Chlorophyll α Fluorescence Parameters as an Indicator to Identify Drought Susceptibility in Common Bush Bean


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Water Deficit Treatment
2.3. Photosynthetic Pigments and Chlorophyll Fluorescence Determination
2.4. Experimental Design and Data Analysis
3. Results
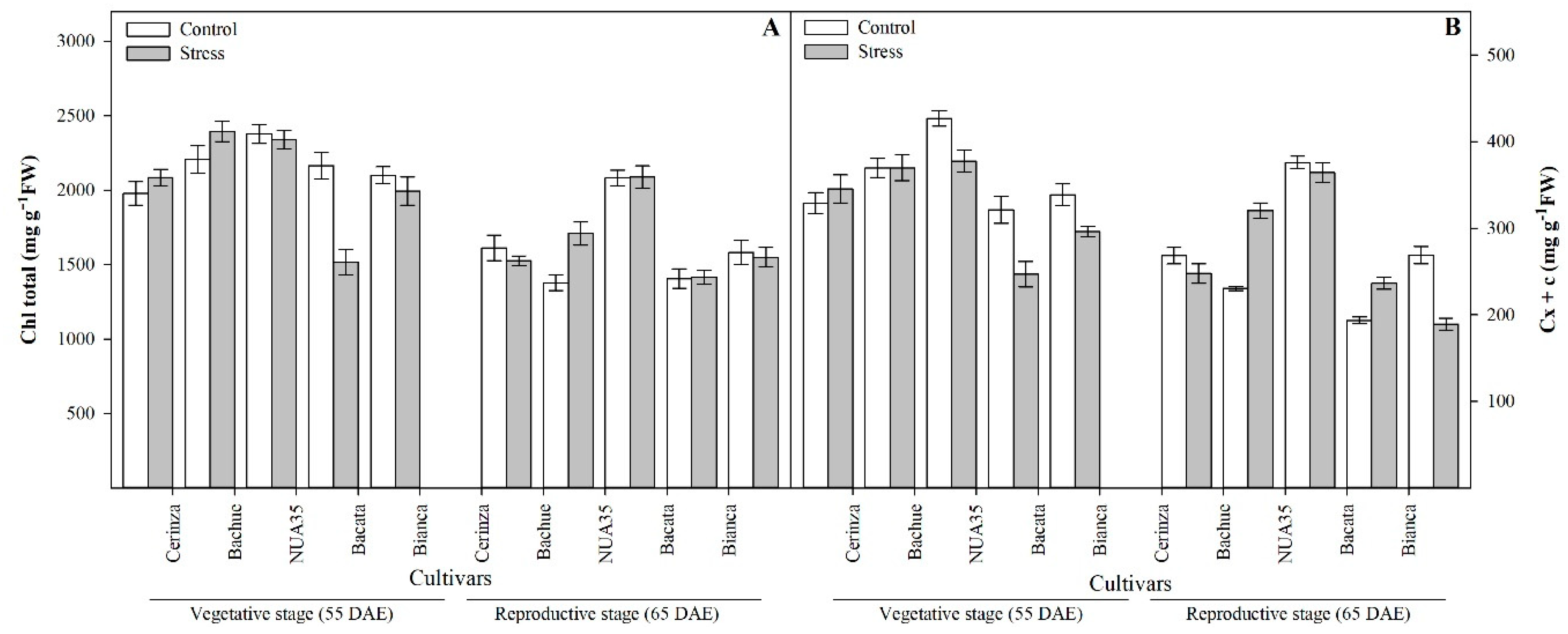
3.1. Chlorophyll and Carotenoids Contents
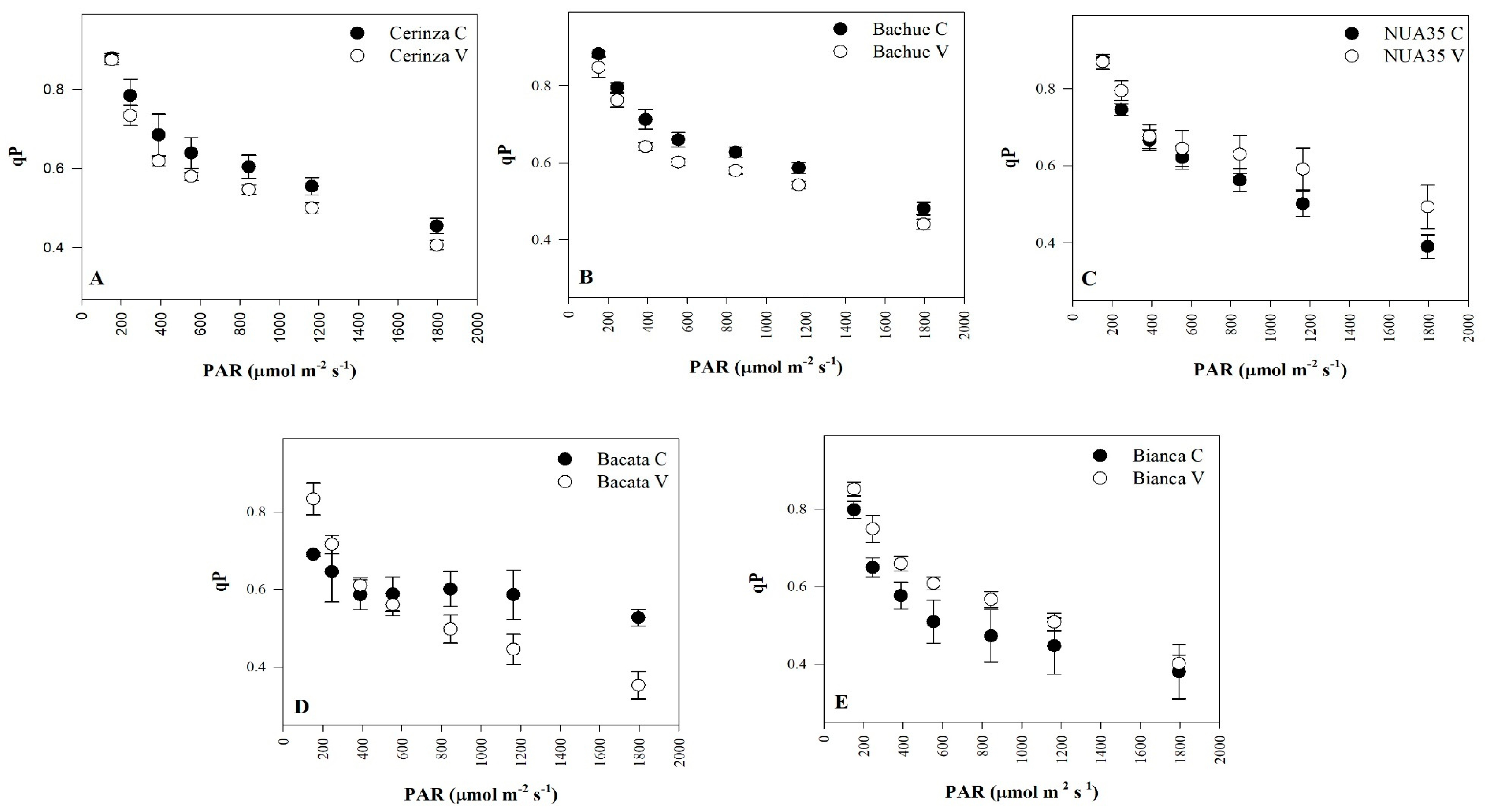
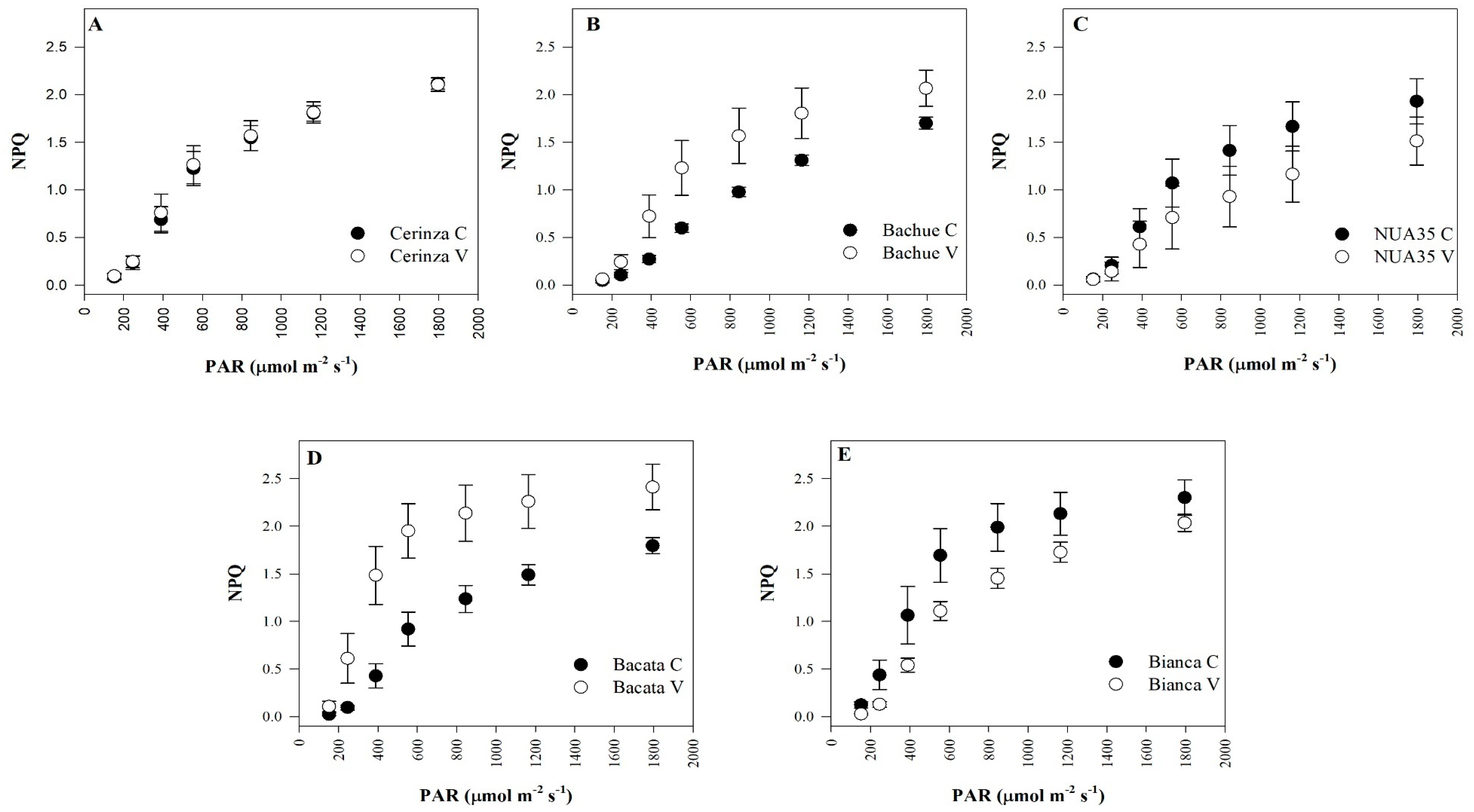
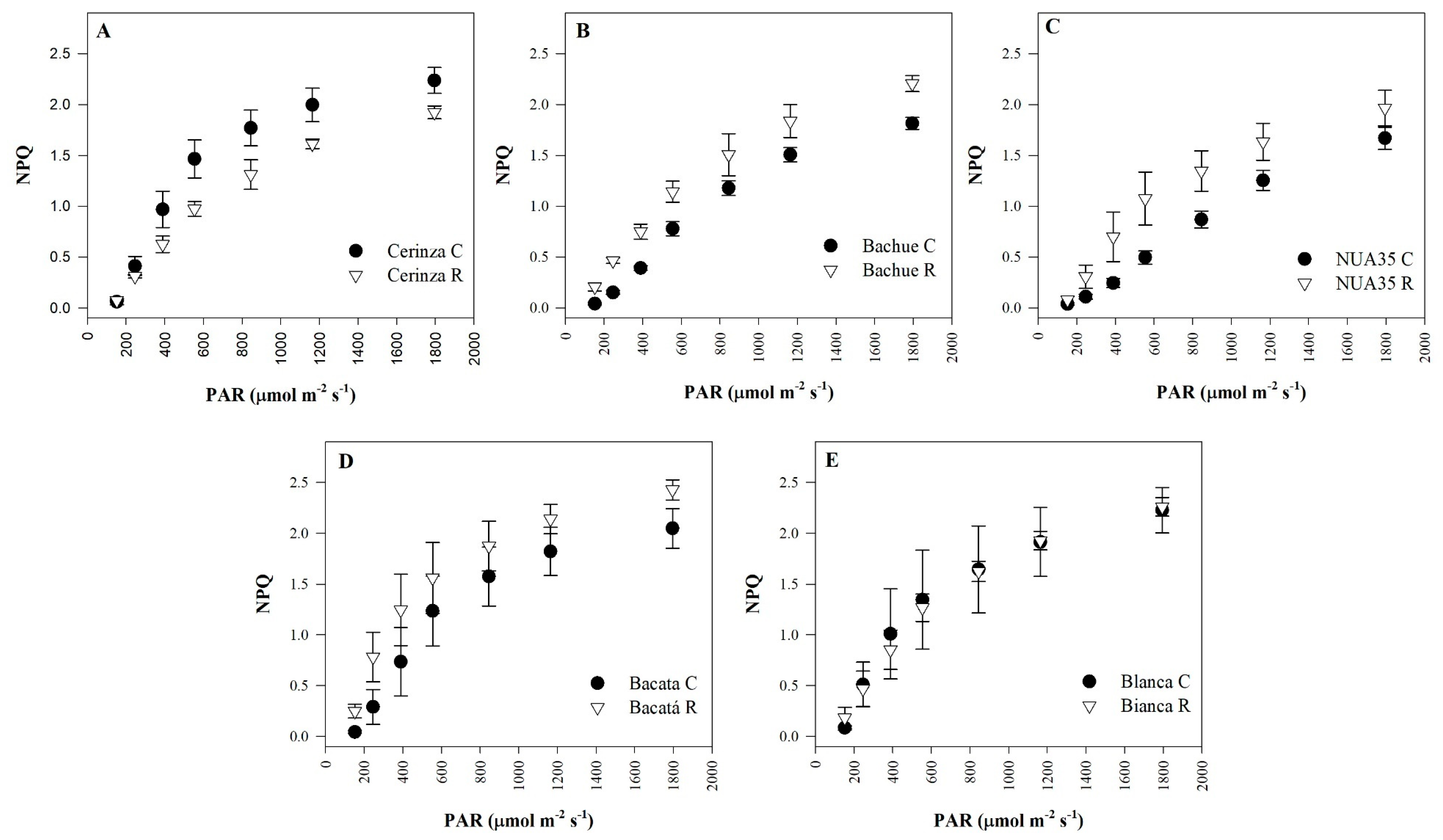
3.2. Fluorescence Parameters
