
Filtering Effect of Rhinogobio cylindricus Gut Microbiota Relieved Influence of the Three Gorges Dam on the Gut Microbiota Composition


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval and Consent to Participate
2.2. Sampling Area and Sample Collection
2.3. Determination of Water Physicochemical Factors
2.4. DNA Extraction and High-Throughput Sequencing
2.5. Data Analysis
3. Results
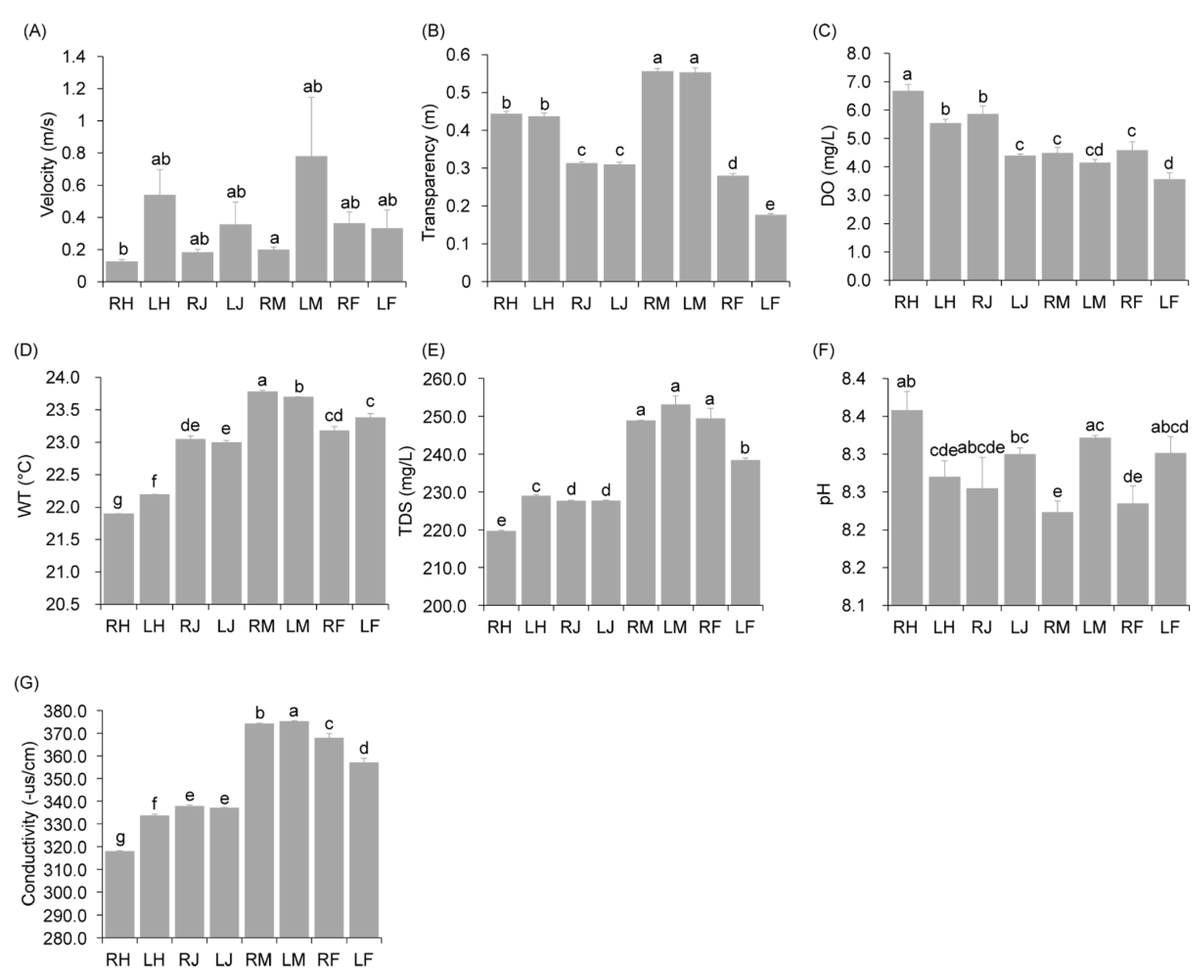
3.1. Changes in Water Physicochemical Indexes in Various Sampling Sites and Fish Biological Indexes
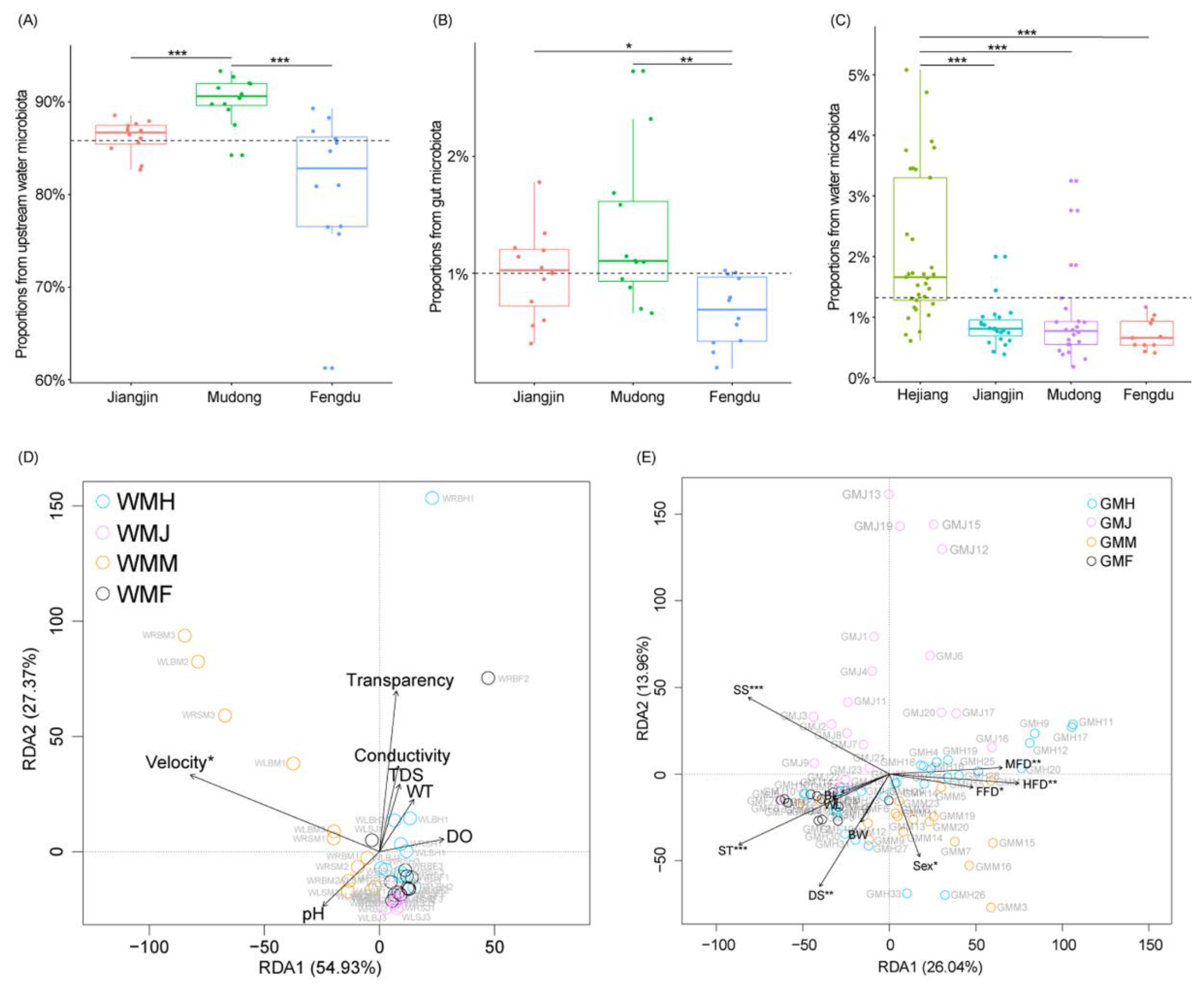
3.2. Changes in Water and R. cylindricus Gut Microbiota Compositions
3.3. Factors That Influence R. cylindricus Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Bureau of Statistics. China Statistical Yearbook 2020; China Statistics Press: Beijing, China, 2020. (In Chinese)
- Zhang, X.; Dong, Z.; Gupta, H.; Wu, G.; Li, D. Impact of the three gorges dam on the hydrology and ecology of the yangtze river. Water 2016, 8, 590. [Google Scholar] [CrossRef]
- Chen, X.; He, D.; Zhou, L.; Cao, Y.; Li, Z. Influence of hydropower stations on the water microbiota in the downstream of Jinsha River, China. PeerJ 2020, 8, e9500. [Google Scholar] [CrossRef]
- Wu, J.; Huang, J.; Han, X.; Gao, X.; He, F.; Jiang, M.; Jiang, Z.; Primack, R.B.; Shen, Z. The Three Gorges Dam: An ecological perspective. Front. Ecol. Environ. 2004, 2, 241–248. [Google Scholar] [CrossRef]
- Galindo-Villegas, J.; García-Moreno, D.; de Oliveira, S.; Meseguer, J.; Mulero, V. Regulation of immunity and disease resistance by commensal microbes and chromatin modifications during zebrafish development. Proc. Natl. Acad. Sci. USA 2012, 109, 2605–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, T.; Balcázar, J.L.; Ruiz-Zarzuela, I.; Halaihel, N.; Vendrell, D.; de Blas, I.; Múzquiz, J.L. Host–microbiota interactions within the fish intestinal ecosystem. Mucosal Immunol. 2010, 3, 355–360. [Google Scholar] [CrossRef]
- Stagaman, K.; Burns, A.; Guillemin, K.; Bohannan, B.J. The role of adaptive immunity as an ecological filter on the gut microbiota in zebrafish. ISME J. 2017, 11, 1630–1639. [Google Scholar] [CrossRef]
