
The In Silico Identification of Potential Members of the Ded1/DDX3 Subfamily of DEAD-Box RNA Helicases from the Protozoan Parasite Leishmania infantum and Their Analyses in Yeast

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Acquisition and Analyses
2.2. Preparation of L. infantum and T. brucei DNA
2.3. Cloning and Yeast Manipulations
2.4. Synthetic Gene Construction
2.5. Ded1 Chimeras of the LINF and TRYP Proteins
2.6. Western Blot Analyses
3. Results
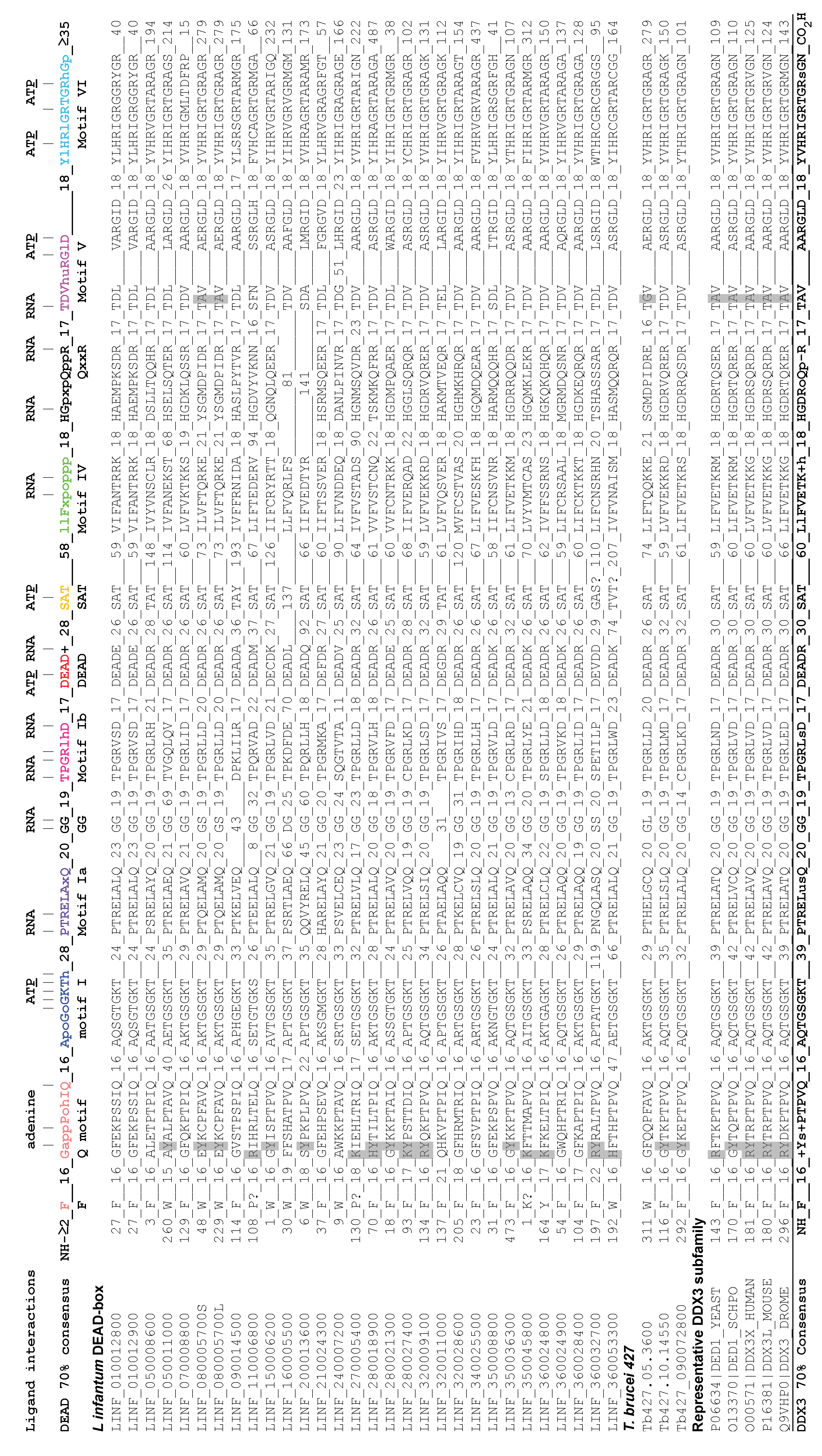
3.1. Bioinformatic Analyses
3.1.1. Identification of L. infantum DEAD-Box Proteins
3.1.2. Phylogenetic Relationship of the L. infantum Proteins and the Ded1/DDX3 Subfamily
3.1.3. Sequence Alignments of the DEAD-Box Proteins
3.1.4. Identification of Ded1/DDX3-Specific Characteristics
3.1.5. Identification of Ded1/DDX3-Specific Motifs outside the RecA-Like Core
3.1.6. Comparisons between the L. infantum and T. brucei Ded1/DDX3-Like Proteins
3.2. Yeast Complementation
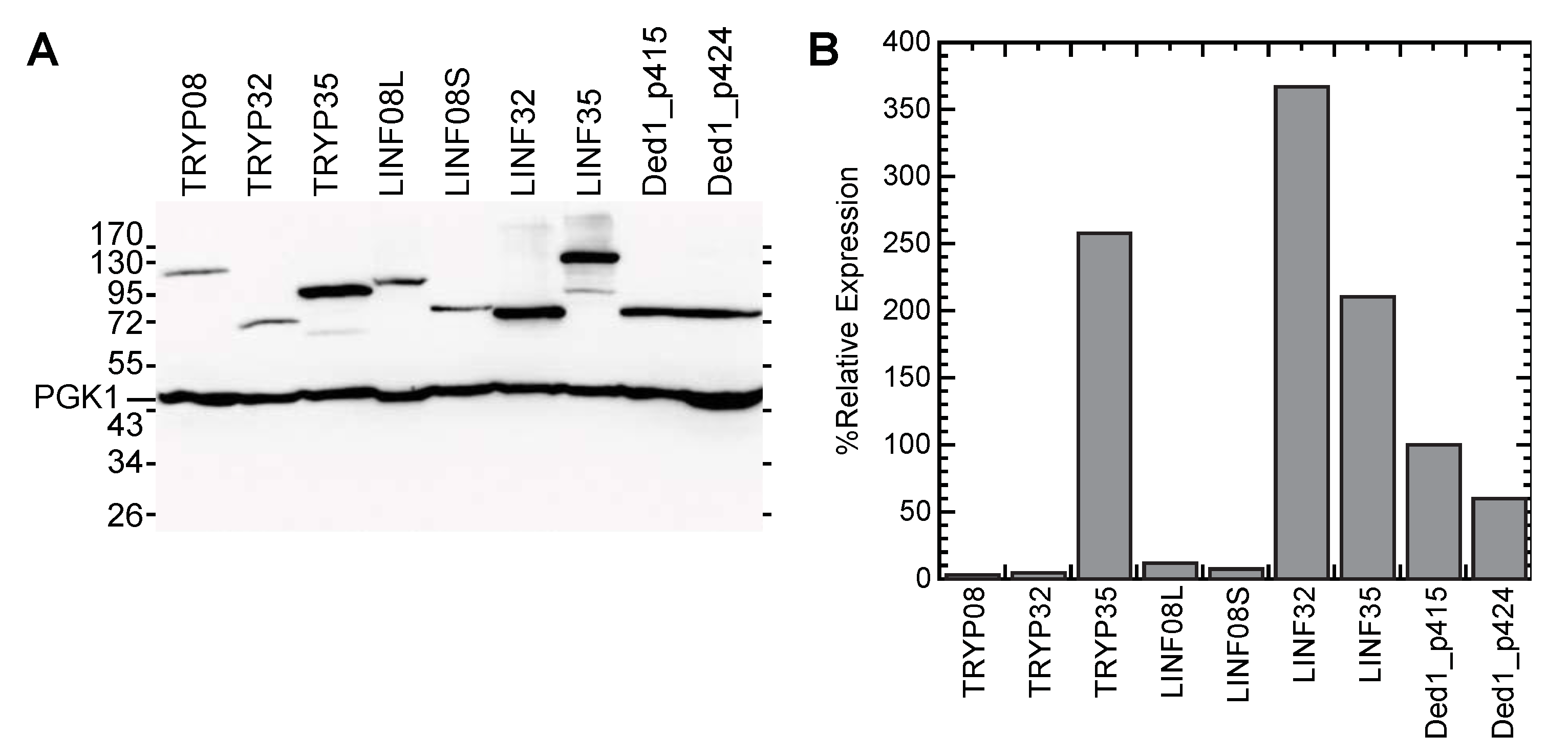
3.3. Protein Expression
3.4. Synthetic Genes
3.5. Ded1 Chimeras
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. Correction: A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004770. [Google Scholar] [CrossRef] [Green Version]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef]
- Akhoundi, M.; Downing, T.; Votýpka, J.; Kuhls, K.; Lukeš, J.; Cannet, A.; Ravel, C.; Marty, P.; Delaunay, P.; Kasbari, M.; et al. Leishmania infections: Molecular targets and diagnosis. Mol. Aspects Med. 2017, 57, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Querol, E.; Rosales, C. Control of Phagocytosis by Microbial Pathogens. Front. Immunol. 2017, 8, 1368. [Google Scholar] [CrossRef] [Green Version]
- Liévin-Le Moal, V.; Loiseau, P.M. Leishmania hijacking of the macrophage intracellular compartments. FEBS J. 2016, 283, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Arango Duque, G.; Descoteaux, A. Leishmania survival in the macrophage: Where the ends justify the means. Curr. Opin. Microbiol. 2015, 26, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef]
- Guizani, I.; Mukhtar, M.; Alvar, J.; Ben Abderrazak, S.; Shaw, J. Encyclopedia of Environmental Health; Nriagu, J.O., Ed.; Elsevier: Burlington, VT, USA, 2011; pp. 453–480. [Google Scholar]
- Herwaldt, B.L. Leishmaniasis. Lancet 1999, 354, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. 2020. Available online: http://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 23 November 2020).
