Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis Peruviana L. Seedlings Exposed to Different Short-Term Waterlogging Periods and Fusarium Wilt Infection


Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Inoculation of the Pathogen
2.2. Waterlogging Treatments
2.3. Analysis of Vascular Wilt Severity
2.4. Stomatal Conductance and Leaf Water Potential
2.5. Chlorophyll Fluorescence Parameters
2.6. Diameter of the Stem and Leaf Area
2.7. Leaf Photosynthetic Pigments
2.8. Proline and Malondialdehyde Content
2.9. Experimental Design and Data Analysis
3. Results and Discussion
3.1. Vascular Wilt Severity
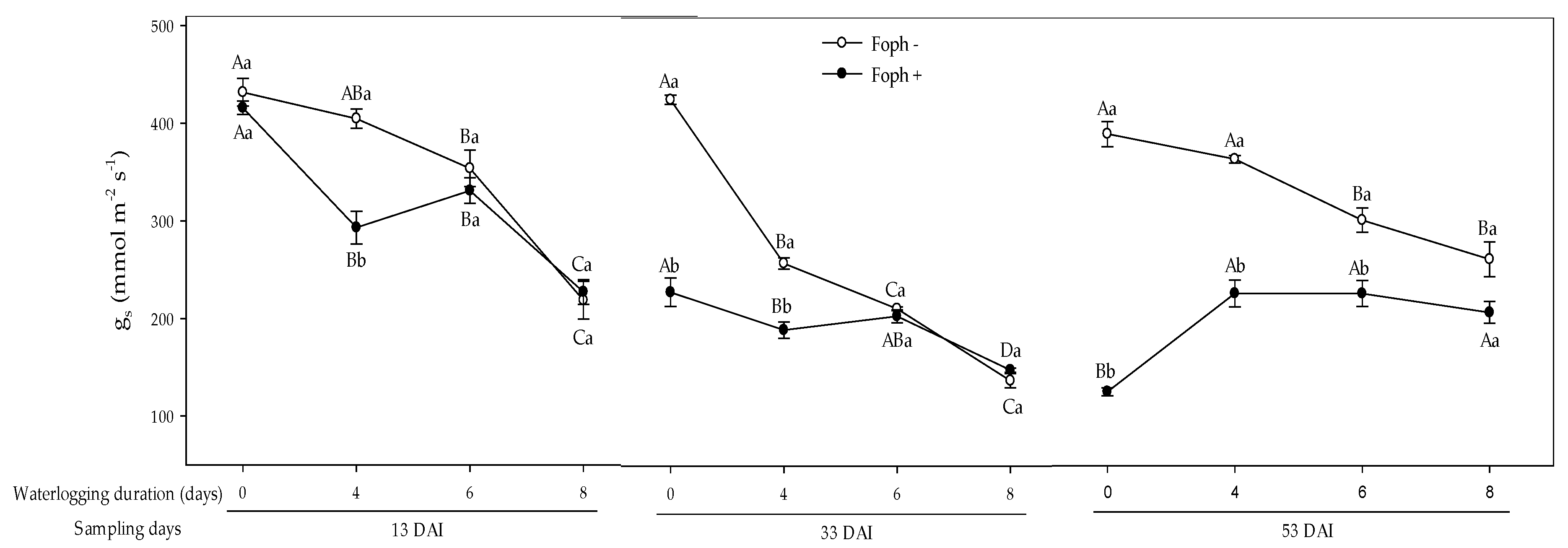
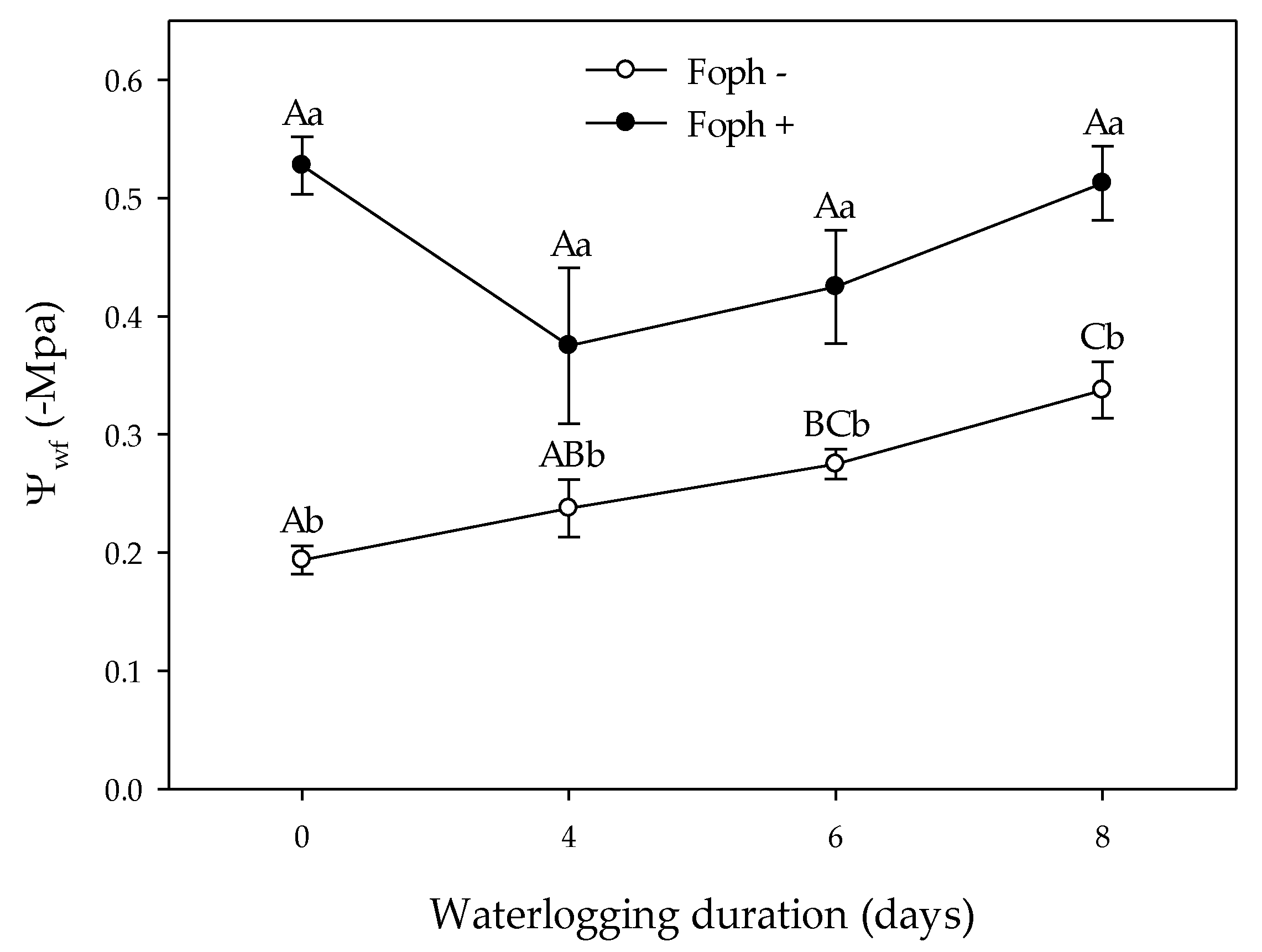
3.2. Stomatal Conductance (gs) and Leaf Water Potential (Ψwf)
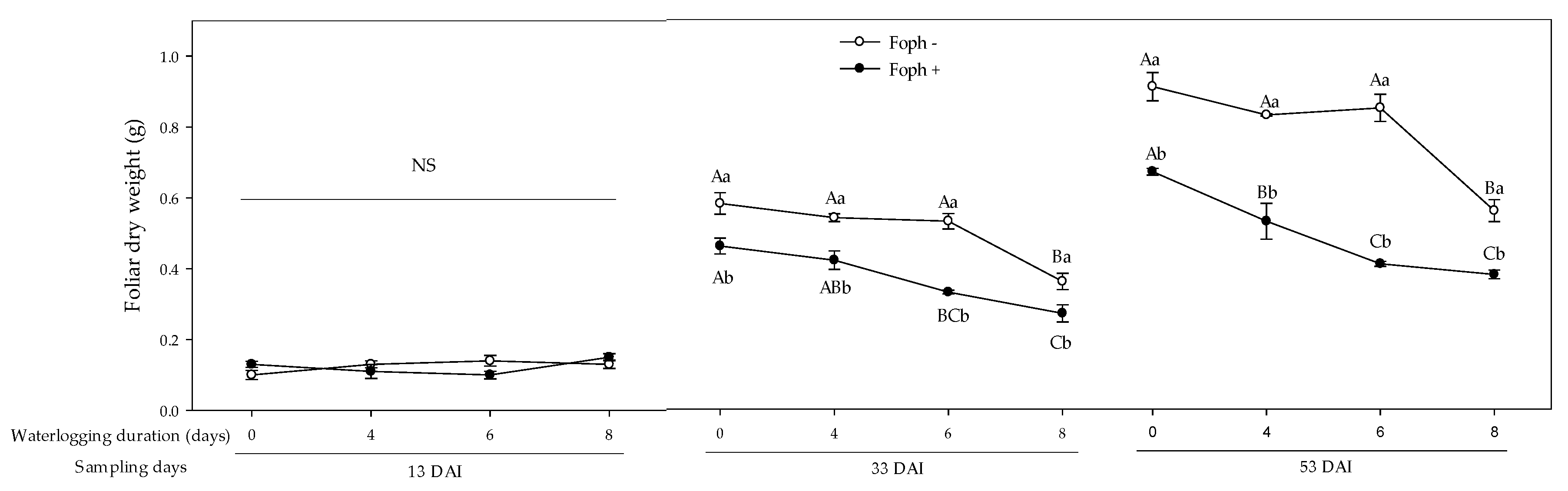
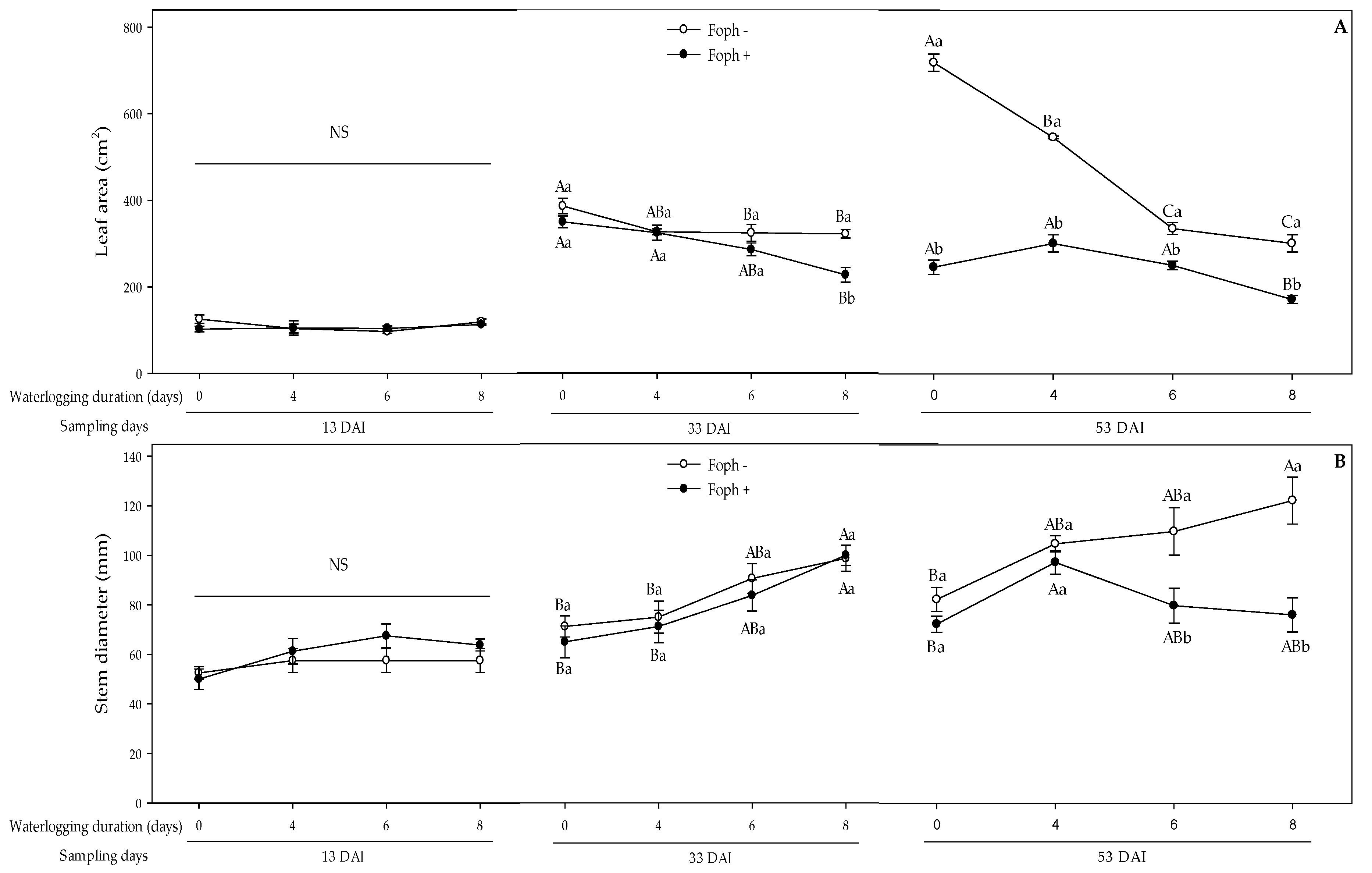
3.3. Growth Response