- Xiong, J.; Nie, L.; Chen, J. Current understanding on the roles of gut microbiota in fish disease and immunity. Zool. Res. 2019, 40, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhou, L.; Yu, Y.; Ni, J.; Xu, W.; Yan, Q. Composition of gut microbiota in the gibel carp (Carassius auratus gibelio) varies with host development. Microb. Ecol. 2017, 74, 239–249. [Google Scholar] [CrossRef]
- Ni, J.; Yu, Y.; Zhang, T.; Gao, L. Comparison of intestinal bacterial communities in grass carp, Ctenopharyngodon idellus, from two different habitats. Chin. J. Oceanol. Limnol. 2012, 30, 757–765. [Google Scholar] [CrossRef]
- Ni, J.; Yan, Q.; Yu, Y.; Zhang, T. Factors influencing the grass carp gut microbiome and its effect on metabolism. FEMS Microbiol. Ecol. 2013, 87, 704–714. [Google Scholar] [CrossRef]
- Yan, Q.; Li, J.; Yu, Y.; Wang, J.; He, Z.; Van Nostrand, J.D.; Kempher, M.L.; Wu, L.; Wang, Y.; Liao, L.; et al. Environmental filtering decreases with fish development for the assembly of gut microbiota. Environ. Microbiol. 2016, 18, 4739–4754. [Google Scholar] [CrossRef]
- D’Aversa, F.; Tortora, A.; Ianiro, G.; Ponziani, F.R.; Annicchiarico, B.E.; Gasbarrini, A. Gut microbiota and metabolic syndrome. Intern. Emerg. Med. 2013, 8, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Ni, J.; Huang, R.; Zhou, H.; Xu, X.; Li, Y.; Cao, P.; Zhong, K.; Ge, M.; Chen, X.; Hou, B.; et al. Analysis of the relationship between the degree of dysbiosis in gut microbiota and prognosis at different stages of primary hepatocellular carcinoma. Front. Microbiol. 2019, 10, 1458. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Opperman, J.J.; Tickner, D.; Speed, R.; Guo, Q.; Chen, D. Managing the Three Gorges Dam to implement environmental flows in the Yangtze River. Front. Environ. Sci. 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.G.; Li, Z.J.; Liu, J.S.; Murphy, B.R. Fisheries of the Yangtze River show immediate impacts of the Three Gorges Dam. Fisheries 2007, 32, 343–344. [Google Scholar]
- Wu, H.; Yao, J.; Liu, Y.; Xue, J.; Cai, Q.; Liu, J. Seasonal variation and longitudinal distribution characters of cladocerans in the Three Gorges Reservoir. Biodiv. Sci. 2009, 17, 512–517. [Google Scholar]