- Didwania, N.; Shadab, M.; Sabur, A.; Ali, N. Alternative to Chemotherapy-The Unmet Demand against Leishmaniasis. Front. Immunol. 2017, 8, 1779. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Engwerda, C. Vaccines to prevent leishmaniasis. Clin. Transl. Immunol. 2014, 3, e13. [Google Scholar] [CrossRef]
- Seyed, N.; Peters, N.C.; Rafati, S. Translating Observations From Leishmanization Into Non-Living Vaccines: The Potential of Dendritic Cell-Based Vaccination Strategies Against Leishmania. Front. Immunol. 2018, 9, 1227. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in humans: Drug or vaccine therapy? Drug Des. Dev. Ther. 2018, 12, 25–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hefnawy, A.; Berg, M.; Dujardin, J.C.; De Muylder, G. Exploiting Knowledge on Leishmania Drug Resistance to Support the Quest for New Drugs. Trends Parasitol. 2017, 33, 162–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roatt, B.M.; de Oliveira Cardoso, J.M.; De Brito, R.C.F.; Coura-Vital, W.; de Oliveira Aguiar-Soares, R.D.; Reis, A.B. Recent advances and new strategies on leishmaniasis treatment. Appl. Microbiol. Biotechnol. 2020, 104, 8965–8977. [Google Scholar] [CrossRef] [PubMed]
- Cordin, O.; Banroques, J.; Tanner, N.K.; Linder, P. The DEAD-box protein family of RNA helicases. Gene 2006, 367, 17–37. [Google Scholar] [CrossRef]
- Linder, P.; Jankowsky, E. From unwinding to clamping—The DEAD box RNA helicase family. Nat. Rev. Mol. Cell Biol. 2011, 12, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Putnam, A.A.; Jankowsky, E. DEAD-box helicases as integrators of RNA, nucleotide and protein binding. Biochim. Biophys. Acta 2013, 1829, 884–893. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.K.; Raney, K.D. Superfamily 2 helicases. Front. Biosci. 2012, 17, 2070–2088. [Google Scholar] [CrossRef]
- Fairman-Williams, M.E.; Guenther, U.P.; Jankowsky, E. SF1 and SF2 helicases: Family matters. Curr. Opin. Struct. Biol. 2010, 20, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Banroques, J.; Cordin, O.; Doère, M.; Linder, P.; Tanner, N.K. Analyses of the functional regions of DEAD-box RNA “helicases” with deletion and chimera constructs tested in vivo and in vitro. J. Mol. Biol. 2011, 413, 451–472. [Google Scholar] [CrossRef] [Green Version]
- de la Cruz, J.; Kressler, D.; Linder, P. Unwinding RNA in Saccharomyces cerevisiae: DEAD-box proteins and related families. Trends Biochem. Sci. 1999, 24, 192–198. [Google Scholar] [CrossRef]
- Cencic, R.; Pelletier, J. Throwing a monkey wrench in the motor: Targeting DExH/D box proteins with small molecule inhibitors. Biochim. Biophys. Acta 2013, 1829, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Bol, G.M.; Xie, M.; Raman, V. DDX3, a potential target for cancer treatment. Mol. Cancer 2015, 14, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heerma van Voss, M.R.; van Diest, P.J.; Raman, V. Targeting RNA helicases in cancer: The translation trap. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 510–520. [Google Scholar] [CrossRef]
- Kwong, A.D.; Rao, B.G.; Jeang, K.T. Viral and cellular RNA helicases as antiviral targets. Nat. Rev. Drug Discov. 2005, 4, 845–853. [Google Scholar] [CrossRef]
- Shadrick, W.R.; Ndjomou, J.; Kolli, R.; Mukherjee, S.; Hanson, A.M.; Frick, D.N. Discovering new medicines targeting helicases: Challenges and recent progress. J. Biomol. Screen. 2013, 18, 761–781. [Google Scholar] [CrossRef] [Green Version]
- Abdelkrim, Y.Z.; Banroques, J.; Kyle Tanner, N. Known Inhibitors of RNA Helicases and Their Therapeutic Potential. Methods Mol. Biol. 2021, 2209, 35–52. [Google Scholar] [CrossRef]
- Sharma, D.; Jankowsky, E. The Ded1/DDX3 subfamily of DEAD-box RNA helicases. Crit Rev. Biochem. Mol. Biol. 2014, 49, 343–360. [Google Scholar] [CrossRef]