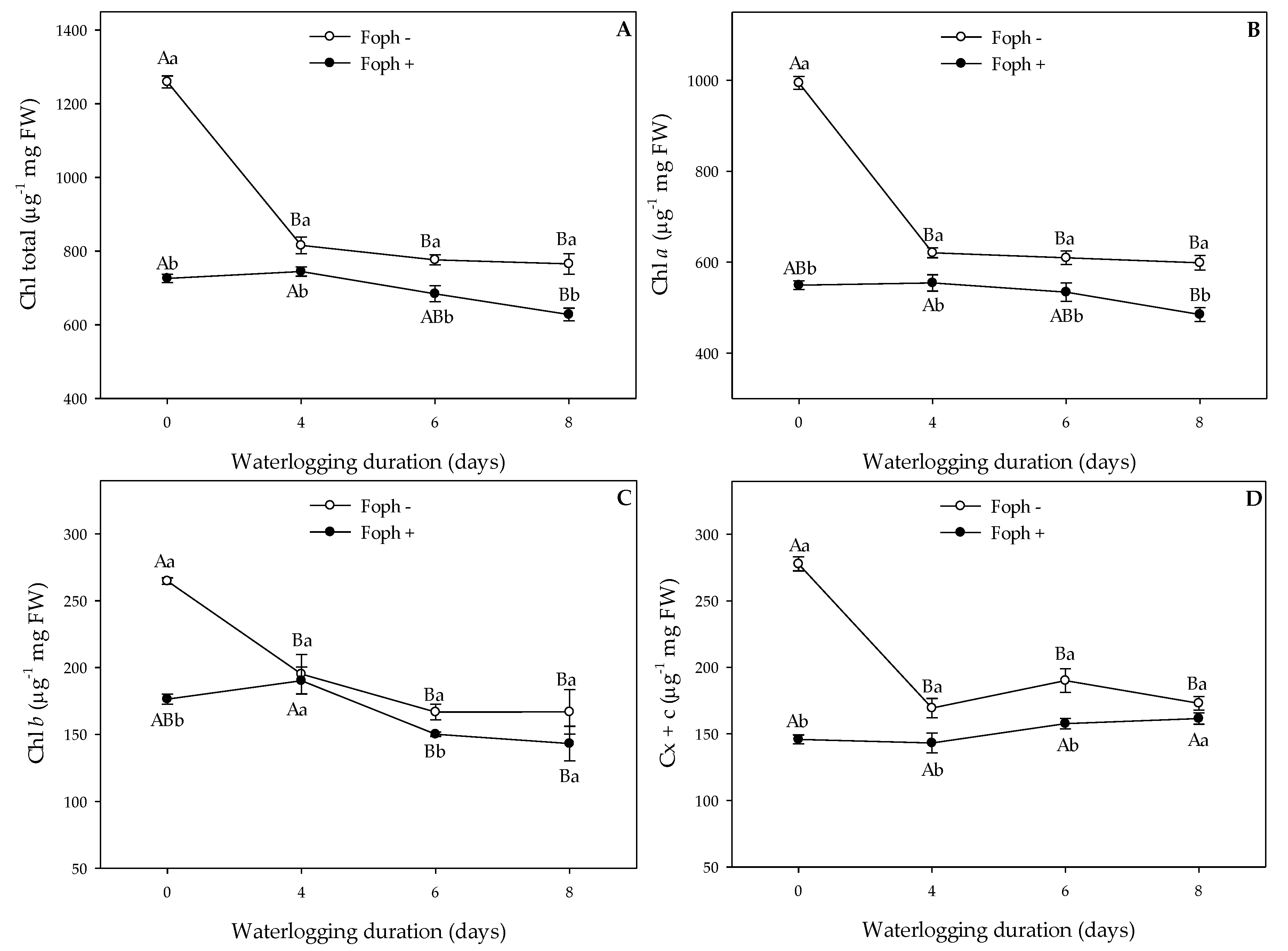
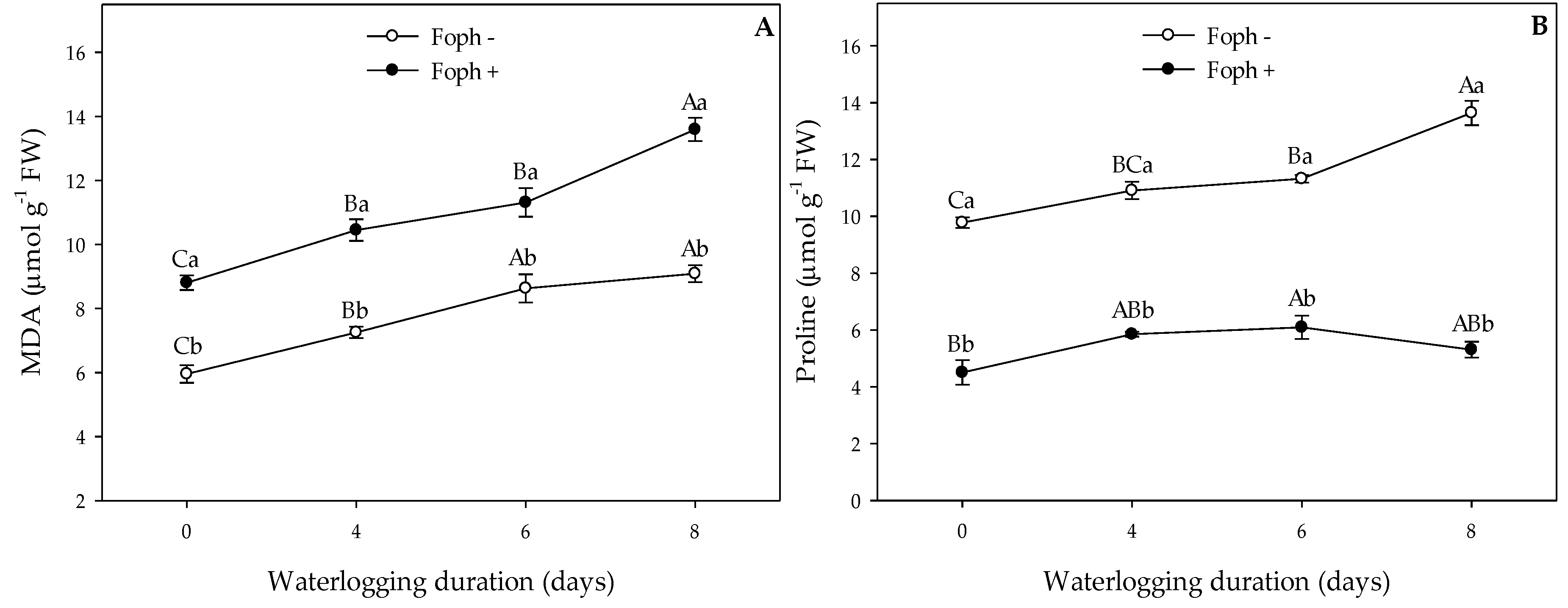
3.4. Biochemical Tests on Leaves (Malondialdehyde Content, Photosynthetic Pigments and Proline)
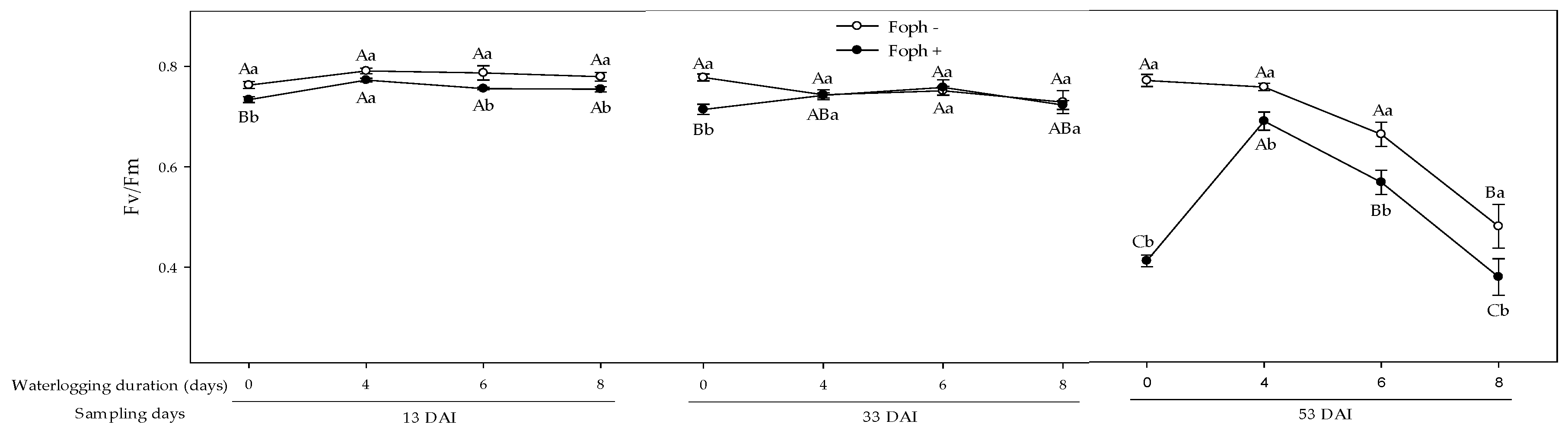
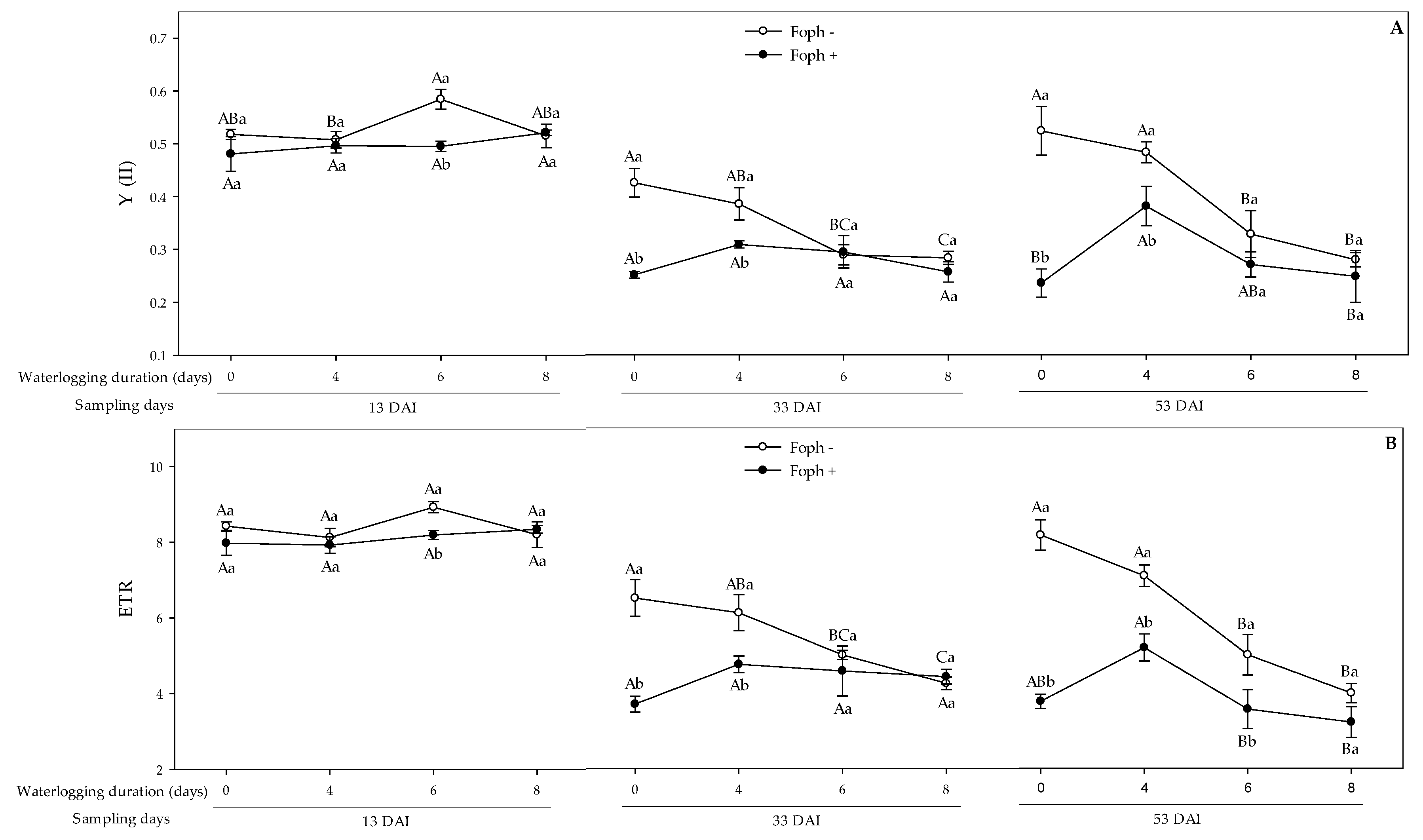
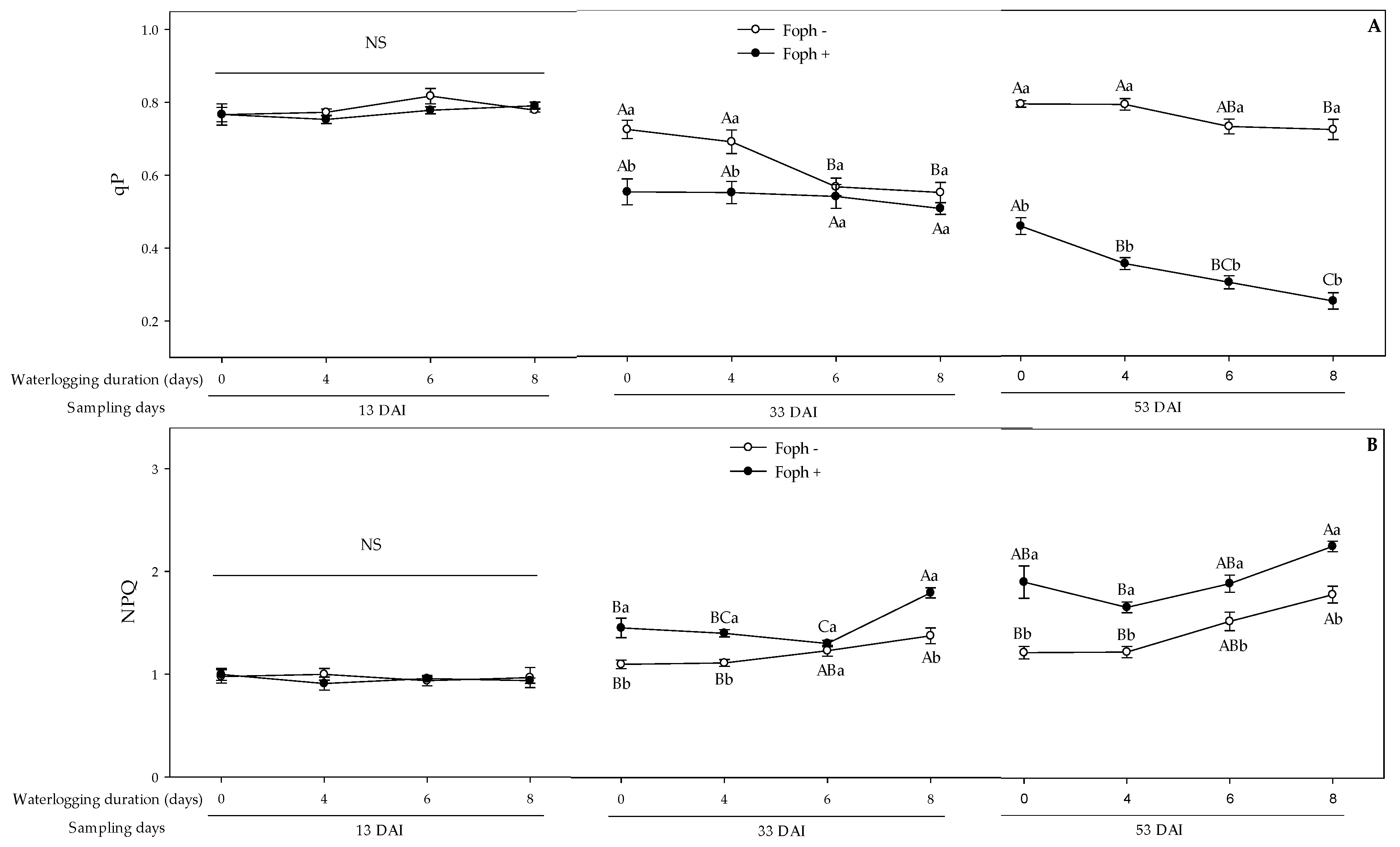
3.5. Chlorophyll Fluorescence Parameters
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fischer, G.; Almanza-Merchán, P.J.; Miranda, D. Importancia y cultivo de la uchuva (Physalis peruviana L.). Rev. Bras. Frutic. 2014, 36, 1–15. [Google Scholar] [CrossRef]
- Ramadan, M.F. Bioactive phytochemicals, nutritional value, and functional properties of cape gooseberry (Physalis peruviana): An overview. Food Res. Int. 2011, 44, 1830–1836. [Google Scholar] [CrossRef]
- Agronet. Available online: http://www.agronet.gov.co/estadistica/Paginas/default.aspx (accessed on 1 October 2019).
