Wheat Germ Agglutinin—From Toxicity to Biomedical Applications


1
Institute of Pharmaceutical Technologies, Faculty of Pharmacy, Academy of Medicine, Lithuanian University of Health Sciences, 44307 Kaunas, Lithuania
2
Panevėžys Institute of Technologies and Business, Kaunas University of Technology, 44249 Kaunas, Lithuania
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(2), 884; https://0-doi-org.brum.beds.ac.uk/10.3390/app11020884
Submission received: 2 December 2020
/
Revised: 14 January 2021
/
Accepted: 15 January 2021
/
Published: 19 January 2021
(This article belongs to the Special Issue Advances in Food, Bioproducts and Natural Byproducts for a Sustainable Future: From Conventional to Innovative Processes)
Abstract
:Featured Application
Wheat germ agglutinin has the potential for enabling and improving targeted drug delivery systems, anticancer drugs, and antibacterial and antifungal therapeutics due to its cytotoxic mechanisms and specific carbohydrate binding.
Abstract
Wheat germ agglutinin is a hevein class N-Acetylglucosamine–binding protein with specific toxicity and biomedical potential. It is extractable from wheat germ—a low-value byproduct of the wheat industry—using well–established extraction methods based on salt precipitation and affinity chromatography. Due to its N-Acetylglucosamine affinity, wheat germ agglutinin exhibits antifungal properties as well as cytotoxic properties. Its anticancer properties have been demonstrated for various cancer cells, and toxicity mechanisms are well described. Wheat germ agglutinin has been demonstrated as a viable solution for various biomedical and therapeutic applications, such as chemotherapy, targeted drug delivery, antibiotic-resistant bacteria monitoring and elimination. This is performed mostly in conjunction with nanoparticles, liposomes, and other carrier mechanisms via surface functionalization. Combined with abundant wheat byproduct sources, wheat germ agglutinin has the potential to improve the biomedical field considerably.
Keywords:
wheat; germ; wheat byproducts; agglutinin; WGA; toxicity; glycosylation; N-Acetylglucosamine; GlcNAc; carbohydrate1. Introduction
Wheat (Triticum aestivum L.) is one of the most essential agricultural staple foods used for human consumption and animal feed. Approximately 21% of the world’s food supplies depend on annual wheat crop harvest [1], which causes the production of byproducts that are not always used efficiently or, in some cases, entirely discarded as waste [2]. Increasing trends of consumption-based economy restructuring to more ecologically-minded circular economy models encourage the scientific development of new technologies for byproduct valorization to create products with high nutritional value. One of the wheat processing byproducts which takes 2–3% of whole wheat grain [3] is wheat germ. They are a source of oils [4], tocopherols [5], various polyphenols [6], and specific proteins, such as agglutinins (lectins) [7]. Even though agglutinins are considered to be antinutrients [8], they might have various prospective applications in biomedicine, biotechnology, and agriculture itself. Considering the high quantities of wheat germ available as low-value byproducts throughout the world, any valorization attempts by extracting specific proteins may lead to high impact worldwide. This review is focused on wheat germ agglutinin, its structure and specificity, extraction and purification methods, biological activity, and possible applications.
2. Wheat Germ Agglutinin Structure
Wheat germ agglutinin (WGA) is one of the first purified lectins extracted at the very beginning of the lectinomics field. WGA structure was described during the 1970s. WGA is a mixture of three closely related major isoforms, named WGA1, WGA2, and WGA3 [9], which are a 36.0 kDa size stable 18.0 kDa polypeptide chain homodimer [10] with twofold axis symmetry [11]. Variability of these three isolectins is observed at 10 sequence positions, with WGA3 being the most distinct form, differing from WGA1 by 8 positions and from WGA2 by 7 positions [12]. The polypeptide chain is stable under high-temperature exposure [13]. Moreover, WGA monomers are highly resistant to acidity, and conformational changes can be reversed by increasing pH [14].
Each polypeptide chain is composed of four hevein domains, named A, B, C, and D [15,16] (Figure 1). WGA3 exhibits higher interdomain similarity than WGA1 or WGA2, suggesting closer relatedness to the common ancestral molecule [12]. Amino acid analysis shows that WGA contains high amounts of glycine and half-cystine, features not typical to most of the lectins [17]. Additionally, the protein is rich in disulfide bridges, with each hevein domain containing eight disulfide-forming cysteines [18]. These disulfides have been shown to play an important role in hevein stability since heveins lack a hydrophobic core [19]. Additionally, the disulfides seem to explain low pH value stability [14]. WGA has been shown to undergo cotranslational processing of glycan addition to the C-terminus, which is later post-translationally removed before WGA reaches mature form [20]. Affinity studies show that subunit specificity to oligosaccharides is much better than to monosaccharides. A, B, and C subunits bind N-Acetylglucosamine (GlcNAc) residues, such as chitin, whereas subunit D accommodates the glycoside aglycones. Monosaccharides are only bound to subunit C [21]. However, low affinity to other carbohydrates, such as N-Acetylneuraminic (sialic) acid, plays an important role in WGA activity [22]. Based on the structure and carbohydrate specificity, WGA is classified as chitin-binding lectin composed of hevein domains [23]. Lectin structure and carbohydrate specificity enable its broad spectrum of biological activities.
3. WGA Extraction and Purification
The most widely used natural WGA purification from wheat germ strategies include a series of protein precipitation and chromatographic purification steps. Untreated wheat germ material goes through a defatting step followed by material disruption with a laboratory mill. The material preparation is followed by extraction in water or aqueous buffers. Crude wheat extracts are used for protein precipitation with salts, mostly ammonium sulfate. Precipitated wheat proteins are resuspended and dialyzed. These steps are followed by chromatographic purification steps, including ion exchange and affinity chromatography. Table 1 represents summarized WGA purification steps and WGA yield (mg per 1 g of raw germ material).
4. WGA Biological Activity
4.1. Antifungal Activity
The main fungal cell wall component is chitin, which is a β(1,4)-homopolymer of GlcNAc. WGA specifically binds to fungal cell walls and disrupts the cell’s structural integrity [27]. It demonstrates WGA function in plant immunity against pathogenic fungi. However, there are not many results published regarding native WGA antifungal activity to human pathogenic fungi [28,29]. Recombinant WGA linked to the effector Fc region of murine IgG2a was designed for mycosis treatment. WGA-Fc antibodies were successfully bound to chitin standard fungal cultures and inhibited the growth of Histoplasma capsulatum, Cryptococcus neoformans, Candida albicans, and Saccharomyces cerevisiae. It also augmented antifungal macrophage functions and fully protected mice against H. capsulatum infections [29]. Other hevein class lectins demonstrate strong antifungal activity against Candida albicans (URM 5901), minimal inhibitory concentration (MIC) 25 μg/mL, against Candida krusei (URM 6391)–MIC 12.5 µg/mL [30] and Candida tropicalis (ATCC 750)–MIC 11.9 μg/mL [31].