- Chang, T.C.; Liu, W.S. The molecular evolution of PL10 homologs. BMC Evol. Biol. 2010, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Tarn, W.Y.; Chang, T.H. The current understanding of Ded1p/DDX3 homologs from yeast to human. RNA Biol. 2009, 6, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Rosner, A.; Rinkevich, B. The DDX3 subfamily of the DEAD box helicases: Divergent roles as unveiled by studying different organisms and in vitro assays. Curr. Med. Chem. 2007, 14, 2517–2525. [Google Scholar] [CrossRef] [PubMed]
- Senissar, M.; Le Saux, A.; Belgareh-Touze, N.; Adam, C.; Banroques, J.; Tanner, N.K. The DEAD-box helicase Ded1 from yeast is an mRNP cap-associated protein that shuttles between the cytoplasm and nucleus. Nucleic Acids Res. 2014, 42, 10005–10022. [Google Scholar] [CrossRef] [Green Version]
- Soto-Rifo, R.; Ohlmann, T. The role of the DEAD-box RNA helicase DDX3 in mRNA metabolism. Wiley Interdiscip. Rev. RNA 2013, 4, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, P.; Kedersha, N.; Anderson, P. Stress Granules and Processing Bodies in Translational Control. Cold Spring Harb. Perspect. Biol. 2019, 11. [Google Scholar] [CrossRef]
- Guzikowski, A.R.; Chen, Y.S.; Zid, B.M. Stress-induced mRNP granules: Form and function of processing bodies and stress granules. Wiley Interdiscip Rev. RNA 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Hondele, M.; Sachdev, R.; Heinrich, S.; Wang, J.; Vallotton, P.; Fontoura, B.M.A.; Weis, K. DEAD-box ATPases are global regulators of phase-separated organelles. Nature 2019, 573, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Mao, Y.; Zhou, J.; Zhao, Y.; Cao, Y.; Chen, X. Multifunctional DDX3: Dual roles in various cancer development and its related signaling pathways. Am. J. Cancer Res. 2016, 6, 387–402. [Google Scholar] [PubMed]
- Kukhanova, M.K.; Karpenko, I.L.; Ivanov, A.V. DEAD-box RNA Helicase DDX3: Functional Properties and Development of DDX3 Inhibitors as Antiviral and Anticancer Drugs. Molecules 2020, 25, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brai, A.; Fazi, R.; Tintori, C.; Zamperini, C.; Bugli, F.; Sanguinetti, M.; Stigliano, E.; Esté, J.; Badia, R.; Franco, S.; et al. Human DDX3 protein is a valuable target to develop broad spectrum antiviral agents. Proc. Natl. Acad. Sci. USA 2016, 113, 5388–5393. [Google Scholar] [CrossRef] [Green Version]
- Garbelli, A.; Radi, M.; Falchi, F.; Beermann, S.; Zanoli, S.; Manetti, F.; Dietrich, U.; Botta, M.; Maga, G. Targeting the human DEAD-box polypeptide 3 (DDX3) RNA helicase as a novel strategy to inhibit viral replication. Curr. Med. Chem. 2011, 18, 3015–3027. [Google Scholar] [CrossRef]
- Abdelhaleem, M. RNA helicases: Regulators of differentiation. Clin. Biochem. 2005, 38, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Lasko, P. The DEAD-box helicase Vasa: Evidence for a multiplicity of functions in RNA processes and developmental biology. Biochim. Biophys. Acta 2013, 1829, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Noce, T.; Okamoto-Ito, S.; Tsunekawa, N. Vasa homolog genes in mammalian germ cell development. Cell Struct. Funct. 2001, 26, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Poon, J.; Wessel, G.M.; Yajima, M. An unregulated regulator: Vasa expression in the development of somatic cells and in tumorigenesis. Dev. Biol. 2016, 415, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. A revised six-kingdom system of life. Biol Rev. Camb. Philos. Soc. 1998, 73, 203–266. [Google Scholar] [CrossRef]
- Fernandes, A.P.; Nelson, K.; Beverley, S.M. Evolution of nuclear ribosomal RNAs in kinetoplastid protozoa: Perspectives on the age and origins of parasitism. Proc. Natl. Acad. Sci. USA 1993, 90, 11608–11612. [Google Scholar] [CrossRef] [Green Version]
- Karamysheva, Z.N.; Gutierrez Guarnizo, S.A.; Karamyshev, A.L. Regulation of Translation in the Protozoan Parasite Leishmania. Int. J. Mol. Sci. 2020, 21, 2981. [Google Scholar] [CrossRef]
- Clayton, C.E. Gene expression in Kinetoplastids. Curr. Opin. Microbiol. 2016, 32, 46–51. [Google Scholar] [CrossRef]