- Näpflin, K.; Schmid-Hempel, P. Host effects on microbiota community assembly. J. Anim. Ecol. 2017, 87, 331–340. [Google Scholar] [CrossRef]
- Shao, K.; Yan, S.; Zhao, Y.; Xiong, M.; Xu, N.; Zhu, B.; Shi, F. Seventeen microsatellite loci isolated from Rhinogobio cylindricus (Gunther), and their cross-amplification in six Gobioninae species. Conserv. Genet. Resour. 2012, 5, 339–342. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Zhang, T. Life history strategy, trophic patterns and community structure of fishes in Biandantang Lake. Ph.D. Thesis, University of Chinese Academy of Sciences, Beijing, China, 2005. [Google Scholar]
- Ni, J.; Yu, Y.; Feng, W.; Yan, Q.; Pan, G.; Yang, B.; Xiang, Z.; Xuemei, L. Impacts of algal blooms removal by chitosan-modified soils on zoo-plankton community in Taihu Lake, China. J. Environ. Sci. 2010, 22, 1500–1507. [Google Scholar] [CrossRef]
- Huang, R.; Li, T.; Ni, J.; Bai, X.; Gao, Y.; Li, Y.; Zhang, P.; Gong, Y. Different Sex-Based Responses of Gut Microbiota During the Development of Hepatocellular Carcinoma in Liver-Specific Tsc1-Knockout Mice. Front. Microbiol. 2018, 9, 1008. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; He, T.; Wang, P.; Xie, M.; Xiang, J.; Ni, J. Opportunistic pathogens are abundant in the gut of cultured giant spiny frog (Paa spinosa). Aquac. Res. 2018, 49, 2033–2041. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Li, X.; He, Z.; Xu, M. A novel method to determine the minimum number of sequences required for reliable microbial community analysis. J. Microbiol. Methods 2017, 139, 196–201. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Parks, D.H.; Beiko, R.G. Identifying biologically relevant differences between metagenomic communities. Bioinformatics 2010, 26, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Knights, D.; Kuczynski, J.; Charlson, E.S.; Zaneveld, J.; Mozer, M.C.; Collman, R.G.; Bushman, F.D.; Knight, R.; Kelley, S.T. Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 2011, 8, 761–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, P.; Wang, J.-T.; Hu, H.-W.; Zheng, Y.-M.; Ge, Y.; Shen, J.-P.; He, J.-Z. Environmental filtering process has more important roles than dispersal limitation in shaping large-scale prokaryotic beta diversity patterns of grassland soils. Microb. Ecol. 2016, 72, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenlord, S.D.; Zak, D.R.; Upchurch, R.A. Dispersal limitation and the assembly of soil Actinobacteria communities in a long-term chronosequence. Ecol. Evol. 2012, 2, 538–549. [Google Scholar] [CrossRef]