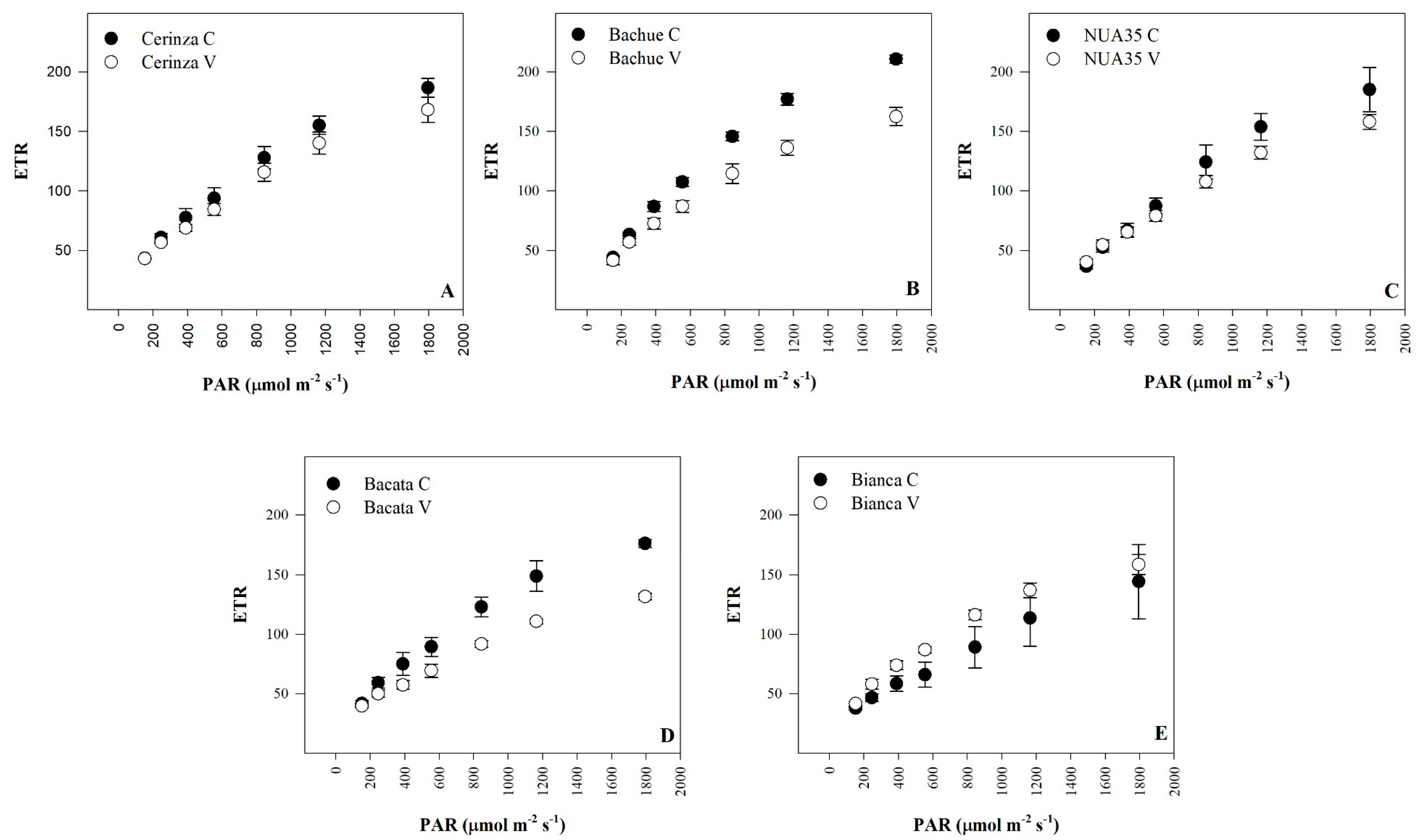
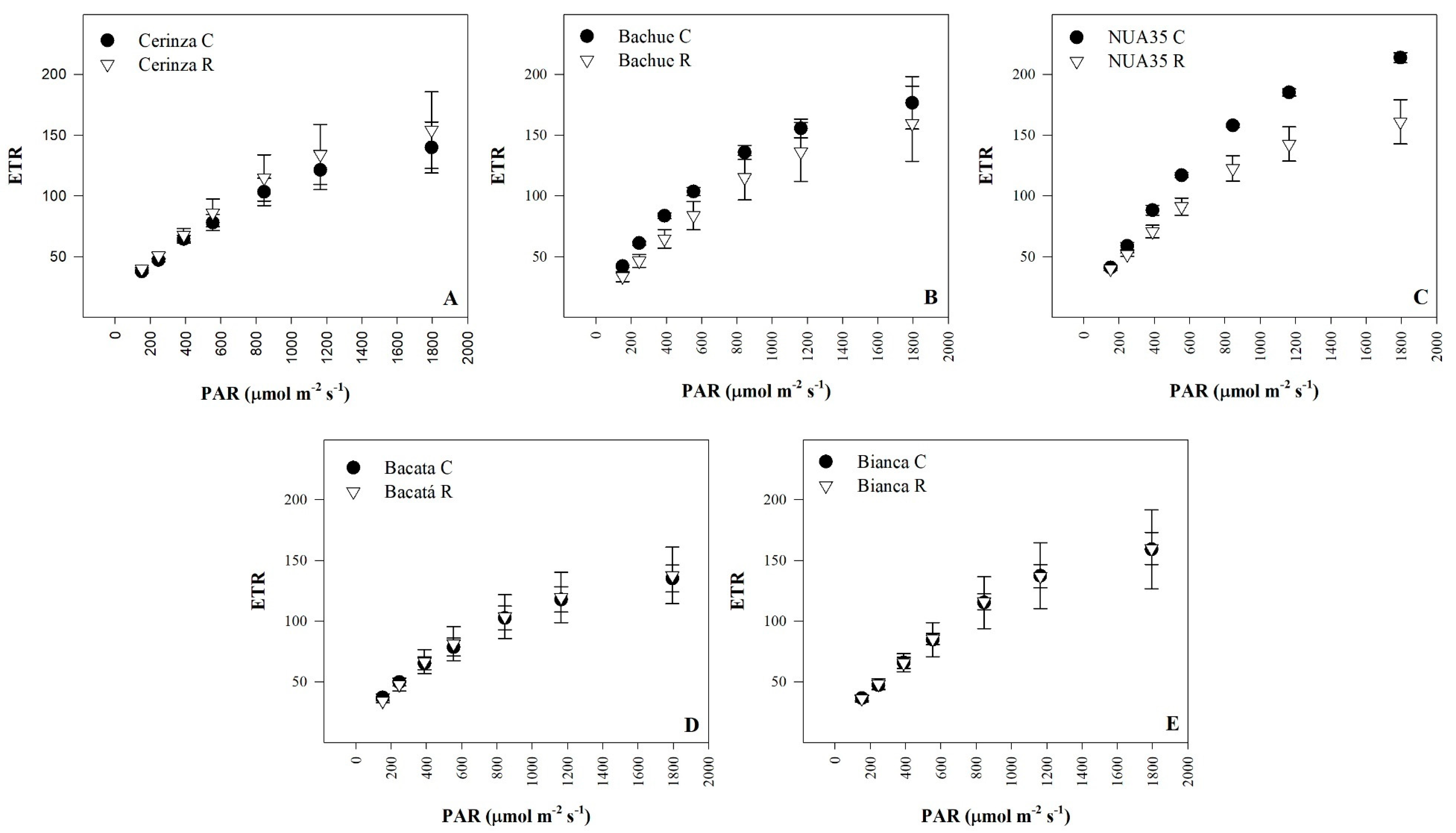
3.3. Rapid Light-Response Curves
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Khoury, C.K.; Bjorkman, A.D.; Dempewolf, H.; Ramirez-Villegas, J.; Guarino, L.; Jarvis, A.; Rieseberg, L.H.; Struik, P.C. Increasing homogeneity in global food supplies and the implications for food security. Proc. Natl. Acad. Sci. USA 2014, 111, 4001–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, T.; Farah, A. Coffee, maté, açaí and beans are the main contributors to the antioxidant capacity of Brazilian’s diet. Eur. J. Nutr. 2017, 56, 1523–1533. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Area Harvested, Yield and Production Quantity 2017: Beans, Dry; Beans, Green. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 15 May 2019).