- De Pablos, L.M.; Ferreira, T.R.; Walrad, P.B. Developmental differentiation in Leishmania lifecycle progression: Post-transcriptional control conducts the orchestra. Curr. Opin. Microbiol. 2016, 34, 82–89. [Google Scholar] [CrossRef]
- Abdelkrim, Y.Z.; Harigua-Souiai, E.; Barhoumi, M.; Banroques, J.; Blondel, A.; Guizani, I.; Tanner, N.K. The steroid derivative 6-aminocholestanol inhibits the DEAD-box helicase eIF4A (LieIF4A) from the Trypanosomatid parasite Leishmania by perturbing the RNA and ATP binding sites. Mol. Biochem. Parasitol. 2018, 226, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Harigua-Souiai, E.; Abdelkrim, Y.Z.; Bassoumi-Jamoussi, I.; Zakraoui, O.; Bouvier, G.; Essafi-Benkhadir, K.; Banroques, J.; Desdouits, N.; Munier-Lehmann, H.; Barhoumi, M.; et al. Identification of novel leishmanicidal molecules by virtual and biochemical screenings targeting Leishmania eukaryotic translation initiation factor 4A. PLoS Negl. Trop. Dis. 2018, 12, e0006160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslett, M.; Aurrecoechea, C.; Berriman, M.; Brestelli, J.; Brunk, B.P.; Carrington, M.; Depledge, D.P.; Fischer, S.; Gajria, B.; Gao, X.; et al. TriTrypDB: A functional genomic resource for the Trypanosomatidae. Nucleic Acids Res. 2010, 38, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Banroques, J.; Tanner, N.K. Bioinformatics and biochemical methods to study the structural and functional elements of DEAD-box RNA helicases. Methods Mol. Biol. 2015, 1259, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Yachdav, G.; Kloppmann, E.; Kajan, L.; Hecht, M.; Goldberg, T.; Hamp, T.; Hönigschmid, P.; Schafferhans, A.; Roos, M.; Bernhofer, M.; et al. PredictProtein--an open resource for online prediction of protein structural and functional features. Nucleic Acids Res. 2014, 42, W337–W343. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- La Cour, T.; Kiemer, L.; Mølgaard, A.; Gupta, R.; Skriver, K.; Brunak, S. Analysis and prediction of leucine-rich nuclear export signals. Protein Eng. Des. Sel. 2004, 17, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [Green Version]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30 (Suppl. 1), S162–S173. [Google Scholar] [CrossRef] [PubMed]
- Oualha, R.; Barhoumi, M.; Marzouki, S.; Harigua-Souiai, E.; Ben Ahmed, M.; Guizani, I. Infection of Human Neutrophils with Leishmania infantum or Leishmania major Strains Triggers Activation and Differential Cytokines Release. Front. Cell Infect. Microbiol. 2019, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guizani, I.; Van Eys, G.J.; Ismail, R.B.; Dellagi, K. Use of recombinant DNA probes for species identification of Old World Leishmania isolates. Am. J. Trop Med. Hyg. 1994, 50, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, C.; Fink, G.R. (Eds.) Guide to Yeast Genetics and Molecular Biology; Academic Press: San Diego, CA, USA, 1991. [Google Scholar]
- Iost, I.; Dreyfus, M.; Linder, P. Ded1p, a DEAD-box protein required for translation initiation in Saccharomyces cerevisiae, is an RNA helicase. J. Biol. Chem. 1999, 274, 17677–17683. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Ang, K.S.; Lee, D.Y. Synthetic Gene Design Using Codon Optimization On-Line (COOL). Methods Mol. Biol. 2017, 1472, 13–34. [Google Scholar] [CrossRef]
- Horvath, A.; Riezman, H. Rapid protein extraction from Saccharomyces cerevisiae. Yeast 1994, 10, 1305–1310. [Google Scholar] [CrossRef]
- Berriman, M.; Ghedin, E.; Hertz-Fowler, C.; Blandin, G.; Renauld, H.; Bartholomeu, D.C.; Lennard, N.J.; Caler, E.; Hamlin, N.E.; Haas, B.; et al. The genome of the African trypanosome Trypanosoma brucei. Science 2005, 309, 416–422. [Google Scholar] [CrossRef] [Green Version]