4.2. Cytotoxicity
Since the early discovery of lectins, they were considered toxic due to the obvious toxicity of first-identified lectins, such as abrin [32] or ricin [33]. Even though only a few lectins are highly toxic in low doses, cytotoxic activity is typically dose-dependent. Lectin cytotoxicity can occur in a lectin-dependent cell-mediated manner or as direct toxicity to cells [34]. Lectins interact with immune cells as antigens and exhibit their cytotoxic activities and induce a humoral response. Concentration-dependent increases with IFN-γ, and TNF-α release were observed after whole blood samples were treated with WGA [35]. Another study has found that WGA binds to glycosylated T3 receptor, which is followed by the formation of inositol phosphate—an intracellular secondary messenger that takes part in the regulation of cell functions, such as apoptosis—in human blood T cells [36]. On the other hand, WGA inhibits T lymphocyte proliferation by inhibiting the responsiveness of the lymphocytes to interleukin 2 (IL-2) rather than interfering with IL-2 production and IL-2 receptor expression [37,38].
Direct lectin cytotoxicity depends on interaction time and affinity to glycoconjugates on the cell surface. WGA demonstrates high specificity to GlcNAc, but interaction with sialic acid is important for WGA cell-killing activity. WGA-induced acute myeloid leukemia cell death is dependent on both GlcNAc binding and interaction with sialic acids. The authors noticed a reduction in WGA cell binding and killing after sialic acid was removed by neuraminidase treatment [39]. Sialic acid specificity plays a crucial role because most of the cancer cells are hyper sialylated [40,41]. Experiments show that WGA is endocytosed into the Chinese hamster ovary (CHO) cells after 30 min of treatment [42]. Later research confirmed the same time-dependent endocytosis with leukemic Jurkat cell line [43]. WGA is endocytosed into cells by clathrin-independent endocytosis mechanism depending on interactions with N-acetylneuraminic acid or N-acetylglucosamine (GlcNAc) on the cell surface of normal and malignant cells [44,45]. WGA binds high molecular weight glycoproteins (HMWAG) expressed on the cell surface. WGA-HMWAG complexes rapidly become localized within endosomes and then move more slowly to a tubular endocytic network within the perinuclear region of the cell [42]. Endocytosed WGA interacts with the cell nuclear envelope pore membrane protein POM 121 and blocks RNA export [46]. This interaction with nuclear pore proteins depends on GlcNAc glycosylation moieties in other cell models of the nuclear transport blockade [47,48]. Endocytosed WGA can suppress rat pancreatic acinar tumor cell line proliferation by blocking auto stimulation and prevention of the trophic effect of glycosylated cholecystokinin or gastrin–like peptides [49].
WGA is shown to exert cytotoxic effects to differing extents with a final outcome (apoptosis or necrosis) depending on the cell line used in toxicity assays. WGA induced paraptosis in L929 murine fibroblast cell line. Vacuolation, loss of cell architecture, and upregulation of the apoptosis-related proteins Bax and caspase-3 were observed. However, internucleosomal DNA fragmentation, apoptotic bodies, and related apoptotic phenotypes were not detected [50]. Meanwhile, WGA intake into pancreatic carcinoma cell lines caused chromatin condensation, nuclear fragmentation, and DNA release consistent with apoptosis [51]. Interaction with the leukemic Jurkat cell line induced a loss of transmembrane potential, disruption of the inner mitochondrial membrane, and release of cytochrome c and caspase-9 activation of apoptosis [43]. These variances of outcomes might also be caused by differences between cell glycosylation patterns [52].
5. WGA Biomedical Perspectives
5.1. Advanced Drug Delivery
Despite the WGA toxicity, glycosylation-targeted reactivity with biological molecules might have great potential for WGA application in modern biomedicine. Recent studies focus on the improvement of drug delivery systems. WGA–anchored nanoparticles improved nanoparticle adhesion onto cells [53]. Paclitaxel-loaded nanoparticles with immobilized WGA molecules demonstrated higher cytotoxicity to cancer cell lines than the paclitaxel alone [54,55]. This efficacy improvement could be attributed to a more efficient cellular uptake via WGA-receptor-mediated endocytosis and isopropyl myristate-facilitated release of paclitaxel from the nanoparticles. This efficacy enhancement is observed with other chemotherapeutic agents, too, such as. docetaxel [56] or doxorubicin [57].
Biotechnologically synthesized macromolecular drugs are considered to be the future of the pharmacy. Good response to these drugs is limited due to poor absorption and high susceptibility to the loss of their activity. WGA as a polypeptide delivery mediator was tested with a fluorescein-labeled (F) bull serum albumin (BSA) complex on the Caco-2 cell line. Results indicated up to 8.7 times higher F-BSA/WGA complex binding to Caco-2 cells as compared with glycyl-F-BSA complex alone. Moreover, about 75% of F-BSA-WGA were bound specifically to Caco-2 cells [58]. More complex nanoparticles (liposomes) with a polymer–WGA surface conjugate led to an increase in the association between liposomes and lung epithelial cells enhanced systemic absorption of calcitonin model peptide improved bio adhesion to lung epithelia also significantly enhanced and prolonged the therapeutic efficacy of calcitonin [59]. WGA-functionalized carboxymethylated kappa–carrageenan microparticles with entrapped insulin molecules were protected from hydrolysis and proteolysis by stomach acids and enzymes. Microparticle interactions with intestinal wall cells and insulin absorption were improved by WGA grafts [60]. Technologically advanced systems with functionalized lectins demonstrate higher lectin efficacy than standalone lectins. Magnetic nanoparticles with immobilized WGA increased apoptosis in prostate cancer cells in vitro and in vivo in the presence of a magnetic field [61]. Lectin interaction with cell glycosylation points facilitated the internalization of chemotherapeutic compounds and magnetic particles, which results in higher targeted efficacy in cancer treatment. WGA was also used to develop a method for photodynamic inactivation of methicillin-resistant S. aureus and Pseudomonas aeruginosa [62]. WGA-modified liposomes loaded with generation II photosensitizers exhibited considerably higher photosensitizers deliverance to the bacteria than the unmodified liposomes, demonstrating WGA–liposome complex use not only for antibiotic-resistant bacteria detection but also elimination. Table 2 summarizes various developed drug delivery systems utilizing WGA.