- Osorio-Guarín, J.A.; Enciso-Rodríguez, F.E.; González, C.; Fernández-Pozo, N.; Mueller, L.A.; Barrero, L.S. Association analysis for disease resistance to Fusarium oxysporum in cape gooseberry (Physalis peruviana L.). BMC Genomics 2016, 17, 248. [Google Scholar] [CrossRef]
- Aldana, F.; García, P.N.; Fischer, G. Effect of waterlogging stress on the growth, development and symptomatology of cape gooseberry (Physalis peruviana L.) plants. Rev. Acad. Colomb. Cienc. Exactas. Fis. Nat. 2014, 38, 393–400. [Google Scholar] [CrossRef]
- Villarreal-Navarrete, A.; Fischer, G.; Melgarejo, L.M.; Correa, G.; Hoyos-Carvajal, L. Growth response of the cape gooseberry (Physalis peruviana L.) to waterlogging stress and Fusarium oxysporum infection. Acta Hortic. 2017, 1178, 161–168. [Google Scholar] [CrossRef]
- Cotes, A.M.; Jiménez, P.; Rodríguez, M.X.; Díaz, A.; Zapata, J.; Gómez, M.; Grijalba, E.; Villamizar, L.; González, C.; Smith, A.; et al. Estrategias de control biológico de Fusarium Oxysporum en el cultivo de la Uchuva (Physalis peruviana L.); Corporación colombiana de investigación agropecuaria (Corpoica): Bogotá, Colombia, 2012; p. 82. [Google Scholar]
- Simbaqueba, J.; Catanzariti, A.M.; González, C.; Jones, D.A. Evidence for horizontal gene transfer and separation of effector recognition from effector function revealed by analysis of effector genes shared between cape gooseberry-and tomato-infecting formae speciales of Fusarium oxysporum. Mol. Plant Pathol. 2018, 19, 2302–2318. [Google Scholar] [CrossRef]
- Flórez-Velasco, N.; Balaguera-López, H.E.; Restrepo-Díaz, H. Effects of foliar urea application on lulo (Solanum quitoense cv. septentrionale) plants grown under different waterlogging and nitrogen conditions. Sci. Hortic. 2015, 186, 154–162. [Google Scholar] [CrossRef]
- Betancourt-Osorio, J.; Sánchez-Canro, D.; Restrepo-Díaz, H. Effect of nitrogen nutritional statuses and waterlogging conditions on growth parameters, nitrogen use efficiency and chlorophyll fluorescence in tamarillo Seedlings. Not. Bot. Horti Agrobot. 2016, 44, 375–381. [Google Scholar] [CrossRef]
- Wu, H.S.; Raza, W.; Fan, J.Q.; Sun, Y.G.; Bao, W.; Liu, D.Y.; Miao, W.G. Antibiotic effect of exogenously applied salicylic acid on in vitro soilborne pathogen. Fusarium oxysporum f. sp. niveum. Chemosphere 2008, 74, 45–50. [Google Scholar] [CrossRef]
- Jiménez-Fernández, D.; Montes-Borrego, M.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M.; Landa, B.B. Identification and quantification of Fusarium oxysporum in planta and soil by means of an improved specific and quantitative PCR assay. Appl. Soil Ecol. 2010, 46, 372–382. [Google Scholar] [CrossRef]
- Portal, N.; Soler, A.; Alphonsine, P.A.M.; Borras-Hidalgo, O.; Portieles, R.; Peña-Rodriguez, L.M.; Walton, J.D. Nonspecific toxins as components of a host-specific culture filtrate from Fusarium oxysporum f. sp. cubense race 1. Plant Pathol. 2018, 67, 467–476. [Google Scholar] [CrossRef]
- Lorenzini, G.; Guidi, L.; Nali, C.; Ciompi, S.; Soldatini, G.F. Photosynthetic response of tomato plants to vascular wilt diseases. Plant Sci. 1997, 124, 143–152. [Google Scholar] [CrossRef]
- Nogués, S.; Cotxarrera, L.; Alegre, L.; Trillas, M.I. Limitations to photosynthesis in tomato leaves induced by Fusarium wilt. N. Phytol. 2002, 154, 461–470. [Google Scholar] [CrossRef]
- Gawehns, F.; Ma, L.; Bruning, O.; Houterman, P.M.; Boeren, S.; Cornelissen, B.J.; Takken, F.L. The effector repertoire of Fusarium oxysporum determines the tomato xylem proteome composition following infection. Front. Plant Sci. 2015, 6, 967. [Google Scholar] [CrossRef]
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, M.; Li, Y.; Gu, Z.; Ling, N.; Shen, Q.; Guo, S. Wilted cucumber plants infected by Fusarium oxysporum f. sp. cucumerinum do not suffer from water shortage. Ann. Bot. 2017, 120, 427–436. [Google Scholar] [CrossRef]
- Ghaemi, A.; Rahimi, A.; Banihashemi, Z. Effects of Water Stress and Fusarium oxysporum f. sp. lycoperseci on Growth (leaf area, plant height, shoot dry matter) and Shoot Nitrogen Content of Tomatoes Under Greenhouse Conditions. Iran Agric. Res. 2010, 29, 51–62. [Google Scholar] [CrossRef]
- Nosir, W.S. Trichoderma harzianum as a Growth Promoter and Bio-Control Agent against Fusarium oxysporum f. sp. tuberosi. Adv. Crop. Sci. Tech. 2016, 4, 1–7. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, M.; Sun, Y.; Gu, Z.; Wang, R.; Saydin, A.; Guo, S. Nitrate increased cucumber tolerance to fusarium wilt by regulating fungal toxin production and distribution. Toxins 2017, 9, 100. [Google Scholar] [CrossRef]
- Wu, X.; Tang, Y.; Li, C.; Wu, C.; Huang, G. Chlorophyll fluorescence and yield responses of winter wheat to waterlogging at different growth stages. Plant Prod. Sci. 2015, 18, 284–294. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Effects of waterlogging on leaf mesophyll cell ultrastructure and photosynthetic characteristics of summer maize. PLoS ONE 2016, 11, e0161424. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Li, Y. Management of flooding effects on growth of vegetable and selected field crops. HortTechnology 2003, 13, 610–616. [Google Scholar] [CrossRef]
- Kallestad, J.C.; Sammis, T.W.; Mexala, J.G.; Gutschick, V. The impact of prolonged flood-irrigation on leaf gas exchange in mature pecans in an orchard setting. Int. J. Plant Prod. 2012, 1, 163–178. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Screening for barley waterlogging tolerance in nordic barley cultivars (Hordeum vulgare L.) using chlorophyll fluorescence on hydroponically-grown plants. Agronomy 2013, 3, 376–390. [Google Scholar] [CrossRef]
- He, L.; Yu, L.; Li, B.; Du, N.; Guo, S. The effect of exogenous calcium on cucumber fruit quality, photosynthesis, chlorophyll fluorescence, and fast chlorophyll fluorescence during the fruiting period under hypoxic stress. BMC Plant Biol. 2018, 18, 180. [Google Scholar] [CrossRef]