- Heinemann, A.B.; Ramirez-Villegas, J.; Stone, L.F.; Didonet, A.D. Climate change determined drought stress profiles in rainfed common bean production systems in Brazil. Agric. For. Meteorol. 2017, 246, 64–77. [Google Scholar] [CrossRef]
- Anjum, S.; Xie, A.; Wang, Y.; Saleem, L.C.; Man, M.F.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Mishra, K.B.; Mishra, A.; Novotná, K.; Rapantová, B.; Hodaňová, P.; Urban, O.; Klem, K. Chlorophyll a fluorescence, under half of the adaptive growth-irradiance, for high-throughput sensing of leaf-water deficit in Arabidopsis thaliana accessions. Plant Methods 2016, 12, 46. [Google Scholar] [CrossRef]
- Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Recent advances in the dissection of drought-stress regulatory networks and strategies for development of drought-tolerant transgenic rice plants. Front. Plant Sci. 2015, 6, 84. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Kouzmanova, M.; Kalaji, H.M.; Yordanov, I.; Krasteva, V.; Alexandrov, V.; Stefanov, D.; Allakhverdiev, S.I.; et al. Drought-induced modifications of photosynthetic electron transport in intact leaves: Analysis and use of neural networks as a tool for a rapid non-invasive estimation. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 1490–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govindjee, E. 63 Years since Kautsky-chlorophyll-a fluorescence. Aust. J. Plant Physiol. 1995, 22, 131–160. [Google Scholar]
- Marques, M.C.; Nascimento, C.W.A.; da Silva, A.J.; da Silva Gouveia-Neto, A. Tolerance of an energy crop (Jatropha curcas L.) to zinc and lead assessed by chlorophyll fluorescence and enzyme activity. S. Afr. J. Bot. 2017, 112, 275–282. [Google Scholar] [CrossRef]
- Do Nascimento, C.W.A.; Marques, M.C. Metabolic alterations and X-ray chlorophyll fluorescence for the early detection of lead stress in castor bean (Ricinus communis) plants. Acta Sci. Agron. 2018, 40. [Google Scholar] [CrossRef]
- Silva, A.J.D.; Nascimento, C.W.; Neto, G.; da Silva, A.; Silva Junior, E.A. Effects of silicon on alleviating arsenic toxicity in maize plants. Rev. Bras. Ciênc. Solo 2015, 39, 289–296. [Google Scholar] [CrossRef]
- Rosenqvist, E.; van Kooten, O. Chlorophyll fluorescence: A general description and nomenclature. In Practical Applications of Chlorophyll Fluorescence in Plant Biology; Springer: Boston, MA, USA, 2003; pp. 31–77. [Google Scholar]
- Flowers, M.D.; Fiscus, E.L.; Burkey, K.O.; Booker, F.L.; Dubois, J.J.B. Photosynthesis, chlorophyll fluorescence, and yield of snap bean (Phaseolus vulgaris L.) genotypes differing in sensitivity to ozone. Environ. Exp. Bot. 2007, 61, 190–198. [Google Scholar] [CrossRef]
- Sanchez-Reinoso, A.D.; Ligarreto-Moreno, G.A.; Restrepo-Diaz, H. Physiological and Biochemical Responses of Common Bush Bean to Drought. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.Z.; Deng, X.P.; Xu, B.C.; Gao, Z.J.; Ding, W.L. Photosynthetic activity and efficiency of Bothriochloa ischaemum and Lespedeza davurica in mixtures across growth periods under water stress. Acta Physiol. Plant. 2014, 36, 1033–1044. [Google Scholar] [CrossRef]
- Rascher, U.; Liebig, M.; Lüttge, U. Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer on site in the field. Plant Cell Environ. 2000, 23, 1397–1405. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Lukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Feller, C.; Bleiholder, H.; Buhr, L.; Hack, H.; Hess, M.; Klose, R.; Meier, U.; Stauss, R.; Van den Boom, T.; Weber, E. Phänologische Entwicklungsstadien von Gemüsepflanzen: II. Fruchtgemüse und Hülsenfrüchte. Nachrichtenbl. Deut. Pflanzenschutzd 1995, 47, 217–232. [Google Scholar]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E.D., Caldwell, M.M., Eds.; Springer: Heidelberg, Germany, 1994; pp. 49–70. [Google Scholar]
- Wu, Q.; Su, N.; Shen, W.; Cui, J. Analyzing photosynthetic activity and growth of Solanum lycopersicum seedlings exposed to different light qualities. Acta Physiol. Plant. 2014, 36, 1411–1420. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypinski, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef]