5.2. Overcoming Physiological Barriers
Drug delivery usually involves crossing specific biological barriers. Nonspecific distribution and inadequate accumulation of therapeutics remain formidable challenges for drug developers. The blood-brain barrier (BBB) might also be overcome with WGA incorporation on nanoparticles [63,64,65]. PEGylated (Polyethylene glycol modified) fourth-generation poly(amidoamine) (PAMAM) dendrimer-based nanocarrier containing doxorubicin (DOX) inside and WGA with transferrin (Tf) as targeting molecules were developed for BBB crossing. A PA-MAM–WGA–Tf nanocarrier increased DOX delivery through BBB by 13.5% compared to free doxorubicin. Experiments also showed increased DOX accumulation in the tumor site due to dual targeting mechanisms. Accumulation led to the complete breakage of the avascular C6 glioma spheroids in vitro [66]. Recent studies suggest a novel intranasal nanoparticle delivery strategy to overcome BBB. WGA-modified PEG-poly(lactic acid) (PLA) nanoparticles with miR132 were administered to rodents. Transmission electron microscopy results demonstrated that the accumulation of WGA–modified nanoparticles in the brain was significantly higher than that of unmodified nanoparticles. Moreover, the results of Morris water maze analysis showed that the intranasal delivery of WGA–PLA–miR132 improved the learning and memory function of APP/PS1 mice [67]. These results suggest novel drug delivery through BBB strategies, which might overcome treatment-resistant intracranial tumors and neurodegenerative diseases, such as Alzheimer’s disease.
WGA-incorporated nanoparticles also demonstrate promising features for targeting drugs to bladder cancer cells. Due to the urothelial barrier, constant active drug substance washout, and short dwelling time, bladder cancers and urinary tract infections are another difficulty for efficient drug delivery. Surgical tumor resection is usually followed by repeated intravesical chemotherapy. WGA alone demonstrates high binding to 5637 bladder cancer cells [68]. Later investigations in flow-cytometric and fluorometric experiments with single cells and cell monolayers showed fluorescein (Fc)-labeled poly-l-glutamic acid (PGA)–WGA drug delivery system binding and the internalization of high molecular weight (>100 kDa) conjugates. Results of specificity studies showed that the interaction between the Fc–PGA–WGA conjugates and the cell surface depended solely on the WGA component [69]. A recent study by Brauner and colleagues demonstrated poly(d,l-lactic-co-glycolic acid) (PLGA)–WGA nanoparticles’ inherent adhesion capability to immortalized human uroepithelial cells. A slight adhesion capacity increase was WGA biospecificity-dependant [70].
WGA–liposome–cyclodextrin complexes were bioengineered for oral cell therapeutics [71]. The designed carrier showed fast attachment to oral cells and sustained co-drug release of ciprofloxacin and betamethasone in saliva in vitro. The complex significantly increased oral cell survival against Aggregatibacter actinomycetemcomitans, a bacterial pathogen responsible for chronic periodontal disease, and simultaneously reduced the inflammation. Crucially, WGA-facilitated cell-binding prevented complex degradation and loss due to salivary flushing and/or mucus turnover and allowed for sustained 24 h drug release.
These promising results might eventually bring some breakthroughs in medicine. However, in vitro and some in vivo results with animal models still have to go through in-depth clinical trials before advanced WGA-facilitated drug delivery systems can be applied in clinics.
6. Non-Medical WGA Applications
Outside of medical applications, WGA potential has been demonstrated as an option for basic research and biotechnological use. Quantum dot–WGA complexes have been used for elucidating the endocytic and exocytic lectin processes [72]. Single-particle tracking techniques of these complexes showed that WGA processes are both actin- and microtubule-dependent, and each contains a five-stage transport route that is transferable to design processes of any lectins as drug carriers or antineoplastic drugs.
Anisotropic silver nanoparticles functionalized with WGA or Lens Culinaris agglutinin have been demonstrated to detect specific Gram-positive and Gram-negative bacteria [73]. The designed photometric assay could detect S. aureus down to 103 cells/mL and E. Coli down to 3 × 103 cells/mL by having the bacteria-bound nanoparticle–lectin compounds resilient to NaCl-induced agglomeration. The assay was demonstrated on urine samples as well.
Extraction of two recombinant human glycoproteins—erythropoietin and Darbepoetin—from equine plasma by immunoaffinity chromatography has been shown to be improved by 15% when pretreating the samples with wheat germ agglutinin immobilized on Sepharose gel [74]. The technique is easily scalable and indicates the WGA potential for naturally or synthetically glycosylated recombinant protein industry.
The various biotechnological applications are already demonstrating the WGA value.
7. Conclusions
A 36 kDa size, hevein class, GlcNAc-specific lectin WGA is considered an antinutrient compound. Carbohydrate specificity enables its biological activities, such as fungistatic, cytotoxic, pro-apoptotic, and pro-necrotic. WGA might modulate immune cell response to antigens and cause changes in immune cell signaling. With regard to biotechnological applications, WGA is a highly promising tool in nanotechnology and advanced drug delivery systems with examples WGA-based nanoparticle, liposome, and other complexes for production, research, bacteria monitoring, and elimination. Considering the large quantities of wheat germ produced worldwide as a waste or low value byproduct, WGA extraction and utilization might be a viable solution for relatively cheap biomedical improvements in the future.
Author Contributions
Conceptualization, G.B.-M.; investigation, G.B.-M. and M.D.; writing—original draft preparation, G.B.-M. and M.D.; writing—review and editing, G.B.-M. and M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Regional Development Fund according to the supported activity ‘Attracting scientists from abroad to carry out research’ under Measure No. 01.2.2-LMT-K-718 (project No. 01.2.2-LMT-K-718-02-0012).
Acknowledgments
Authors also acknowledge Panevėžys Mechatronics Center for providing workspace, literature, and the technical base.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Enghiad, A.; Ufer, D.; Countryman, A.M.; Thilmany, D.D. An Overview of Global Wheat Market Fundamentals in an Era of Climate Concerns. Available online: https://www.hindawi.com/journals/ija/2017/3931897/ (accessed on 1 April 2020).