- Casierra-Posada, F.; Cutler, J. Photosystem II fluorescence and growth in cabbage plants (Brassica oleracea var. capitata) grown under waterlogging stress. Rev. U.D.C.A Actual. Divulg. Cient. 2017, 20, 321–328. [Google Scholar] [CrossRef]
- Yanar, Y.; Lipps, P.E.; Deep, I.W. Effect of soil saturation duration and soil water content on root rot of maize caused by Pythium arrhenomanes. Plant Dis. 1997, 81, 475–480. [Google Scholar] [CrossRef]
- Kirkpatrick, M.T.; Rothrock, C.S.; Rupe, J.C.; Gbur, E.E. The effect of Pythium ultimum and soil flooding on two soybean cultivars. Plant Dis. 2006, 90, 597–602. [Google Scholar] [CrossRef]
- Newcombe, M. Some effects of water and anaerobic conditions on Fusarium oxysporum f. cubense in soil. Trans. Br. Mycol. Soc. 1960, 43, 51–59. [Google Scholar] [CrossRef]
- Jain, R.; Singh, S.P.; Singh, A.; Singh, S.; Chandra, A.; Solomon, S. Response of foliar application of nitrogen compounds on sugarcane grown under waterlogging stress. Sugar Tech. 2015, 18, 433–436. [Google Scholar] [CrossRef]
- Rao, G.S.; Reddy, N.N.R.; Surekha, C. Induction of plant systemic resistance in Legumes Cajanus cajan, Vigna radiata, Vigna mungo against plant pathogens Fusarium oxysporum and Alternaria alternata–a Trichoderma viride mediated reprogramming of plant defense mechanism. Int. J. Recent Sci. Res. 2015, 6, 4270–4280. [Google Scholar]
- Jain, S.; Vaishnav, A.; Kumari, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. Chitinolytic Bacillus-mediated induction of jasmonic acid and defense-related proteins in soybean (Glycine max L. Merrill) plant against Rhizoctonia solani and Fusarium oxysporum. J. Plant Growth Regul. 2016, 36, 200–214. [Google Scholar] [CrossRef]
- Rasheed, R.; Iqbal, M.; Ashraf, M.A.; Hussain, I.; Shafiq, F.; Yousaf, A.; Zaheer, A. Glycine betaine counteracts the inhibitory effects of waterlogging on growth, photosynthetic pigments, oxidative defence system, nutrient composition, and fruit quality in tomato. J. Hortic. Sci. Biotechnol. 2018, 93, 385–391. [Google Scholar] [CrossRef]
- Fortunato, A.A.; Rodrigues, F.Á.; do Nascimento, K.J.T. Physiological and biochemical aspects of the resistance of banana plants to Fusarium wilt potentiated by silicon. Phytopathology 2012, 102, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Dar, M.I.; Naikoo, M.I.; Rehman, F.; Naushin, F.; Khan, F.A. Proline Accumulation in Plants: Roles in Stress Tolerance and Plant Development. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Iqbal, N., Nazar, R., A. Khan, N., Eds.; Springer: New Delhi, India, 2016; pp. 155–166. ISBN 978-81-322-2615-4. [Google Scholar]
- Tuo, X.Q.; Li, S.; Wu, Q.S.; Zou, Y.N. Alleviation of waterlogged stress in peach seedlings inoculated with Funneliformis mosseae: Changes in chlorophyll and proline metabolism. Sci. Hortic. 2015, 197, 130–134. [Google Scholar] [CrossRef]
- 1Hao, Z.; Christie, P.; Qin, L.; Wang, C.; Li, X. Control of fusarium wilt of cucumber seedlings by inoculation with an arbuscular mycorrhical fungus. J. Plant Nutr. 2005, 28, 1961–1974. [Google Scholar] [CrossRef]
- Das, B.; Padhiary, A.K.; Behera, S.; Mishra, S.P.; Jena, M.; Swain, S.; Rout, S. Biochemical Changes in Some Rice Varieties in Response to Waterlogged and Submerged Conditions. Int. J. Pure Appl. Biosci. 2017, 5, 972–978. [Google Scholar] [CrossRef]
- Bansal, R.; Srivastava, J.P. Effect of waterlogging on photosynthetic and biochemical parameters in pigeonpea. Russ. J. Plant Physiol. 2015, 62, 322–327. [Google Scholar] [CrossRef]
- Verma, K.K.; Singh, M.; Gupta, R.K.; Verma, C.L. Photosynthetic gas exchange, chlorophyll fluorescence, antioxidant enzymes, and growth responses of Jatropha curcas during soil flooding. Turk. J. Bot. 2014, 38, 130–140. [Google Scholar] [CrossRef]
- Rajeswari, P. Management of Fusarium wilt caused by Fusarium oxysporum by Trichoderma Species and Pseudomonas fluorescens on Arachis hypogaea (Groundnut). J. Biol. Control 2014, 28, 133–136. [Google Scholar]
- Rejeb, I.; Pastor, V.; Mauch-Mani, B. Plant responses to simultaneous biotic and abiotic stress: Molecular mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef]
- Dikilitas, M.; Karakas, S.; Hashem, A.; Abd_Allah, E.F.; Ahmad, P. Oxidative stress and plant responses to pathogens under drought conditions. In Water Stress and Crop Plants: A Sustainable Approach, 1st ed.; Parvaiz, A., Ed.; John Wiley Sons, Ltd.: Chichester, UK, 2016; Volume 1, pp. 102–123. [Google Scholar]
- Deepthi, V.; Raksha, B.; Pooja, S.; Subashkumar, R.; Vivekanandhan, G.; Siva, R.; Babu, S. Differential response of cultivated rice to pathogen challenge and abiotic stresses with reference to cationic peroxidase. Arch. Phytopathol. Plant Prot. 2014, 47, 1390–1399. [Google Scholar] [CrossRef]
- Coakley, S.M.; Scherm, H.; Chakraborty, S. Climate change and plant disease management. Annu. Rev. Phytopathol. 1999, 37, 399–426. [Google Scholar] [CrossRef]
- Petzoldt, C.; Seaman, A. Climate change effects on insects and pathogens. Clim. Chang. Agric Promot. Pract. Profitab. Responses 2006, 3, 6–16. [Google Scholar]
- Sanogo, S.; El-Sebai, O.I.; Sanderson, R. Severity of Verticillium wilt, plant growth, and spectral reflectance indices of chile pepper under periodic flooding and no-flooding conditions. HortScience 2008, 43, 414–419. [Google Scholar] [CrossRef]
- Keatinge, J.D.H.; Lin, L.J.; Ebert, A.W.; Chen, W.Y.; Hughes, J.A.; Luther, G.C.; Ravishankar, M. Overcoming biotic and abiotic stresses in the Solanaceae through grafting: Current status and future perspectives. Biol. Agric. Hortic. 2014, 30, 272–287. [Google Scholar] [CrossRef]
- Ocampo, O. El cambio climático y su impacto en el agro. Rev. Ing. 2011, 33, 115–123. [Google Scholar]