- Gargantini, P.R.; Lujan, H.D.; Pereira, C.A. In silico analysis of trypanosomatids’ helicases. FEMS Microbiol. Lett. 2012, 335, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Rochette, A.; Raymond, F.; Ubeda, J.M.; Smith, M.; Messier, N.; Boisvert, S.; Rigault, P.; Corbeil, J.; Ouellette, M.; Papadopoulou, B. Genome-wide gene expression profiling analysis of Leishmania major and Leishmania infantum developmental stages reveals substantial differences between the two species. BMC Genom. 2008, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Requena, J.M. LINF_050006300. Medeley Data 2020, V1. [Google Scholar] [CrossRef]
- González-de la Fuente, S.; Peiró-Pastor, R.; Rastrojo, A.; Moreno, J.; Carrasco-Ramiro, F.; Requena, J.M.; Aguado, B. Resequencing of the Leishmania infantum (strain JPCM5) genome and de novo assembly into 36 contigs. Sci. Rep. 2017, 7, 18050. [Google Scholar] [CrossRef] [PubMed]
- Zinoviev, A.; Akum, Y.; Yahav, T.; Shapira, M. Gene duplication in trypanosomatids—Two DED1 paralogs are functionally redundant and differentially expressed during the life cycle. Mol. Biochem. Parasitol. 2012, 185, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, P.K.; Samant, M.; Cloutier, S.; Simard, M.J.; Papadopoulou, B. Apoptosis-like programmed cell death induces antisense ribosomal RNA (rRNA) fragmentation and rRNA degradation in Leishmania. Cell Death Differ. 2012, 19, 1972–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floor, S.N.; Condon, K.J.; Sharma, D.; Jankowsky, E.; Doudna, J.A. Autoinhibitory Interdomain Interactions and Subfamily-specific Extensions Redefine the Catalytic Core of the Human DEAD-box Protein DDX3. J. Biol. Chem. 2016, 291, 2412–2421. [Google Scholar] [CrossRef] [Green Version]
- Shih, J.W.; Tsai, T.Y.; Chao, C.H.; Wu Lee, Y.H. Candidate tumor suppressor DDX3 RNA helicase specifically represses cap-dependent translation by acting as an eIF4E inhibitory protein. Oncogene 2008, 27, 700–714. [Google Scholar] [CrossRef] [Green Version]
- Ozgur, S.; Buchwald, G.; Falk, S.; Chakrabarti, S.; Prabu, J.R.; Conti, E. The conformational plasticity of eukaryotic RNA-dependent ATPases. FEBS J. 2015, 282, 850–863. [Google Scholar] [CrossRef]
- Sengoku, T.; Nureki, O.; Nakamura, A.; Kobayashi, S.; Yokoyama, S. Structural basis for RNA unwinding by the DEAD-box protein Drosophila Vasa. Cell 2006, 125, 287–300. [Google Scholar] [CrossRef] [Green Version]
- The-UniProt-Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [Green Version]
- Tanner, N.K.; Cordin, O.; Banroques, J.; Doere, M.; Linder, P. The Q motif: A newly identified motif in DEAD box helicases may regulate ATP binding and hydrolysis. Mol. Cell 2003, 11, 127–138. [Google Scholar] [CrossRef]
- Banroques, J.; Cordin, O.; Doère, M.; Linder, P.; Tanner, N.K. A conserved phenylalanine of motif IV in superfamily 2 helicases is required for cooperative, ATP-dependent binding of RNA substrates in DEAD-box proteins. Mol. Cell Biol. 2008, 28, 3359–3371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbelli, A.; Beermann, S.; Di Cicco, G.; Dietrich, U.; Maga, G. A motif unique to the human DEAD-box protein DDX3 is important for nucleic acid binding, ATP hydrolysis, RNA/DNA unwinding and HIV-1 replication. PLoS ONE 2011, 6, e19810. [Google Scholar] [CrossRef] [PubMed]
- Askjaer, P.; Bachi, A.; Wilm, M.; Bischoff, F.R.; Weeks, D.L.; Ogniewski, V.; Ohno, M.; Niehrs, C.; Kjems, J.; Mattaj, I.W.; et al. RanGTP-regulated interactions of CRM1 with nucleoporins and a shuttling DEAD-box helicase. Mol. Cell Biol. 1999, 19, 6276–6285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanova, M.; Portalès, P.; Blaineau, C.; Crobu, L.; Bastien, P.; Pagès, M. Inhibition of active nuclear transport is an intrinsic trigger of programmed cell death in trypanosomatids. Cell Death Differ. 2008, 15, 1910–1920. [Google Scholar] [CrossRef] [PubMed]
- Mader, S.; Lee, H.; Pause, A.; Sonenberg, N. The translation initiation factor eIF-4E binds to a common motif shared by the translation factor eIF-4 γ and the translational repressors 4E-binding proteins. Mol. Cell Biol. 1995, 15, 4990–4997. [Google Scholar] [CrossRef] [Green Version]