- Bacenetti, J.; Duca, D.; Negri, M.; Fusi, A.; Fiala, M. Mitigation Strategies in the Agro-Food Sector: The Anaerobic Digestion of Tomato Purée by-Products. An Italian Case Study. Sci. Total Environ. 2015, 526, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Šramková, Z.; Gregová, E.; Šturdík, E. Chemical Composition and Nutritional Quality of Wheat Grain. Acta Chim. Slovaca 2009, 2, 115–138. [Google Scholar]
- Edwin Geo, V.; Prabhu, C.; Thiyagarajan, S.; Maiyalagan, T.; Aloui, F. Comparative Analysis of Various Techniques to Improve the Performance of Novel Wheat Germ Oil—An Experimental Study. Int. J. Hydrog. Energy 2020, 45, 5745–5756. [Google Scholar] [CrossRef]
- Boukid, F.; Folloni, S.; Ranieri, R.; Vittadini, E. A Compendium of Wheat Germ: Separation, Stabilization and Food Applications. Trends Food Sci. Technol. 2018, 78, 120–133. [Google Scholar] [CrossRef]
- Teslić, N.; Bojanić, N.; Rakić, D.; Takači, A.; Zeković, Z.; Fišteš, A.; Bodroža-Solarov, M.; Pavlić, B. Defatted Wheat Germ as Source of Polyphenols—Optimization of Microwave-Assisted Extraction by RSM and ANN Approach. Chem. Eng. Process.-Process Intensif. 2019, 143, 107634. [Google Scholar] [CrossRef]
- Schwefel, D.; Maierhofer, C.; Beck, J.G.; Seeberger, S.; Diederichs, K.; Möller, H.M.; Welte, W.; Wittmann, V. Structural Basis of Multivalent Binding to Wheat Germ Agglutinin. J. Am. Chem. Soc. 2010, 132, 8704–8719. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Arntfield, S.D.; Nickerson, M. Changes in Levels of Phytic Acid, Lectins and Oxalates during Soaking and Cooking of Canadian Pulses. Food Res. Int. 2018, 107, 660–668. [Google Scholar] [CrossRef]
- Smith, J.J.; Raikhel, N.V. Nucleotide Sequences of CDNA Clones Encoding Wheat Germ Agglutinin Isolectins A and D. Plant Mol. Biol. 1989, 13, 601–603. [Google Scholar] [CrossRef]
- Peumans, W.J.; Stinissen, H.M.; Carlier, A.R. Isolation and Partial Characterization of Wheat-Germ-Agglutinin-like Lectins from Rye (Secale Cereale) and Barley (Hordeum Vulgare) Embryos. Biochem. J. 1982, 203, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Rice, R.H.; Etzler, M.E. Subunit Structure of Wheat Germ Agglutinin. Biochem. Biophys. Res. Commun. 1974, 59, 414–419. [Google Scholar] [CrossRef]
- Wright, C.S.; Raikhel, N. Sequence Variability in Three Wheat Germ Agglutinin Isolectins: Products of Multiple Genes in Polyploid Wheat. J. Mol. Evol. 1989, 28, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Matucci, A.; Veneri, G.; Dalla Pellegrina, C.; Zoccatelli, G.; Vincenzi, S.; Chignola, R.; Peruffo, A.D.B.; Rizzi, C. Temperature-Dependent Decay of Wheat Germ Agglutinin Activity and Its Implications for Food Processing and Analysis. Food Control 2004, 15, 391–395. [Google Scholar] [CrossRef]
- Portillo-Téllez, M.D.C.; Bello, M.; Salcedo, G.; Gutiérrez, G.; Gómez-Vidales, V.; García-Hernández, E. Folding and Homodimerization of Wheat Germ Agglutinin. Biophys. J. 2011, 101, 1423–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Romero, A.; Ravichandran, K.G.; Soriano-García, M. Crystal Structure of Hevein at 2.8 Å Resolution. Febs. Lett. 1991, 291, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Leyva, E.; Medrano-Cerano, J.L.; Cano-Sánchez, P.; López-González, I.; Gómez-Velasco, H.; Del Río-Portilla, F.; García-Hernández, E. Bacterial Expression, Purification and Biophysical Characterization of Wheat Germ Agglutinin and Its Four Hevein-like Domains. Biopolymers 2019, 110, e23242. [Google Scholar] [CrossRef]
- Nagata, Y.; Goldberg, A.R.; Burger, M.M. The isolation and purification of wheat germ and other agglutinins. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1974; Volume 32, pp. 611–615. ISBN 978-0-12-181895-1. [Google Scholar]
- Wright, H.T.; Sandrasegaram, G.; Wright, C.S. Evolution of a Family of N-Acetylglucosamine Binding Proteins Containing the Disulfide-Rich Domain of Wheat Germ Agglutinin. J. Mol. Evol. 1991, 33, 283–294. [Google Scholar] [CrossRef]
- Hernández-Arana, A.; Rojo-Domínguez, A.; Soriano-García, M.; Rodríguez-Romero, A. The Thermal Unfolding of Hevein, a Small Disulfide-Rich Protein. Eur. J. Biochem. 1995, 228, 649–652. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Peumans, W.J.; Raikhel, N.V. Wheat-Germ Agglutinin Is Synthesized as a Glycosylated Precursor. Planta 1988, 173, 482–489. [Google Scholar] [CrossRef]
