [Google Scholar]
- Feng, J.; Zhao, L.; Yu, Q. Receptor-mediated stimulatory effect of oligochitosan in macrophages. Biochem. Biophys. Res. Commun. 2004, 317, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Otsuka, F.; Mukai, T.; Otani, H.; Inagaki, K.; Miyoshi, T. TNF-α inhibition of BMP-2-induced osteoblast differentiation by regulating Smad signaling and Ras/Rho-MAPK pathway. Clin. Chim. Acta. 2008, 7, 890–891. [Google Scholar]
- Wi, T.I.; Byeon, Y.; Won, J.E.; Lee, J.M.; Kang, T.H.; Lee, J.-W.; Lee, Y.J.; Sood, A.K.; Han, H.D.; Park, Y.M. Selective Tumor-Specific Antigen Delivery to Dendritic Cells Using Mannose-Labeled Poly(d, l-lactide-co-glycolide) Nanoparticles for Cancer Immunotherapy. J. Biomed. Nanotechnol. 2020, 16, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.H.; Qin, F.H.; Ji, Z.H.; Fei, W.; Tan, Z.; Hu, Y.; Zheng, C.H. Mannose-Modified PLGA Nanoparticles for Sustained and Targeted Delivery in Hepatitis B Virus Immunoprophylaxis. AAPS Pharm. Sci. Tech. 2019, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.B.; Fan, J.; Song, Z.H.; Du, X.G.; Chen, Y.; Wang, J.S.; Song, G.D. Characterization and immunoenhancement activities of Eucommia ulmoides polysaccharides. Carbohydr. Polym. 2016, 136, 803–811. [Google Scholar] [CrossRef]
- Feng, H.B.; Fan, J.; Lin, L.; Liu, Y.J.; Chai, D.K.; Yang, J. Immunomodulatory Effects of Phosphorylated Radix Cyathulae officinalis Polysaccharides in Immunosuppressed Mice. Molecules 2019, 24, 4150. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.B.; Fan, J.; He, B.; Xi, T.; He, B.; Wang, X. Selenylation modification can enhance immune-enhancing activity of Chuanminshen violaceum polysaccharide. Carbohydr. Polym. 2016, 153, 302–311. [Google Scholar]
- Feng, H.B.; Du, X.G.; Liu, J.; Han, X.F.; Cao, X.H.; Zeng, X.Y. Novel polysaccharide from Radix Cyathulae officinalis Kuan can improve immune response to ovalbumin in mice. Int. J. Biol. Macromol. 2014, 65, 121–128. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, B.; Yu, X.; Zha, X.; Zhang, X.; Chen, Y. Controlled release of PEI/DNA complexes from mannose bearing chitosan microspheres as a potent delivery system to enhance immune response to HBV DNA vaccine. J. Control. Release 2007, 121, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Q.; Yang, L.; Wan, F.; Bera, H.; Cun, D.M.; Rantanen, J.; Yang, M. Quality by design thinking in the development of long-acting injectable PLGA/PLA-based microspheres for peptide and protein drug delivery. Int. J. Pharm. 2020, 585, 119441. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ban, J.; Huang, S. Influencing factors and control of drug loading and entrapment rate of PLGA microspheres. J. Guangdong Inst. Pharm. 2011, 27, 643–648. [Google Scholar]
- Ulusoy, A.; Onur, M.A. Measurement of in vitro phagocytic activity using functional groups carrying monodisperse poly(glycidyl methacrylate) microspheres in rat blood. J. Biomater. Sci. Polym. Ed. 2003, 14, 1299–1310. [Google Scholar] [CrossRef]