- Hilliker, A.; Gao, Z.; Jankowsky, E.; Parker, R. The DEAD-box protein Ded1 modulates translation by the formation and resolution of an eIF4F-mRNA complex. Mol. Cell 2011, 43, 962–972. [Google Scholar] [CrossRef] [Green Version]
- Dhalia, R.; Reis, C.R.; Freire, E.R.; Rocha, P.O.; Katz, R.; Muniz, J.R.; Standart, N.; de Melo Neto, O.P. Translation initiation in Leishmania major: Characterisation of multiple eIF4F subunit homologues. Mol. Biochem. Parasitol. 2005, 140, 23–41. [Google Scholar] [CrossRef]
- Ross-Kaschitza, D.; Altmann, M. eIF4E and Interactors from Unicellular Eukaryotes. Int. J. Mol. Sci. 2020, 21, 2170. [Google Scholar] [CrossRef] [Green Version]
- Freire, E.R.; Sturm, N.R.; Campbell, D.A.; de Melo Neto, O.P. The Role of Cytoplasmic mRNA Cap-Binding Protein Complexes in Trypanosoma brucei and Other Trypanosomatids. Pathogens 2017, 6, 55. [Google Scholar] [CrossRef]
- Yoffe, Y.; Zuberek, J.; Lerer, A.; Lewdorowicz, M.; Stepinski, J.; Altmann, M.; Darzynkiewicz, E.; Shapira, M. Binding specificities and potential roles of isoforms of eukaryotic initiation factor 4E in Leishmania. Eukaryot. Cell 2006, 5, 1969–1979. [Google Scholar] [CrossRef] [Green Version]
- Banroques, J.; Doère, M.; Dreyfus, M.; Linder, P.; Tanner, N.K. Motif III in superfamily 2 “helicases” helps convert the binding energy of ATP into a high-affinity RNA binding site in the yeast DEAD-box protein Ded1. J. Mol. Biol. 2010, 396, 949–966. [Google Scholar] [CrossRef] [PubMed]
- Robbins, C.J.; Bou-Dargham, M.J.; Sanchez, K.; Rosen, M.C.; Sang, Q.A. Decoding Somatic Driver Gene Mutations and Affected Signaling Pathways in Human Medulloblastoma Subgroups. J. Cancer 2018, 9, 4596–4610. [Google Scholar] [CrossRef] [PubMed]
- Goos, C.; Dejung, M.; Janzen, C.J.; Butter, F.; Kramer, S. The nuclear proteome of Trypanosoma brucei. PLoS ONE 2017, 12, e0181884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumberg, D.; Müller, R.; Funk, M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene 1995, 156, 119–122. [Google Scholar] [CrossRef]
- Jamieson, D.J.; Beggs, J.D. A suppressor of yeast spp81/ded1 mutations encodes a very similar putative ATP-dependent RNA helicase. Mol. Microbiol. 1991, 5, 805–812. [Google Scholar] [CrossRef]
- Beckham, C.; Hilliker, A.; Cziko, A.M.; Noueiry, A.; Ramaswami, M.; Parker, R. The DEAD-box RNA helicase Ded1p affects and accumulates in Saccharomyces cerevisiae P-bodies. Mol. Biol. Cell 2008, 19, 984–993. [Google Scholar] [CrossRef] [Green Version]
- Mattanovich, D.; Gasser, B.; Hohenblum, H.; Sauer, M. Stress in recombinant protein producing yeasts. J. Biotechnol. 2004, 113, 121–135. [Google Scholar] [CrossRef]
- Fullam, A.; Schröder, M. DExD/H-box RNA helicases as mediators of anti-viral innate immunity and essential host factors for viral replication. Biochim. Biophys. Acta 2013, 1829, 854–865. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, P.K.; Zghidi-Abouzid, O.; Samant, M.; Dumas, C.; Aguiar, B.G.; Estaquier, J.; Papadopoulou, B. DDX3 DEAD-box RNA helicase plays a central role in mitochondrial protein quality control in Leishmania. Cell Death Dis. 2016, 7, e2406. [Google Scholar] [CrossRef]
- Mochizuki, K.; Nishimiya-Fujisawa, C.; Fujisawa, T. Universal occurrence of the vasa-related genes among metazoans and their germline expression in Hydra. Dev. Genes Evol. 2001, 211, 299–308. [Google Scholar] [CrossRef]
- Cruz, A.K.; Freitas-Castro, F. Genome and transcriptome analyses of Leishmania spp.: Opening Pandora’s box. Curr. Opin. Microbiol. 2019, 52, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Sulkowska, A.; Auber, A.; Sikorski, P.J.; Silhavy, D.N.; Auth, M.; Sitkiewicz, E.; Jean, V.; Merret, R.M.; Bousquet-Antonelli, C.C.; Kufel, J. RNA Helicases from the DEA(D/H)-Box Family Contribute to Plant NMD Efficiency. Plant. Cell Physiol. 2020, 61, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Zinoviev, A.; Léger, M.; Wagner, G.; Shapira, M. A novel 4E-interacting protein in Leishmania is involved in stage-specific translation pathways. Nucleic Acids Res. 2011, 39, 8404–8415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brotherton, M.C.; Racine, G.; Foucher, A.L.; Drummelsmith, J.; Papadopoulou, B.; Ouellette, M. Analysis of stage-specific expression of basic proteins in Leishmania infantum. J. Proteome Res. 2010, 9, 3842–3853. [Google Scholar] [CrossRef] [PubMed]