- Allen, A.K.; Neuberger, A.; Sharon, N. The Purification, Composition and Specificity of Wheat-Germ Agglutinin. Biochem. J. 1973, 131, 155–162. [Google Scholar] [CrossRef]
- Gallagher, J.T.; Morris, A.; Dexter, T.M. Identification of Two Binding Sites for Wheat-Germ Agglutinin on Polylactosamine-Type Oligosaccharides. Biochem. J. 1985, 231, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Peumans, W.J.; van Damme, E.J.M.; Barre, A.; Rougé, P. Classification of Plant Lectins in Families Of Structurally and Evolutionary Related Proteins. Adv. Exp. Med. Biol. 2001, 491, 27–54. [Google Scholar] [CrossRef] [PubMed]
- Harata, K.; Nagahora, H.; Jigami, Y. X-ray Structure of Wheat Germ Agglutinin Isolectin 3. Acta Cryst. D 1995, 51, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Lotan, R.; Gussin, A.E.S.; Lis, H.; Sharon, N. Purification of Wheat Germ Agglutinin by Affinity Chromatography on a Sepharose-Bound N-Acetylglucosamine Derivative. Biochem. Biophys. Res. Commun. 1973, 52, 656–662. [Google Scholar] [CrossRef]
- Baieli, M.F.; Urtasun, N.; Miranda, M.V.; Cascone, O.; Wolman, F.J. Efficient Wheat Germ Agglutinin Purification with a Chitosan-Based Affinity Chromatographic Matrix. J. Sep. Sci. 2012, 35, 231–238. [Google Scholar] [CrossRef]
- Ciopraga, J.; Gozia, O.; Tudor, R.; Brezuica, L.; Doyle, R.J. Fusarium Sp. Growth Inhibition by Wheat Germ Agglutinin. Biochim. Biophys. Acta 1999, 1428, 424–432. [Google Scholar] [CrossRef]
- Tonkal, A.M. In Vitro Antitrichomonal Effect of Nigella Sativa Aqueous Extract and Wheat Germ Agglutinin. J. King Abdulaziz Univ. Med. Sci. 2009, 16, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Liedke, S.C.; Miranda, D.Z.; Gomes, K.X.; Gonçalves, J.L.S.; Frases, S.; Nosanchuk, J.D.; Rodrigues, M.L.; Nimrichter, L.; Peralta, J.M.; Guimarães, A.J. Characterization of the Antifungal Functions of a WGA-Fc (IgG2a) Fusion Protein Binding to Cell Wall Chitin Oligomers. Sci. Rep. 2017, 7, 12187. [Google Scholar] [CrossRef]
- da Silva, P.M.; de Moura, M.C.; Gomes, F.S.; da Silva Trentin, D.; Silva de Oliveira, A.P.; de Mello, G.S.V.; da Rocha Pitta, M.G.; de Melo Rego, M.J.B.; Coelho, L.C.B.B.; Macedo, A.J.; et al. PgTeL, the Lectin Found in Punica Granatum Juice, Is an Antifungal Agent against Candida Albicans and Candida Krusei. Int. J. Biol. Macromol. 2018, 108, 391–400. [Google Scholar] [CrossRef]
- Kanokwiroon, K.; Teanpaisan, R.; Wititsuwannakul, D.; Hooper, A.B.; Wititsuwannakul, R. Antimicrobial Activity of a Protein Purified from the Latex of Hevea Brasiliensis on Oral Microorganisms. Mycoses 2008, 51, 301–307. [Google Scholar] [CrossRef]
- Dickers, K.J.; Bradberry, S.M.; Rice, P.; Griffiths, G.D.; Vale, J.A. Abrin Poisoning. Toxicol. Rev. 2003, 22, 137–142. [Google Scholar] [CrossRef]
- Lopez Nunez, O.F.; Pizon, A.F.; Tamama, K. Ricin Poisoning after Oral Ingestion of Castor Beans: A Case Report and Review of the Literature and Laboratory Testing. J. Emerg. Med. 2017, 53, e67–e71. [Google Scholar] [CrossRef]
- Gorelik, E. Mechanisms of Cytotoxic Activity of Lectins. Trends Glycosci. Glycotechnol. 1994, 6, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Fraser, S.; Sadofsky, L.; Hart, S. Peripheral Blood Leukocyte Immune Responses Are Distinctly Altered in Sarcoidosis. Eur. Respir. J. 2015, 46. [Google Scholar] [CrossRef]
- Clevers, H.C.; de Bresser, A.; Kleinveld, H.; Gmelig-Meyling, F.H.; Ballieux, R.E. Wheat Germ Agglutinin Activates Human T Lymphocytes by Stimulation of Phosphoinositide Hydrolysis. J. Immunol. 1986, 136, 3180–3183. [Google Scholar] [PubMed]
- Kawakami, K.; Yamamoto, Y.; Onoue, K. Effect of Wheat Germ Agglutinin on T Lymphocyte Activation. Microbiol. Immunol. 1988, 32, 413–422. [Google Scholar] [CrossRef]
- Reed, J.C.; Robb, R.J.; Greene, W.C.; Nowell, P.C. Effect of Wheat Germ Agglutinin on the Interleukin Pathway of Human T Lymphocyte Activation. J. Immunol. 1985, 134, 314–323. [Google Scholar]
- Ryva, B.; Zhang, K.; Asthana, A.; Wong, D.; Vicioso, Y.; Parameswaran, R. Wheat Germ Agglutinin as a Potential Therapeutic Agent for Leukemia. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Boligan, K.F.; Mesa, C.; Fernandez, L.E.; von Gunten, S. Cancer Intelligence Acquired (CIA): Tumor Glycosylation and Sialylation Codes Dismantling Antitumor Defense. Cell. Mol. Life Sci. 2015, 72, 1231–1248. [Google Scholar] [CrossRef]