- Singh, M.; Kazzaz, J.; Ugozzoli, M.; Chesko, J.; O’Hagan, D.T. Charged polylactide co-glycolide microparticles as antigen delivery systems. Expert Opin. Biol. 2004, 4, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Mandal, B.; Kempf, M.; Merkle, H.P.; Walter, E. Immobilisation of gm-csf onto particulate vaccine carrier systems. Int. J. Pharm. 2004, 269, 259–265. [Google Scholar] [CrossRef]
- Kumar, M.; Bakowsky, U.; Lehr, C.M. Preparation and characterization of cationic plga nanospheres as DNA carriers. Biomaterials 2004, 25, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.N.; Mohapatra, S.S.; Kong, X.; Jena, P.K.; Bakowsky, U.; Lehr, C.M. Cationic poly(lactide-coglycolide) nanoparticles as efficient in vivo gene transfection agents. J. Nanosci. Nanotechnol. 2004, 4, 990–994. [Google Scholar] [CrossRef]
- Munier, S.; Messai, I.; Delair, T.; Verrier, B.; Ataman-Onal, Y. Cationic pla nanoparticles for DNA delivery:Comparison of three surface polycations for DNA binding, protection and transfection properties. Colloids Surf. B Biointerfaces 2005, 43, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Tsung, M.J.; Burgess, D.J. Preparation and characterization of gelatin surface modified plga microspheres. AAPS PharmSci 2001, 3, E11. [Google Scholar]
- Darrell, J.I.; Benjamin, J.R. Shaping humoral immunity to vaccines through antigen-displaying nanoparticles. Curr. Opin. Immunol. 2020, 65, 1–6. [Google Scholar]
- Li, H.; Tae-EunPark, B.; Hong, Z.; Kang, S.; Cho, C.; Choi, Y. Mannan-decorated thiolated Eudragit microspheres for targeting antigen presenting cells via nasal vaccination. Eur. J. Pharm. Sci. 2015, 80, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, G.K.; Abhinav, A.; Jamal, S.L. Innate and Adaptive Immunity: The Immune Response to Foreign Materials. Biomater. Sci. 2020, 747–776.e1. [Google Scholar] [CrossRef]
- Singh, R.; Alape, D.; de Lima, A.; Ascanio, J.; Majid, A.; Gangadharan, S.P. Regulatory T Cells in Respiratory Health and Diseases. BMC Pulm. Med. 2019, 20, 1907807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saigusa, R.; Winkels, H.; Ley, K. T cell subsets and functions in atherosclerosis. Nat. Rev. Cardiol. 2020, 17, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Vlies, A.; Simeoni, E.; Angeli, V.; Randolph, G.; Neil, C. Exploiting lymphatic transport and complement activation in nanoparticle vaccines. Nat. Biotechnol. 2007, 25, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Constantino, J.; Gomes, C.; Falcão, A.; Neves, B.M.; Cruz, M.T. Dendritic cell-based immunotherapy: A basic review and recent advances. Immunol. Res. 2017, 65, 798–810. [Google Scholar] [CrossRef]
- Christian, E.B.; Sarah, S.; Benjamin, K.; Michael, S.P.; Phillip, D.F.; Derek, N.J.H. Dendritic cells as cancer therapeutics. Semin. Cell Dev. Biol. 2019, 86, 77–88. [Google Scholar]
- Hartgers, F.; Figdor, C.; Adema, G. Towards a molecular understanding of dendritic cell immunobilogy. Immunol. Today 2000, 21, 542–545. [Google Scholar] [CrossRef] [Green Version]
- Wculek, S.K.; Cueto, F.J.; Mujal, A.M.; Melero, I.; Krummel, M.F.; Sancho, D. Dendritic cells in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2020, 20, 7–24. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and innate immunity: New perspectives on host defence. Semin Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef]
