
The Potential Role of Myokines/Hepatokines in the Progression of Neuronal Damage in Streptozotocin and High-Fat Diet-Induced Type 2 Diabetes Mellitus Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
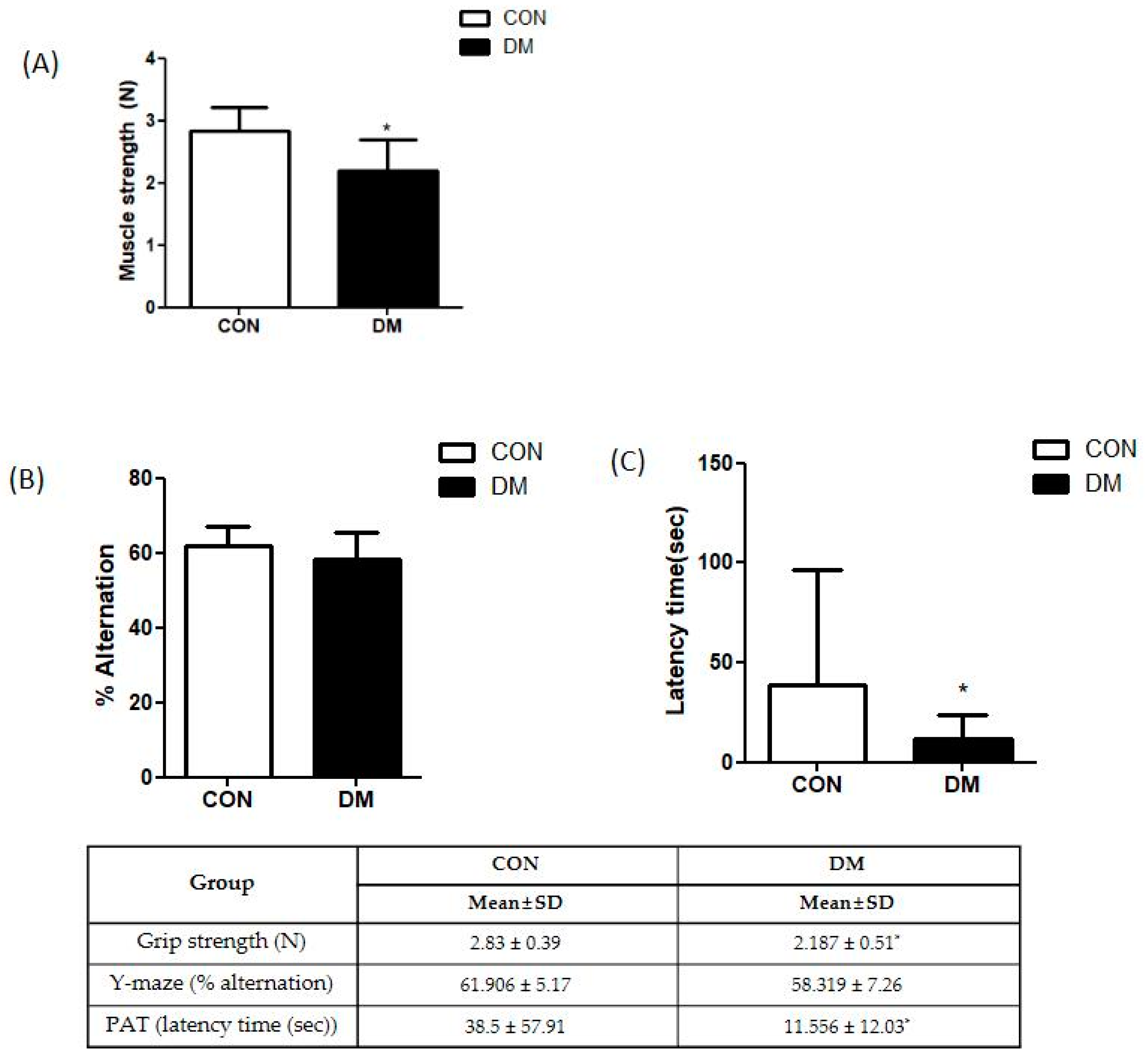
2.2. Muscle Function Test
2.3. Cognitive Function Test
2.4. Measurement of Body Composition
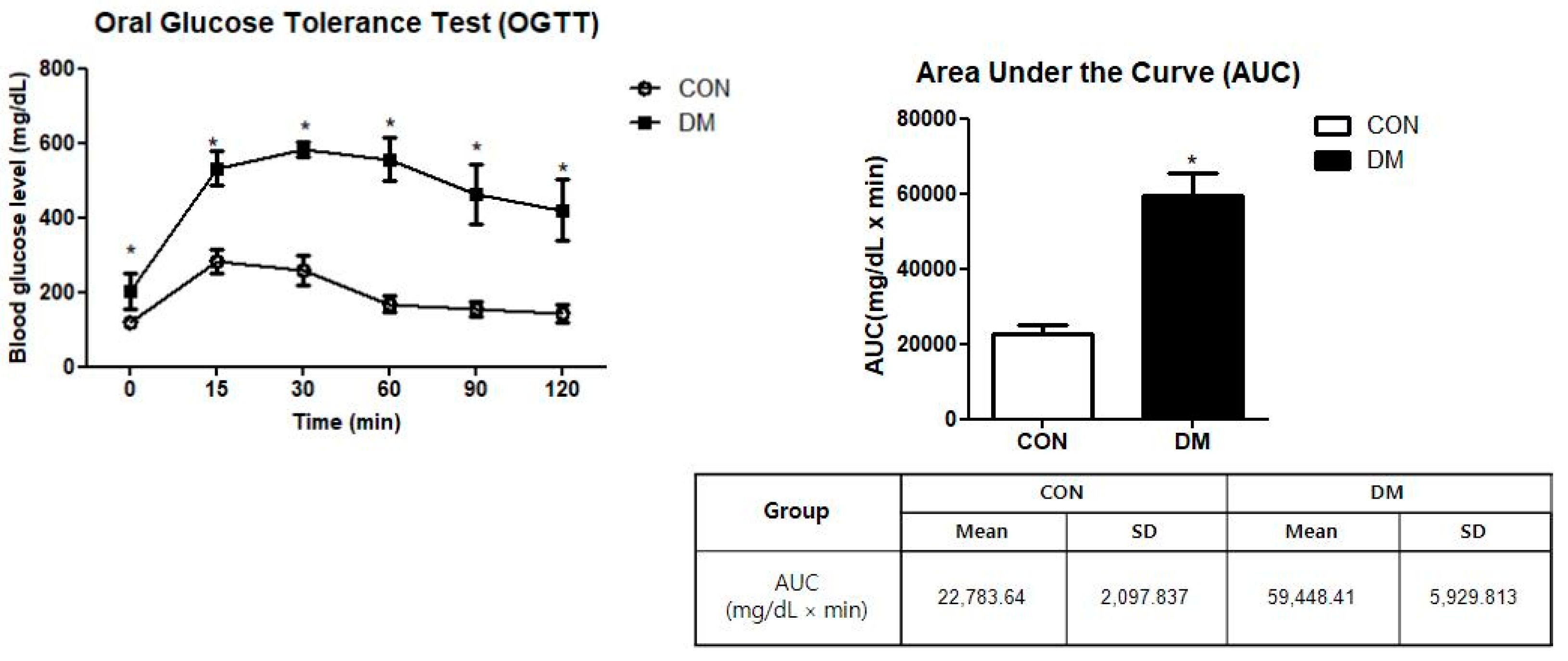