- Pearce, O.M.T.; Läubli, H. Sialic Acids in Cancer Biology and Immunity. Glycobiology 2016, 26, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Raub, T.J.; Koroly, M.J.; Roberts, R.M. Endocytosis of Wheat Germ Agglutinin Binding Sites from the Cell Surface into a Tubular Endosomal Network. J. Cell. Physiol. 1990, 143, 1–12. [Google Scholar] [CrossRef]
- Gastman, B.; Wang, K.; Han, J.; Zhu, Z.; Huang, X.; Wang, G.-Q.; Rabinowich, H.; Gorelik, E. A Novel Apoptotic Pathway as Defined by Lectin Cellular Initiation. Biochem. Biophys. Res. Commun. 2004, 316, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Wirth, C.; Schwuchow, J.; Jonas, L. Internalization of Wheat Germ Agglutinin (WGA) by Rat Pancreatic Cells in Vivo and in Vitro. Acta Histochem. 1996, 98, 165–172. [Google Scholar] [CrossRef]
- Pellegrina, C.D.; Matucci, A.; Zoccatelli, G.; Rizzi, C.; Vincenzi, S.; Veneri, G.; Andrighetto, G.; Peruffo, A.D.B.; Chignola, R. Studies on the Joint Cytotoxicity of Wheat Germ Agglutinin and Monensin. Toxicol. Vitr. 2004, 18, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, E.; Wozniak, R.; Blobel, G. An Integral Membrane Protein of the Pore Membrane Domain of the Nuclear Envelope Contains a Nucleoporin-like Region. J. Cell. Biol. 1993, 122, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W.; Hanover, J.A. Functional Nuclear Pores Reconstituted with Beta 1-4 Galactose-Modified O-Linked N-Acetylglucosamine Glycoproteins. J. Biol. Chem. 1994, 269, 9289–9297. [Google Scholar] [CrossRef]
- Heese-Peck, A.; Cole, R.N.; Borkhsenious, O.N.; Hart, G.W.; Raikhel, N.V. Plant Nuclear Pore Complex Proteins Are Modified by Novel Oligosaccharides with Terminal N-Acetylglucosamine. Plant Cell 1995, 7, 1459–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, C.; Nebe, B.; Walzel, H.; Weber, H.; Jonas, L. Inhibitory Effect of the Lectin Wheat Germ Agglutinin (WGA) on the Proliferation of AR42J Cells. Acta Histochem. 2009, 111, 336–343. [Google Scholar] [CrossRef]
- Tsai, T.L.; Wang, H.C.; Hung, C.H.; Lin, P.C.; Lee, Y.S.; Chen, H.H.W.; Su, W.C. Wheat Germ Agglutinin-Induced Paraptosis-like Cell Death and Protective Autophagy Is Mediated by Autophagy-Linked FYVE Inhibition. Oncotarget 2017, 8, 91209–91222. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, R.E.; Wojciechowicz, D.C.; Picon, A.I.; Schwarz, M.A.; Paty, P.B. Wheatgerm Agglutinin-Mediated Toxicity in Pancreatic Cancer Cells. Br. J. Cancer 1999, 80, 1754–1762. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Seigner, C.; Bastid, C.; Choux, R.; Payan, M.J.; Reggio, H. Carcinoembryonic Antigen Has a Different Molecular Weight in Normal Colon and in Cancer Cells Due to N-Glycosylation Differences. Cancer Res. 1991, 51, 5679–5686. [Google Scholar]
- Weissenböck, A.; Wirth, M.; Gabor, F. WGA-Grafted PLGA-Nanospheres: Preparation and Association with Caco-2 Single Cells. J. Control. Release 2004, 99, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Lim, L.-Y. Preparation and in Vitro Anticancer Activity of Wheat Germ Agglutinin (WGA)-Conjugated PLGA Nanoparticles Loaded with Paclitaxel and Isopropyl Myristate. J. Control. Release 2005, 107, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ho, P.C.; Lim, L.Y. Wheat Germ Agglutinin-Conjugated PLGA Nanoparticles for Enhanced Intracellular Delivery of Paclitaxel to Colon Cancer Cells. Int. J. Pharm. 2010, 400, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.I.; Ayub, A.D.; Mat Yusuf, S.N.A.; Yahaya, N.; Abd Kadir, E.; Lim, V. Docetaxel-Loaded Disulfide Cross-Linked Nanoparticles Derived from Thiolated Sodium Alginate for Colon Cancer Drug Delivery. Pharmaceutics 2020, 12, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.-C.; Chang, Y.-H.; Rajesh, R. Targeted Delivery of Etoposide, Carmustine and Doxorubicin to Human Glioblastoma Cells Using Methoxy Poly(Ethylene Glycol)-poly(Ε-caprolactone) Nanoparticles Conjugated with Wheat Germ Agglutinin and Folic Acid. Mater. Sci. Eng. C 2019, 96, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Gabor, F.; Schwarzbauer, A.; Wirth, M. Lectin-Mediated Drug Delivery: Binding and Uptake of BSA-WGA Conjugates Using the Caco-2 Model. Int. J. Pharm. 2002, 237, 227–239. [Google Scholar] [CrossRef]