- Ramirez-Villegas, J.; Salazar, M.; Jarvis, A.; Navarro-Racines, C.E. A way forward on adaptation to climate change in Colombian agriculture: Perspectives towards 2050. Clim. Chang. 2012, 115, 611–628. [Google Scholar] [CrossRef]
- Rekah, Y.; Shtienberg, D.; Katan, J. Disease development following infection of tomato and basil foliage by airborne conidia of the soilborne pathogens Fusarium oxysporum f. sp. radicis-lycopersici and F. oxysporum f. sp. basilici. Phytopathology 2000, 90, 1322–1329. [Google Scholar] [CrossRef]
- Park, D. Isolation of Fusarium oxysporum from soils. Trans. Br. Mycol. Soc. 1961, 44, 119–122. [Google Scholar] [CrossRef]
- Namiki, F.; Shiomi, T.; Kayamura, T.; Tsuge, T. Characterization of the formae speciales of Fusarium oxysporum causing wilts of cucurbits by DNA fingerprinting with nuclear repetitive DNA sequences. Appl. Environ. Microbiol. 1994, 60, 2684–2691. [Google Scholar]
- Hainaut, P.; Remacle, T.; Decamps, C.; Lambert, R.; Sadok, W. Higher forage yields under temperate drought explained by lower transpiration rates under increasing evaporative demand. Eur. J. Agron. 2016, 72, 91–98. [Google Scholar] [CrossRef]
- Moreno, C. Interactions between Bacillus amyloliquefaciens Bs006, Fusarium oxysporun Map5 and Cape gooseberry (Physalis peruviana). Ph.D. Thesis, Universidad Nacioanl de Colombia, Bogotá, Colombia, 2017. [Google Scholar]
- Townsend, G.R.; Heuberger, J.W. Methods for estimating losses caused by diseases in fungicide experiments. Plant Dis. Report. 1943, 27, 340–343. [Google Scholar]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; Wiley-Interscience: New York, NY, USA, 1990; p. 532. [Google Scholar]
- Alves, D.P.; Tomaz, R.S.; Laurindo, B.S.; Laurindo, R.D.F.; Cruz, C.D.; Nick, C.; Silva, D.J.H.D. Artificial neural network for prediction of the area under the disease progress curve of tomato late blight. Sci. Agric. 2017, 74, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, J.; Singh, S.; Ahmed, S.; Chaudhary, R.G.; Sarker, A. Vascular wilt disease of lentil: A review. J. Lentil Res. 2010, 4, 1–14. [Google Scholar]
- Li, Y.P.; You, M.P.; Colmer, T.D.; Barbetti, M.J. Effect of timing and duration of soil saturation on soilborne Pythium diseases of common bean (Phaseolus vulgaris). Plant Dis. 2015, 99, 112–118. [Google Scholar] [CrossRef]
- Chaudhary, S.; Kusakabe, A.; Melgar, J.C. Phytophthora infection in flooded citrus trees reduces root hydraulic conductance more than under non-flooded condition. Sci. Hortic. 2016, 202, 107–110. [Google Scholar] [CrossRef]
- Moslemi, A.; Ades, P.K.; Groom, T.; Nicolas, M.E.; Taylor, P.W. Influence of waterlogging on growth of pyrethrum plants infected by the crown and root rot pathogens, Fusarium oxysporum, Fusarium avenaceum and Paraphoma vinacea. Australas. Plant Pathol. 2018, 47, 205–213. [Google Scholar] [CrossRef]
- Rasheed-Depardieu, C.; Parelle, J.; Tatin-Froux, F.; Parent, C.; Capelli, N. Short-term response to waterlogging in Quercus petraea and Quercus robur. A study of the root hydraulic responses and the transcriptional pattern of aquaporins. Plant Physiol. Biochem. 2015, 97, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Jurczyk, B.; Pociecha, E.; Janowiak, F.; Kabała, D.; Rapacz, M. Variation in waterlogging-triggered stomatal behavior contributes to changes in the cold acclimation process in prehardened Lolium perenne and Festuca pratensis. Plant Physiol. Biochem. 2016, 109, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Olmo-Vega, A.; García-Sánchez, F.; Simón-Grao, S.; Simón, I.; Lidón, V.; Nieves, M.; Martínez-Nicolás, J.J. Physiological responses of three pomegranate cultivars under flooded conditions. Sci. Hortic. 2017, 224, 171–179. [Google Scholar] [CrossRef]
- Herrera, A. Responses to flooding of plant water relations and leaf gas exchange in tropical tolerant trees of a black-water wetland. Front. Plant Sci. 2013, 4, 106. [Google Scholar] [CrossRef]
- Castonguay, Y.; Nadeau, P.; Simard, R.R. Effects of flooding on carbohydrate and ABA levels in roots and shoots of alfalfa. Plant Cell Environ. 1993, 16, 695–702. [Google Scholar] [CrossRef]
- Striker, G.G.; Colmer, T.D. Flooding tolerance of forage legumes. J. Exp. Bot. 2016, 68, 1851–1872. [Google Scholar] [CrossRef]
- Wang, M.; Sun, Y.; Sun, G.; Liu, X.; Zhai, L.; Shen, Q.; Guo, S. Water balance altered in cucumber plants infected with Fusarium oxysporum f. sp. cucumerinum. Sci. Rep. 2015, 5, 7722. [Google Scholar] [CrossRef]
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.; Guo, S. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol. Biochem. 2012, 60, 171–179. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.W.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R.L. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Abdel-Motaal, F.; El-Sayed, M.; Jogaiah, S.; Shigyo, M.; Ito, S.I.; Tran, L.S.P. Dissection of Trichoderma longibrachiatum-induced defense in onion (Allium cepa L.) against Fusarium oxysporum f. sp. cepa by target metabolite profiling. Plant Sci. 2016, 246, 128–138. [Google Scholar] [CrossRef]
- Alwathnani, H.A.; Perveen, K. Biological control of Fusarium wilt of tomato by antagonist fungi and cyanobacteria. Afr. J. Biotechnol. 2012, 11, 1100–1105. [Google Scholar] [CrossRef]
- Sánchez-Reinoso, A.D.; Ligarreto-Moreno, G.A.; Restrepo-Díaz, H. Physiological and biochemical expressions of a determinated growth common bean genotype (Phaseolus vulgaris L.) to water deficit stress periods. J. Anim. Plant Sci. 2018, 28, 119–127. [Google Scholar]
- Pérez-Jiménez, M.; Hernández-Munuera, M.; Piñero Zapata, M.C.; López-Ortega, G.; del Amor, F.M. Two minuses can make a plus: Waterlogging and elevated CO2 interactions in sweet cherry (Prunus avium) cultivars. Physiol. Plant. 2017, 161, 257–272. [Google Scholar] [CrossRef]