- Logares, R.; Deutschmann, I.M.; Junger, P.C.; Giner, C.R.; Krabberød, A.K.; Schmidt, T.S.B.; Rubinat-Ripoll, L.; Mestre, M.; Salazar, G.; Ruiz-González, C. Disentangling the mechanisms shaping the surface ocean microbiota. Microbiome 2020, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Stone, R. China’s environmental challenges: Three Gorges Dam: Into the Unknown. Science 2008, 321, 628–632. [Google Scholar] [CrossRef]
- Cheng, F.; Li, W.; Castello, L.; Murphy, B.R.; Xie, S. Potential effects of dam cascade on fish: Lessons from the Yangtze River. Rev. Fish Biol. Fish. 2015, 25, 569–585. [Google Scholar] [CrossRef]
- Fu, C.; Wu, J.; Chen, J.; Wu, Q.; Lei, G. Freshwater fish biodiversity in the Yangtze River basin of China: Patterns, threats and conservation. Biodivers. Conserv. 2003, 12, 1649–1685. [Google Scholar] [CrossRef]
- Huang, F.; Xia, Z.; Zhang, N.; Lu, Z. Does hydrologic regime affect fish diversity? -A case study of the Yangtze Basin (China). Environ. Boil. Fishes 2011, 92, 569–584. [Google Scholar] [CrossRef]
- Xu, W.; Yang, Z.; Wan, L.; Tang, H.; Chen, X. Natural reproduction status of fish species producing pelagic eggs before and after impoundment of Yin-pan hydropower station in the lower Wujiang River. J. Hydroecol. 2019, 40, 8–15. [Google Scholar]
- Ruff, S.E.; Biddle, J.F.; Teske, A.P.; Knittel, K.; Boetius, A.; Ramette, A. Global dispersion and local diversification of the me-thane seep microbiome. Proc. Natl. Acad. Sci. USA 2015, 112, 4015–4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, Y.; Lindström, E.S.; Eiler, A.; Svanbäck, R. Different roles of environmental selection, dispersal, and drift in the assembly of intestinal microbial communities of freshwater fish with and without a stomach. Front. Ecol. Evol. 2020, 8, 152. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, H.; Tang, F.; Koide, R.; Cui, M.; Liu, Y.; Sun, Q.; Insam, H.; Zhang, Q. Relative roles of competition, environmental selection and spatial processes in structuring soil bacterial communities in the Qinghai-Tibetan Plateau. Appl. Soil Ecol. 2017, 117–118, 223–232. [Google Scholar] [CrossRef]