- Murata, M.; Yonamine, T.; Tanaka, S.; Tahara, K.; Tozuka, Y.; Takeuchi, H. Surface Modification of Liposomes Using Polymer-Wheat Germ Agglutinin Conjugates to Improve the Absorption of Peptide Drugs by Pulmonary Administration. J. Pharm. Sci. 2013, 102, 1281–1289. [Google Scholar] [CrossRef]
- Leong, K.H.; Chung, L.Y.; Noordin, M.I.; Onuki, Y.; Morishita, M.; Takayama, K. Lectin-Functionalized Carboxymethylated Kappa-Carrageenan Microparticles for Oral Insulin Delivery. Carbohydr. Polym. 2011, 86, 555–565. [Google Scholar] [CrossRef]
- AlSadek, D.M.M.; Badr, H.A.; Al-Shafie, T.A.; El-Bahr, S.M.; El-Houseini, M.E.; Djansugurova, L.B.; Li, C.-Z.; Ahmed, H. Cancer Cell Death Induced by Nanomagnetolectin. Eur. J. Cell Biol. 2017, 96, 600–611. [Google Scholar] [CrossRef]
- Yang, K.; Gitter, B.; Rüger, R.; Albrecht, V.; Wieland, G.D.; Fahr, A. Wheat Germ Agglutinin Modified Liposomes for the Photodynamic Inactivation of Bacteria†. Photochem. Photobiol. 2012, 88, 548–556. [Google Scholar] [CrossRef]
- Gao, X.; Wu, B.; Zhang, Q.; Chen, J.; Zhu, J.; Zhang, W.; Rong, Z.; Chen, H.; Jiang, X. Brain Delivery of Vasoactive Intestinal Peptide Enhanced with the Nanoparticles Conjugated with Wheat Germ Agglutinin Following Intranasal Administration. J. Control. Release 2007, 121, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.; Xiao, C.; Davis, F.; Lambert, M.; Viola, K.; Lacor, P.; Dravid, V.; Klein, W. Intranasal Passive Immunization Using WGA-Modified Oligomer Antibodies Greatly Improves Learning and Memory in Alzheimer’s 5XFAD Mice. Alzheimer’s Dement. 2012, 8, P197. [Google Scholar] [CrossRef]
- Zhang, Y.; Walker, J.B.; Minic, Z.; Liu, F.; Goshgarian, H.; Mao, G. Transporter Protein and Drug-Conjugated Gold Nanoparticles Capable of Bypassing the Blood-Brain Barrier. Sci. Rep. 2016, 6, 25794. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, Y.; Jia, X.-R.; Du, J.; Ying, X.; Lu, W.-L.; Lou, J.-N.; Wei, Y. PEGylated Poly(Amidoamine) Dendrimer-Based Dual-Targeting Carrier for Treating Brain Tumors. Biomaterials 2011, 32, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Sun, B.; Gao, X.; Dong, X.; Fu, L.; Zhang, Y.; Li, Z.; Wang, Y.; Jiang, H.; Han, B. Intranasal Delivery of Targeted Nanoparticles Loaded With MiR-132 to Brain for the Treatment of Neurodegenerative Diseases. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Plattner, V.E.; Wagner, M.; Ratzinger, G.; Gabor, F.; Wirth, M. Targeted Drug Delivery: Binding and Uptake of Plant Lectins Using Human 5637 Bladder Cancer Cells. Eur. J. Pharm. Biopharm. 2008, 70, 572–576. [Google Scholar] [CrossRef]
- Apfelthaler, C.; Anzengruber, M.; Gabor, F.; Wirth, M. Poly-(l)-Glutamic Acid Drug Delivery System for the Intravesical Therapy of Bladder Cancer Using WGA as Targeting Moiety. Eur. J. Pharm. Biopharm. 2017, 115, 131–139. [Google Scholar] [CrossRef]
- Brauner, B.; Semmler, J.; Rauch, D.; Nokaj, M.; Haiss, P.; Schwarz, P.; Wirth, M.; Gabor, F. Trimethoprim-Loaded PLGA Nanoparticles Grafted with WGA as Potential Intravesical Therapy of Urinary Tract Infections—Studies on Adhesion to SV-HUCs Under Varying Time, PH, and Drug-Loading Conditions. Acs Omega 2020, 5, 17377–17384. [Google Scholar] [CrossRef]
- Wijetunge, S.S.; Wen, J.; Yeh, C.-K.; Sun, Y. Wheat Germ Agglutinin Liposomes with Surface Grafted Cyclodextrins as Bioadhesive Dual-Drug Delivery Nanocarriers to Treat Oral Cells. Colloids Surf. B Biointerfaces 2020, 185, 110572. [Google Scholar] [CrossRef]
- Liu, S.-L.; Zhang, Z.-L.; Sun, E.-Z.; Peng, J.; Xie, M.; Tian, Z.-Q.; Lin, Y.; Pang, D.-W. Visualizing the Endocytic and Exocytic Processes of Wheat Germ Agglutinin by Quantum Dot-Based Single-Particle Tracking. Biomaterials 2011, 32, 7616–7624. [Google Scholar] [CrossRef]
- Mikaelyan, M.V.; Poghosyan, G.G.; Hendrickson, O.D.; Dzantiev, B.B.; Gasparyan, V.K. Wheat Germ Agglutinin and Lens Culinaris Agglutinin Sensitized Anisotropic Silver Nanoparticles in Detection of Bacteria: A Simple Photometric Assay. Anal. Chim. Acta 2017, 981, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Stanley, S.M.R.; Chua, D. Improved Recovery of Erythropoietin and Darbepoetin from Equine Plasma by the Application of a Wheat Germ Agglutinin Mediated Pre-Extraction Prior to Immunoaffinity Chromatography. Adv. Biosci. Biotechnol. 2014. [Google Scholar] [CrossRef] [Green Version]
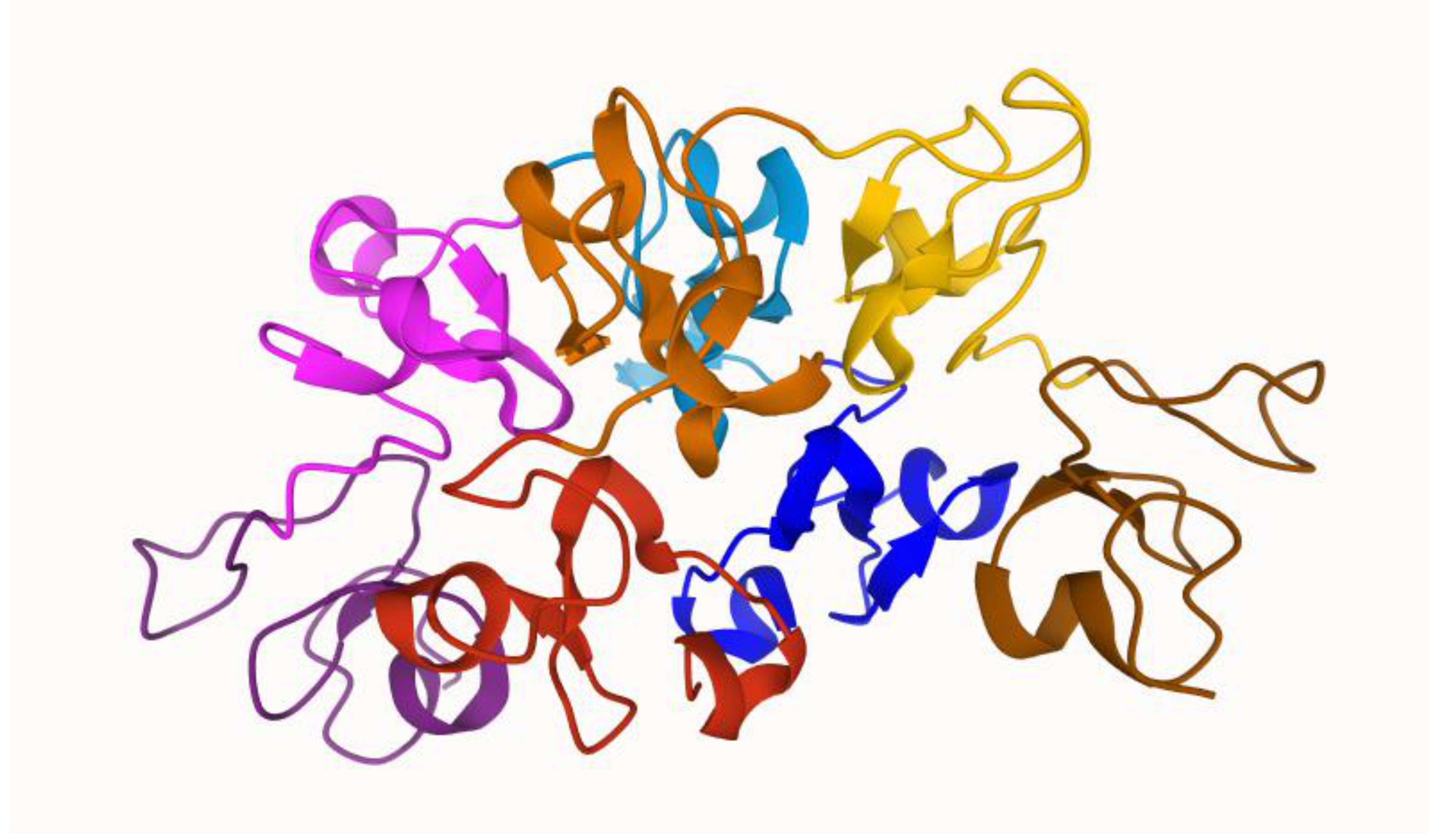
Figure 1.