- Chavez-Arias, C.C.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis Peruviana L. Seedlings Exposed to Different Short-Term Waterlogging Periods and Fusarium Wilt Infection. Agronomy 2018, 9, 213. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R. Tolerance to stress combination in tomato plants: New insights in the protective role of melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.O.; Rosenqvist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant. 2015, 153, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, S.; Tahmasebi-Sarvestani, Z.; Modarres-Sanavy, S.A.M.; Mokhtassi-Bidgoli, A.; Nicola, S. Effects of water stress and light intensity on chlorophyll fluorescence parameters and pigments of Aloe vera L. Plant Physiol. Biochem. 2016, 106, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Belshe, E.F.; Durako, M.J.; Blum, J.E. Photosynthetic rapid light curves (RLC) of Thalassia testudinum exhibit diurnal variation. J. Exp. Mar. Biol. Ecol. 2007, 342, 253–268. [Google Scholar] [CrossRef]
- Ralph, P.J.; Gademann, R. Rapid light curves: A powerful tool to assess photosynthetic activity. Aquat. Bot. 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Ihnken, S.; Eggert, A.; Beardall, J. Exposure times in rapid light curves affect photosynthetic parameters in algae. Aquat. Bot. 2010, 93, 185–194. [Google Scholar] [CrossRef]
- Serôdio, J.; Vieira, S.; Cruz, S.; Coelho, H. Rapid light-response curves of chlorophyll fluorescence in microalgae: Relationship to steady-state light curves and non-photochemical quenching in benthic diatom-dominated assemblages. Photosynth. Res. 2006, 90, 29–43. [Google Scholar] [CrossRef]
- Hussain, S.; Rao, M.J.; Anjum, M.A.; Ejaz, S.; Zakir, I.; Ali, M.A.; Ahmad, N.; Ahmad, S. Oxidative stress and antioxidant defense in plants under drought conditions. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 207–219. [Google Scholar]
- Kusvuran, S.; Dasgan, H.Y. Effects of drought stress on physiological and biochemical changes in Phaseolus vulgaris L. Legum. Res. 2017, 40, 55–62. [Google Scholar]
- Mibei, E.K.; Ambuko, J.; Giovannoni, J.J.; Onyango, A.N.; Owino, W.O. Carotenoid profiling of the leaves of selected African eggplant accessions subjected to drought stress. Food Sci. Nutr. 2017, 5, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, G.; Rasool, F.; Habib, R.; Javed, T.; Sardar, K.; Ayub, M.M.; Ayub, M.A.; Rasool, A. A review of morphological, physiological and biochemical responses of plants under drought stress conditions. Imp. J. Interdiscip. Res. 2016, 2, 1600–1606. [Google Scholar]
- David, O.A.; Osonubi, O.; Ajiboye, A.A.; Ajewole, T.O. Agronomic Components of Drought Stressed Wheat Plants under Different Soil Properties. Vegetos-An Int. J. Plant Res. 2018, 31, 82–91. [Google Scholar] [CrossRef]
- Sánchez-Reinoso, A.D.; Ligarreto-Moreno, G.A.; Restrepo-Díaz, H. Evaluation of drought indices to identify tolerant genotypes in common bean bush (Phaseolus vulgaris L.). J. Integr. Agric. 2019, 18, 2–10. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Abbreviation | Variation Source | |||||
|---|---|---|---|---|---|---|---|
| Vegetative Stage (55 DAE) | Reproductive Stage (65 DAE) | ||||||
| Stage | Cultivar | Stage × Cultivar | Stage | Cultivar | Stage × Cultivar | ||
| Total Chlorophyll | Chl total | *** | *** | *** | NS | *** | * |
| Carotenoids | Cx + c | *** | *** | * | ** | *** | *** |
| Maximum efficiency of PSII | Fv/Fm | *** | ** | ** | * | NS | NS |
| Photochemical quenching | qP | NS | NS | NS | NS | * | NS |
| Non-photochemical quenching | NPQ | NS | * | * | *** | *** | NS |
| Initial slope | α | NS | ** | ** | NS | * | NS |
| Maximum electron transport rate | ETRmax | NS | NS | NS | *** | *** | *** |
| Light saturation point | Ik | NS | NS | ** | NS | * | NS |
| Treatment | Vegetative Stage (55 DAE) | Treatment | Reproductive Stage (65 DAE) | ||||
|---|---|---|---|---|---|---|---|
| Fv/Fm | qP | NPQ | Fv/Fm | qP | NPQ | ||
| Stress stage | Stress stage | ||||||
| Control | 0.78 az | 0.61 | 1.10 | Control | 0.79 a | 0.60 | 1.07 |
| Vegetative | 0.74 b | 0.60 | 1.23 | Reproductive | 0.75 b | 0.63 | 1.21 |
| Significance | * | NS | NSy | NS | ** | NS | NS |
| Cultivar | Cultivar | ||||||