- Chanu, W.S.; Sarangthem, K. Changes in proline accumulation, amino acid, sugar and chlorophyll content in leaf and culm of Phourel-amubi, a rice cultivar of Manipur in response to flash flood. Indian J. Plant Physiol. 2015, 20, 10–13. [Google Scholar] [CrossRef]
- Chaerle, L.; Hagenbeek, D.; De Bruyne, E.; Van Der Straeten, D. Chlorophyll fluorescence imaging for disease-resistance screening of sugar beet. Plant Cell Tissue Organ. Cult. 2007, 91, 97–106. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant 2016, 38, 102. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Khan, N.A. Ethylene reverses photosynthetic inhibition by nickel and zinc in mustard through changes in PSII activity, photosynthetic nitrogen use efficiency, and antioxidant metabolism. Protoplasma 2014, 251, 1007–1019. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, P.; Yang, W.; Sui, X.; Li, X.; Li, W.; Xu, N. Effects of flooding stress on the photosynthetic apparatus of leaves of two Physocarpus cultivars. J. Forest Res. 2017, 1–11. [Google Scholar] [CrossRef]
- Bhar, A.; Gupta, S.; Chatterjee, M.; Sen, S.; Das, S. Differential expressions of photosynthetic genes provide clues to the resistance mechanism during Fusarium oxysporum f. sp. ciceri race 1 (Foc1) infection in chickpea (Cicer arietinum L.). Eur. J. Plant Pathol. 2017, 148, 533–549. [Google Scholar] [CrossRef]
- Dong, X.; Wang, M.; Ling, N.; Shen, Q.; Guo, S. Potential role of photosynthesis-related factors in banana metabolism and defense against Fusarium oxysporum f. sp. cubense. Environ. Exp. Bot. 2016, 129, 4–12. [Google Scholar] [CrossRef]
- Sofo, A. Drought stress tolerance and photoprotection in two varieties of olive tree. Acta Agric. Scand. B Soil Plant Sci. 2011, 61, 711–720. [Google Scholar] [CrossRef]
- Reeksting, B.J.; Taylor, N.J.; Van den Berg, N. Flooding and Phytophthora cinnamomi: Effects on photosynthesis and chlorophyll fluorescence in shoots of non-grafted Persea americana (Mill.) rootstocks differing in tolerance to Phytophthora root rot. South Afr. J. Bot. 2014, 95, 40–53. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, T.; Feng, B.; Zhang, C.; Peng, S.; Zhang, X.; Tao, L. Non-photochemical quenching plays a key role in light acclimation of rice plants differing in leaf color. Front. Plant Sci. 2017, 7, 1968. [Google Scholar] [CrossRef]
- Zhou, J.; Qi, A.G.; Zhang, Y.C.; Wan, S.W.; Qin, P. Adventitious root growth and relative physiological responses to waterlogging in the seedlings of seashore mallow (Kosteletzkya virginica), a biodiesel plant. Aust. J. Crop Sci. 2012, 6, 73–80. [Google Scholar]
- Pshibytko, N.L.; Zenevich, L.A.; Kabashnikova, L.F. Changes in the photosynthetic apparatus during Fusarium wilt of tomato. Rus. J. Plant Physiol. 2006, 53, 25–31. [Google Scholar] [CrossRef]
- Nali, C.; Lorenzini, G.; Guidi, L. Ecophysiology of Susceptible and Resistant Tomato Plants Inoculated with Fusarium oxysporum f. sp. lycopersici. Acta Hortic. 2011, 914, 419–422. [Google Scholar] [CrossRef]
- Barczak-Brzyżek, A.K.; Kiełkiewicz, M.; Gawroński, P.; Kot, K.; Filipecki, M.; Karpińska, B. Cross-talk between high light stress and plant defence to the two-spotted spider mite in Arabidopsis thaliana. Exp. Appl. Acarol. 2017, 73, 177–189. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbr. | Source Variation | |||
|---|---|---|---|---|
| Foph | Waterlogging | Foph × Waterlogging | ||
| Chlorophyll a | Chl a | ***z | *** | *** |
| Chlorophyll b | Chl b | *** | *** | ** |
| Total chlorophyll | Chl total | *** | *** | *** |
| Carotenoids | Cx + c | *** | *** | *** |
| Malondialdehyde | MDA | *** | *** | * |
| Proline | *** | *** | *** |
| Source Variation | df | gs | Ψwf | SD | LA | FDW | Fv/Fm | Y(II) | qP | NPQ | ETR |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Foph | 1 | <0.001 | <0.001 | 0.0011 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Waterlogging (W) | 3 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Foph*W | 3 | <0.001 | 0.0221 | 0.3399 | <0.001 | <0.01 | <0.001 | <0.001 | 0.2914 | 0.408 | <0.001 |
| Time | 2 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Time*Foph | 2 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.0051 | <0.001 | <0.001 | <0.001 |
| Time*W | 6 | <0.001 | <0.001 | 0.0163 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Time*W*Foph | 6 | <0.001 | 0.3034 | <0.05 | <0.001 | <0.05 | <0.001 | <0.01 | <0.01 | 0.149 | <0.01 |
| C.V. (%) 1 | 8.78 | 18.53 | 12.7 | 8.42 | 10.11 | 3.69 | 13.1 | 9.56 | 7.2 | 10.98 |
| Treatment | AUDPC | Disease Index |
|---|---|---|
| F. oxysporum | 45.25 c 1 | 2.5 b |
| 4 days + Foph | 41.25 c | 2.75 ab |
| 6 days + Foph | 55.25 b | 3.75 ab |
| 8 days + Foph | 64.25 a | 4.25 a |
| Significance | *** 2 | ** |
| CV (%) 3 | 8.29 | 26.23 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chávez-Arias, C.C.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis Peruviana L. Seedlings Exposed to Different Short-Term Waterlogging Periods and Fusarium Wilt Infection. Agronomy 2019, 9, 213. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9050213
Chávez-Arias CC, Gómez-Caro S, Restrepo-Díaz H. Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis Peruviana L. Seedlings Exposed to Different Short-Term Waterlogging Periods and Fusarium Wilt Infection. Agronomy. 2019; 9(5):213. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9050213
Chicago/Turabian StyleChávez-Arias, Cristhian C., Sandra Gómez-Caro, and Hermann Restrepo-Díaz. 2019. "Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis Peruviana L. Seedlings Exposed to Different Short-Term Waterlogging Periods and Fusarium Wilt Infection" Agronomy 9, no. 5: 213. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9050213