- Nirujogi, R.S.; Pawar, H.; Renuse, S.; Kumar, P.; Chavan, S.; Sathe, G.; Sharma, J.; Khobragade, S.; Pande, J.; Modak, B.; et al. Moving from unsequenced to sequenced genome: Reanalysis of the proteome of Leishmania donovani. J. Proteom. 2014, 97, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Sanchiz, Á.; Morato, E.; Rastrojo, A.; Camacho, E.; González-de la Fuente, S.G.; Marina, A.; Aguado, B.; Requena, J.M. The Experimental Proteome of Leishmania infantum Promastigote and Its Usefulness for Improving Gene Annotations. Genes 2020, 11, 1036. [Google Scholar] [CrossRef]
- Pandey, S.C.; Pande, V.; Samant, M. DDX3 DEAD-box RNA helicase (Hel67) gene disruption impairs infectivity of Leishmania donovani and induces protective immunity against visceral leishmaniasis. Sci Rep. 2020, 10, 18218. [Google Scholar] [CrossRef]
- de Pablos, L.M.; Ferreira, T.R.; Dowle, A.A.; Forrester, S.; Parry, E.; Newling, K.; Walrad, P.B. The mRNA-bound Proteome of Leishmania mexicana: Novel Genetic Insight into an Ancient Parasite. Mol. Cell Proteom. 2019, 18, 1271–1284. [Google Scholar] [CrossRef] [Green Version]
- Alsford, S.; Turner, D.J.; Obado, S.O.; Sanchez-Flores, A.; Glover, L.; Berriman, M.; Hertz-Fowler, C.; Horn, D. High-throughput phenotyping using parallel sequencing of RNA interference targets in the African trypanosome. Genome Res. 2011, 21, 915–924. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Identification b | MW (Da) | Size (aa) | %Identity c | pKi d | %G/C |
|---|---|---|---|---|---|---|
| Ded1_S.c. | P06634 | 65,550 | 605 | 100.0 | 8.22 | 46.1 |
| Ded1_S.p. | O13370 | 69,756 | 636 | 68.0 | 8.80 | 45.8 |
| DDX3X_Hu | O00571 | 73,241 | 662 | 62.0 | 7.17 | 45.9 |
| DDX3L_Mu | P16381 | 73,138 | 660 | 61.0 | 7.17 | 50.7 |
| DDX3_D.m. | Q9VHP0 | 85,078 | 798 | 61.0 | 7.70 | 56.4 |
| LINF08S | LINF_080005700 | 75,345 | 686 | 31.0 | 6.70 | 61.2 |
| LINF08L | LINF_080005700 | 93,670 | 867 | 31.0 | 6.14 | 61.4 |
| LINF32 | LINF_320009100 | 66,806 | 615 | 51.0 | 9.04 | 64.6 |
| LINF35 | LINF_350036300 | 100,533 | 925 | 46.0 | 6.40 | 65.4 |
| TRYP08 | Tb427.05.3600 | 103,320 | 949 | 33.0 | 6.20 | 50.2 |
| TRYP32 | Tb427.10.14550 | 66,562 | 619 | 51.0 | 9.25 | 55.2 |
| TRYP35 | Tb427_090072800 | 82,641 | 736 | 49.0 | 8.78 | 53.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokdadi, M.; Abdelkrim, Y.Z.; Banroques, J.; Huvelle, E.; Oualha, R.; Yeter-Alat, H.; Guizani, I.; Barhoumi, M.; Tanner, N.K. The In Silico Identification of Potential Members of the Ded1/DDX3 Subfamily of DEAD-Box RNA Helicases from the Protozoan Parasite Leishmania infantum and Their Analyses in Yeast. Genes 2021, 12, 212. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020212
Mokdadi M, Abdelkrim YZ, Banroques J, Huvelle E, Oualha R, Yeter-Alat H, Guizani I, Barhoumi M, Tanner NK. The In Silico Identification of Potential Members of the Ded1/DDX3 Subfamily of DEAD-Box RNA Helicases from the Protozoan Parasite Leishmania infantum and Their Analyses in Yeast. Genes. 2021; 12(2):212. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020212
Chicago/Turabian StyleMokdadi, Molka, Yosser Zina Abdelkrim, Josette Banroques, Emmeline Huvelle, Rafeh Oualha, Hilal Yeter-Alat, Ikram Guizani, Mourad Barhoumi, and N. Kyle Tanner. 2021. "The In Silico Identification of Potential Members of the Ded1/DDX3 Subfamily of DEAD-Box RNA Helicases from the Protozoan Parasite Leishmania infantum and Their Analyses in Yeast" Genes 12, no. 2: 212. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020212