- Dowling, J.K.; Mansell, A. Toll-like receptors: The swiss army knife of immunity and vaccine development. Clin. Transl. Immunol. 2016, 20, e85. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.; de Mingo Pulido, Á.; Ruffell, B. Dendritic Cells and Their Role in Immunotherapy. Front. Immunol. 2020, 11, 924. [Google Scholar] [CrossRef]
- Keselowsky, B.G.; Lewis, J.S. Dendritic cells in the host response to implanted materials. Semin Immunol. 2017, 29, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PLGA | PVA | OVA | |
|---|---|---|---|
| 1 | 10 | 1 | 50 |
| 2 | 20 | 2 | 100 |
| 3 | 40 | 3 | 150 |
| Chitosan | Mannose-Modified Chitosan |
|---|---|
| C:39.99 | C:39.26 |
| H:6.42 | H:6.87 |
| O:45.37 | O:45.52 |
| N:7.63 | N:7.24 |
| PLGA | PVA | OVA | LE(%) | |
|---|---|---|---|---|
| 1 | 10 | 1 | 50 | 3.25 |
| 2 | 10 | 2 | 100 | 3.15 |
| 3 | 10 | 3 | 150 | 1.27 |
| 4 | 20 | 1 | 100 | 1.69 |
| 5 | 20 | 2 | 150 | 6.48 |
| 6 | 20 | 3 | 50 | 3.22 |
| 7 | 40 | 1 | 150 | 5.83 |
| 8 | 40 | 2 | 50 | 8.4 |
| 9 | 40 | 3 | 100 | 8.1 |
| K1 | 2.55 | 3.59 | 4.95 | |
| K2 | 3.79 | 6.01 | 4.31 | |
| K3 | 7.44 | 4.19 | 4.52 | |
| R | 4.88 | 2.42 | 0.43 | |
| A3B2C1 | ||||
| Group | Zeta EMF (mv) | Particle Size (μm) | Drug Loading (%) |
|---|---|---|---|
| OVA-PLGA-MPs | −14.8 ± 1.9 a | 6.5 ± 0.4 a | 7.9 ± 0.2 a |
| CS-OVA-PLGA-MPs | 31.6 ± 0.4 b | 9.0 ± 0.5 b | 6.5 ± 0.2 b |
| MAN-CS-OVA-PLGA-MPs | 30.8 ± 1.1 b | 7.9 ± 0.5 c | 6.2 ± 1.2 b |
| Group | IL-2 (pg/mL) | IL-4 (pg/mL) | IL-6 (pg/mL) | IFN-γ (pg/mL) |
|---|---|---|---|---|
| Saline | 4.295 ± 0.045 a | 11.540 ± 2.820 a | 36.835 ± 0.485 a | 27.200 ± 0.630 a |
| OVA | 4.210 ± 0.580 a | 12.735 ± 0.835 a,b | 37.320 ± 1.230 a | 28.665 ± 0.845 a,b |
| OVA-PLGA-MPs | 4.245 ± 0.315 a | 16.015 ± 2.845 a,b | 37.805 ± 1.275 a,b | 28.900 ± 0.430 a,b |
| CS-OVA-PLGA-MPs | 4.525 ± 0.165 a | 16.255 ± 1.185 b | 39.930 ± 0.750 b | 29.565 ± 0.075 a,b |
| MAN-CS-OVA-PLGA-MPs | 4.635 ± 0.045 a | 21.860 ± 3.720 c | 40.175 ± 1.975 c | 30.525 ± 1.065 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, H.; Yang, X.; Zhang, L.; Liu, Q.; Feng, Y.; Wu, D.; Liu, Y.; Yang, J. Mannose-Modified Chitosan Poly(lactic-co-glycolic acid) Microspheres Act as a Mannose Receptor-Mediated Delivery System Enhancing the Immune Response. Polymers 2021, 13, 2208. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13132208
Feng H, Yang X, Zhang L, Liu Q, Feng Y, Wu D, Liu Y, Yang J. Mannose-Modified Chitosan Poly(lactic-co-glycolic acid) Microspheres Act as a Mannose Receptor-Mediated Delivery System Enhancing the Immune Response. Polymers. 2021; 13(13):2208. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13132208
Chicago/Turabian StyleFeng, Haibo, Xiaonong Yang, Linzi Zhang, Qianqian Liu, Yangyang Feng, Daiyan Wu, Yunjie Liu, and Jie Yang. 2021. "Mannose-Modified Chitosan Poly(lactic-co-glycolic acid) Microspheres Act as a Mannose Receptor-Mediated Delivery System Enhancing the Immune Response" Polymers 13, no. 13: 2208. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13132208