- Liu, Q.; Lai, Z.; Gao, Y.; Wang, C.; Zeng, Y.; Liu, E.; Mai, Y.; Yang, W.; Li, H. Connection between the gut microbiota of largemouth bass (Micropterus salmoides) and microbiota of the pond culture environment. Microorganisms 2021, 9, 1770. [Google Scholar] [CrossRef] [PubMed]
- Moeller, A.H.; Suzuki, T.A.; Lin, D.; Lacey, E.A.; Wasser, S.K.; Nachman, M.W. Dispersal limitation promotes the diversification of the mammalian gut microbiota. Proc. Natl. Acad. Sci. USA 2017, 114, 13768–13773. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Yan, Q.; Yu, Y.; Zhang, T. Fish gut microecosystem: A model for detecting spatial pattern of microorganisms. Chin. J. Oceanol. Limnol. 2013, 32, 54–57. [Google Scholar] [CrossRef] [Green Version]
- Pastor, H.J.B.; Gordon, D.M. Effects of dispersal limitation in the face of intense selection via dietary intervention on the faecal microbiota of rats. Environ. Microbiol. Rep. 2015, 8, 187–195. [Google Scholar] [CrossRef]
- Li, J.; Ni, J.; Wang, C.; Yu, Y.; Zhang, T. Different response patterns of fish foregut and hindgut microbiota to host habitats and genotypes. Appl. Ecol. Environ. Res. 2020, 18, 7369–7378. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Zhu, Q.; Yang, Z.; Sun, H.; Zhao, N.; Ni, J. Filtering Effect of Rhinogobio cylindricus Gut Microbiota Relieved Influence of the Three Gorges Dam on the Gut Microbiota Composition. Water 2021, 13, 2697. https://0-doi-org.brum.beds.ac.uk/10.3390/w13192697
Chen X, Zhu Q, Yang Z, Sun H, Zhao N, Ni J. Filtering Effect of Rhinogobio cylindricus Gut Microbiota Relieved Influence of the Three Gorges Dam on the Gut Microbiota Composition. Water. 2021; 13(19):2697. https://0-doi-org.brum.beds.ac.uk/10.3390/w13192697
Chicago/Turabian StyleChen, Xiaojuan, Qiguang Zhu, Zhi Yang, Hang Sun, Na Zhao, and Jiajia Ni. 2021. "Filtering Effect of Rhinogobio cylindricus Gut Microbiota Relieved Influence of the Three Gorges Dam on the Gut Microbiota Composition" Water 13, no. 19: 2697. https://0-doi-org.brum.beds.ac.uk/10.3390/w13192697