Dimeric wheat germ agglutinin 3 (WGA3) structure by [24]. Red, orange, yellow, and brown colors represent the subunits A, B, C, and D of the first protein, respectively. Blue, light blue, pink, and purple represent the subunits A, B, C, and D of the second protein, respectively. The sugar-binding site is located at the interface of both WGA monomers.
Figure 1.
Dimeric wheat germ agglutinin 3 (WGA3) structure by [24]. Red, orange, yellow, and brown colors represent the subunits A, B, C, and D of the first protein, respectively. Blue, light blue, pink, and purple represent the subunits A, B, C, and D of the second protein, respectively. The sugar-binding site is located at the interface of both WGA monomers.


{kind=link}
Table 1.
Wheat germ agglutinin (WGA) purification strategies and yields.
| Defatting Agent | Purification Strategy | Number of Steps | Yield of WGA (mg Per 1 g Raw Wheat Germ) | Reference |
|---|---|---|---|---|
| Light petroleum | Extraction in water Protein precipitation 55% (NH4)2SO4 Dialysis in water Ion exchange chromatography using a DEAE 1-cellulose column, two SE 2-Sephadex columns, and QAE 3-Sephadex (×4) | 7 | 0.61 | [22] |
| Light petroleum | Extraction in water Protein precipitation 55% (NH4)2SO4 Dialysis in water Affinity chromatography CNAG 4-Sepharose (×1) | 4 | 0.3 | [25] |
| Acetone | Extraction in 0.05 N HC1 buffer Protein precipitation 35% (NH4)2SO4 n-Butanol treatment Dialysis in 0.05 N HC1 and second protein precipitation 35% (NH4)2SO4 Ion exchange chromatography on DEAE-cellulose column (×1) | 6 | 0.48 | [17] |
| n-Hexane | Extraction in 0.05 N HC1 buffer Tangential flow filtration MWCO 5 10 kDa Batch affinity purification on chitosan matrices (×1) | 3 | [26] |
1–DEAE–diethylaminoethyl; 2–sulphoethyl; 3–Diethyl-(2-hydroxypropyl)aminoethyl; 4–CNAG-2-acetamido-N-(ϵ-aminocaproyl)-2-deoxy-β-D-glucopyranosylamine; 5–MWCO–Molecular weight cut-off.
Table 2.
Summarized studies on WGA grafted drug delivery systems.
| Carrier Particle | Drug | Excipients | Animal/Cell Line | Outcome | Reference |
|---|---|---|---|---|---|
| Polymeric nanoparticles | Paclitaxel | Poly (lactic-co-glycolic acid) Isopropyl myristate | A549 H1299 CCL-186 cell lines | Demonstrated higher cytotoxicity | [54] |
| Polymeric nanoparticles | Paclitaxel | Poly (lactic-co-glycolic acid) | Caco-2 HT-29 cell lines | Increased intracellular retention in the Caco-2 and HT-29 cells Endocytosed nanoparticles could successfully escape from the endo-lysosome compartment and release into the cytosol with increasing incubation time | [55] |
| Polymeric nanoparticles | Docetaxel | Thiolated sodium alginate | HT-29 L929 cell lines | IC50 values of 52.9 µg/mL for HT-29 cells and 201.6 µg/mL for L929 cells Selectivity towards HT-29 cells over L929 cells | [56] |
| Polymeric nanoparticles | Etoposide Carmustine Doxorubicin | Methoxy poly(ethylene glycol)-poly(ε-caprolactone) Folic acid | U87MG cell line | WGA-modified surface promoted BBB permeation, and folic acid facilitated target site on U87MG cells Anti-proliferation against U87MG cells | [57] |
| Liposomes | Calcitonin | Carbopol | A549 cell line | Carbopol-WGA modification enhanced interaction with A549 lung epithelial cells compared with unmodified or CP-modified liposomes Enhanced and prolonged efficacy of calcitonin. | [59] |
| Microparticles | Insulin | Carboxymethylated kappa–carrageenan microparticles | Caco-2 cell line Sprague–Dawley rats | WGA-functionalized microparticles at 20 mg/mL showed a reduction in cell viability upon exposure The oral administration of insulin entrapped in the microparticles led to a prolonged duration of the hypoglycemic effect, up to 12–24 h, in diabetic rats. | [60] |
| Metal-oxide based nanoparticles | - | Iron oxide Fe3O4 | PZ-HPV-7 DU-145 PC-3 LNCaP cell lines DU-145 e BALB/c-nu/nu mice | 2.46 nM nanomagnet lectin exposure for 15 min induce apoptosis of cancer cells; Xenografted (DU-145) e BALB/c-nu/nu mice, where the tumor was not only completely arrested but also reduced. | [61] |
| Liposomes | Temoporfin | 1,2-dipalmitoyl-sn-glycero-3-phosphocholin N-[1-(2,3-dioleoyloxy)propyl]-N,N,N-trimethylammonium methyl sulfate 1,2- distearoyl-sn-glycero-3-phosphoethanolamine-N-[3-(N-succinimidyloxyglutaryl)aminopropyl(polyethyleneglycol)-2000-carbamyl] | Methicillin-resistant Staphylococcus aureus (MRSA) and Pseudomonas aeruginosa cultures | The WGA-modified liposomes eradicated all MRSA and significantly enhanced the photodynamic inactivation of P. aeruginosa. | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Balčiūnaitė-Murzienė, G.; Dzikaras, M. Wheat Germ Agglutinin—From Toxicity to Biomedical Applications. Appl. Sci. 2021, 11, 884. https://0-doi-org.brum.beds.ac.uk/10.3390/app11020884
AMA Style
Balčiūnaitė-Murzienė G, Dzikaras M. Wheat Germ Agglutinin—From Toxicity to Biomedical Applications. Applied Sciences. 2021; 11(2):884. https://0-doi-org.brum.beds.ac.uk/10.3390/app11020884
Chicago/Turabian StyleBalčiūnaitė-Murzienė, Gabrielė, and Mindaugas Dzikaras. 2021. "Wheat Germ Agglutinin—From Toxicity to Biomedical Applications" Applied Sciences 11, no. 2: 884. https://0-doi-org.brum.beds.ac.uk/10.3390/app11020884
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.