| Cerinza | 0.77 a | 0.63 | 1.12 | Cerinza | 0.77 | 0.56 | 1.27 ab |
| Bachue | 0.76 ab | 0.63 | 0.89 | Bachue | 0.75 | 0.65 | 1.10 ab |
| NUA35 | 0.74 b | 0.62 | 1.16 | NUA35 | 0.76 | 0.66 | 0.97 b |
| Bacata | 0.77 ab | 0.57 | 1.58 | Bacata | 0.75 | 0.58 | 1.60 a |
| Bianca | 0.77 ab | 0.58 | 1.29 | Bianca | 0.75 | 0.63 | 1.41 ab |
| Significance | ** | NS | NS | Significance | NS | NS | *** |
| Interaction | Interaction | ||||||
| Cerinza × C | 0.79 a | 0.64 | 1.22 ab | Cerinza × C | 0.79 | 0.55 | 1.47 |
| Bachue × C | 0.79 a | 0.66 | 0.60 b | Bachue × C | 0.80 | 0.65 | 0.78 |
| NUA35 × C | 0.78 a | 0.62 | 1.07 ab | NUA35 × C | 0.79 | 0.71 | 0.50 |
| Bacata × C | 0.79 a | 0.62 | 0.92 ab | Bacata × C | 0.78 | 0.54 | 1.25 |
| Bianca × C | 0.76 a | 0.51 | 1.69 ab | Bianca × C | 0.79 | 0.57 | 1.35 |
| Cerinza × V | 0.79 a | 0.58 | 1.26 ab | Cerinza × R | 0.77 | 0.57 | 0.98 |
| Bachue × V | 0.74 ab | 0.64 | 1.23 ab | Bachue × R | 0.73 | 0.65 | 1.14 |
| NUA35 × V | 0.67 b | 0.65 | 0.71 ab | NUA35 × R | 0.76 | 0.64 | 1.08 |
| Bacata × V | 0.75 a | 0.54 | 1.86 a | Bacata × R | 0.73 | 0.66 | 1.56 |
| Bianca × V | 0.77 a | 0.61 | 1.11 ab | Bianca × R | 0.77 | 0.62 | 1.27 |
| Significance | * | NS | * | Significance | NS | NS | NS |
| xCV (%) | 4.31 | 11.40 | 42.44 | CV (%) | 5.63 | 13.04 | 32.0 |
| Vegetative Stage (55 DAE) | Reproductive Stage (65 DAE) | ||||||
|---|---|---|---|---|---|---|---|
| Treatment | α | ETRmax | Ik | Treatment | α | ETRmax | Ik |
| (μmol∙m−2∙s−1) | (μmol∙m−2∙s−1) | (μmol∙m−2∙s−1) | (μmol∙m−2∙s−1) | ||||
| Stress stage | Stress stage | ||||||
| Control | 0.23 a z | 207.42 | 936.47 | Control | 0.24 | 190.38 a | 795.48 |
| Vegetative | 0.21 ab | 191.87 | 876.60 | Reproductive | 0.22 | 165.71 b | 811.98 |
| Significance | NSy | NS | NS | Significance | NS | *** | NS |
| Interaction | Interaction | ||||||
| Cerinza | 0.22 ab | 241.35 | 1122.83 | Cerinza | 0.22 b | 151.75 b | 768.83 ab |
| Bachue | 0.24 a | 209.75 | 896.54 | Bachue | 0.23 ab | 194.09 a | 830.91 ab |
| NUA35 | 0.22 ab | 204.39 | 916.10 | NUA35 | 0.24 a | 204.87 a | 893.33 ab |
| Bacata | 0.19 b | 169.94 | 850.35 | Bacata | 0.19 b | 150.75 b | 685.04 b |
| Bianca | 0.21 b | 203.09 | 976.53 | Bianca | 0.21 b | 188.75 a | 850.14 a |
| Significance | ** | NS | NS | Significance | * | *** | * |
| Interaction | Interaction | ||||||
| Cerinza × C | 0.23 abc | 232.35 | 1052.72 ab | Cerinza × C | 0.21 | 167.64 bcd | 773.82 |
| Bachue × C | 0.27 a | 221.03 | 838.06 ab | Bachue × C | 0.27 | 195.83 b | 728.18 |
| NUA35 × C | 0.24 a | 185.92 | 786.85 b | NUA35 × C | 0.28 | 251.22 a | 910.54 |
| Bacata × C | 0.23 abc | 207.85 | 884.87 ab | Bacata × C | 0.21 | 159.22 cd | 707.97 |
| Bianca × C | 0.16 c | 189.95 | 1119.85 a | Bianca × C | 0.21 | 188.56 bc | 856.91 |
| Cerinza × V | 0.21 abc | 201.21 | 953.87 ab | Cerinza × R | 0.23 | 145.86 d | 758.85 |
| Bachue × V | 0.23 ab | 196.73 | 844.59 ab | Bachue × R | 0.20 | 196.91 bc | 905.40 |
| NUA35 × V | 0.21 abc | 235.22 | 976.33 ab | NUA35 × R | 0.24 | 158.5 bcd | 774.97 |
| Bacata × V | 0.17 bc | 148.54 | 838.19 ab | Bacata × R | 0.21 | 152.89 bcd | 718.17 |
| Bianca × V | 0.23 abc | 177.65 | 769.99 b | Bianca × R | 0.21 | 188.93 bc | 902.48 |
| Significance | ** | NS | ** | Significance | NS | *** | NS |
| CVx (%) | 12.50 | 19.66 | 13.16 | CVx (%) | 14.22 | 10.21 | 13.76 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Reinoso, A.D.; Ligarreto-Moreno, G.A.; Restrepo-Díaz, H. Chlorophyll α Fluorescence Parameters as an Indicator to Identify Drought Susceptibility in Common Bush Bean. Agronomy 2019, 9, 526. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090526
Sánchez-Reinoso AD, Ligarreto-Moreno GA, Restrepo-Díaz H. Chlorophyll α Fluorescence Parameters as an Indicator to Identify Drought Susceptibility in Common Bush Bean. Agronomy. 2019; 9(9):526. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090526
Chicago/Turabian StyleSánchez-Reinoso, Alefsi David, Gustavo Adolfo Ligarreto-Moreno, and Hermann Restrepo-Díaz. 2019. "Chlorophyll α Fluorescence Parameters as an Indicator to Identify Drought Susceptibility in Common Bush Bean" Agronomy 9, no. 9: 526. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090526