2.5. Oral Glucose Tolerance Test (OGTT)
= {15 min × (BG0 min + BG15 min) × ½} + {15 min × (BG15 min + BG30 min)
× ½} + {30 min × (BG30 min + BG60 min) × ½} + {30 min × (BG60 min + BG90
min) × ½} + {30 min × (BG90 min + BG120 min) × ½}
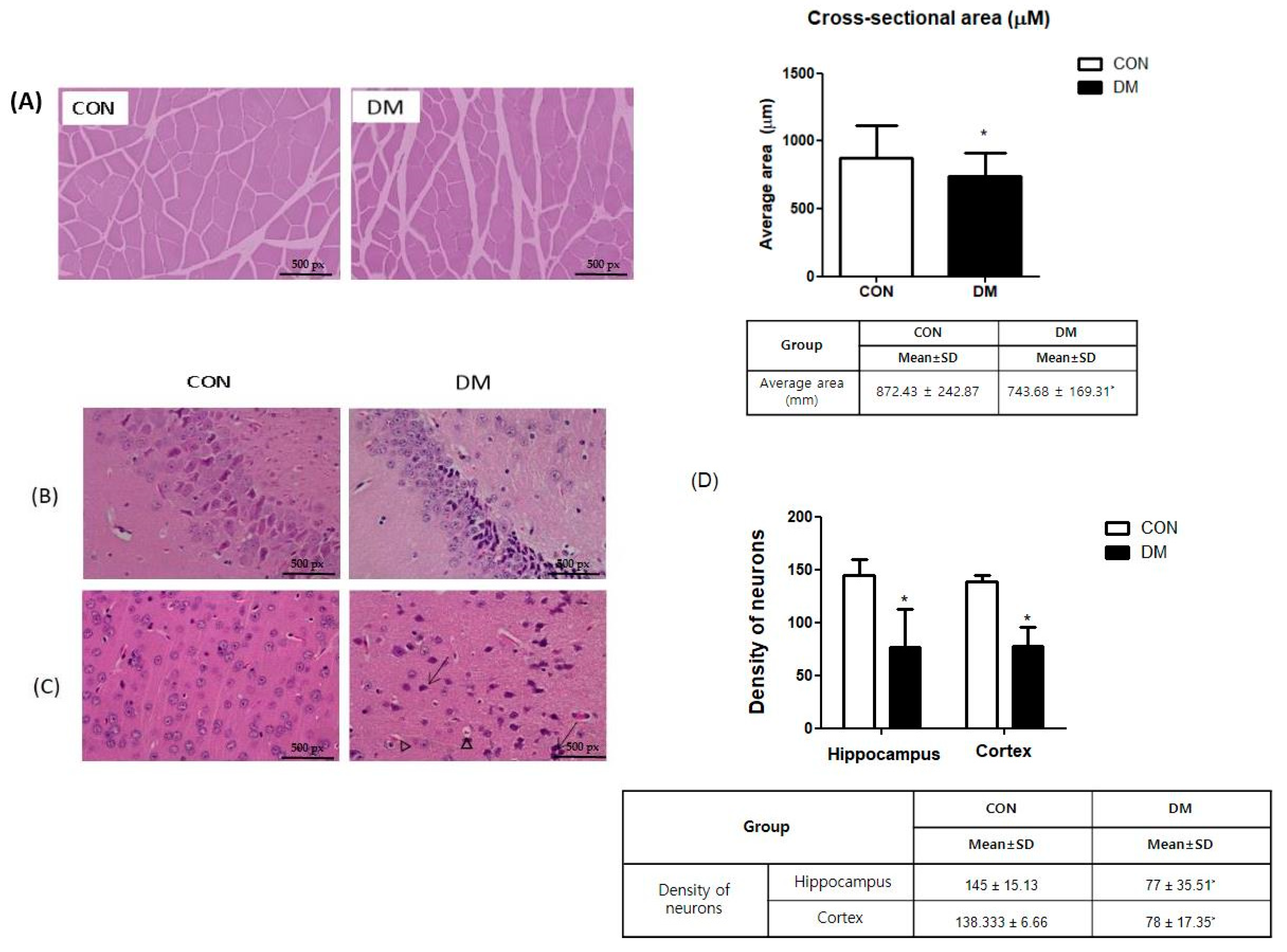
2.6. Histological Analysis
2.7. Hemoglobin A1c (HbA1c) % Level
2.8. Plasma Insulin Level
2.9. Homeostasis Model Assessment of IR (HOMA-IR) Level
2.10. Protein Extraction and Western Blot Analysis
2.11. Statistical Analysis
3. Results
3.1. Body Weight and Composition, Food Intake, and Glycemic Regulation in T2DM Mice
3.2. Glucose Tolerance in T2DM Mice
3.3. Skeletal Muscle Function in T2DM Mice
3.4. Cognitive Function in T2DM Mice
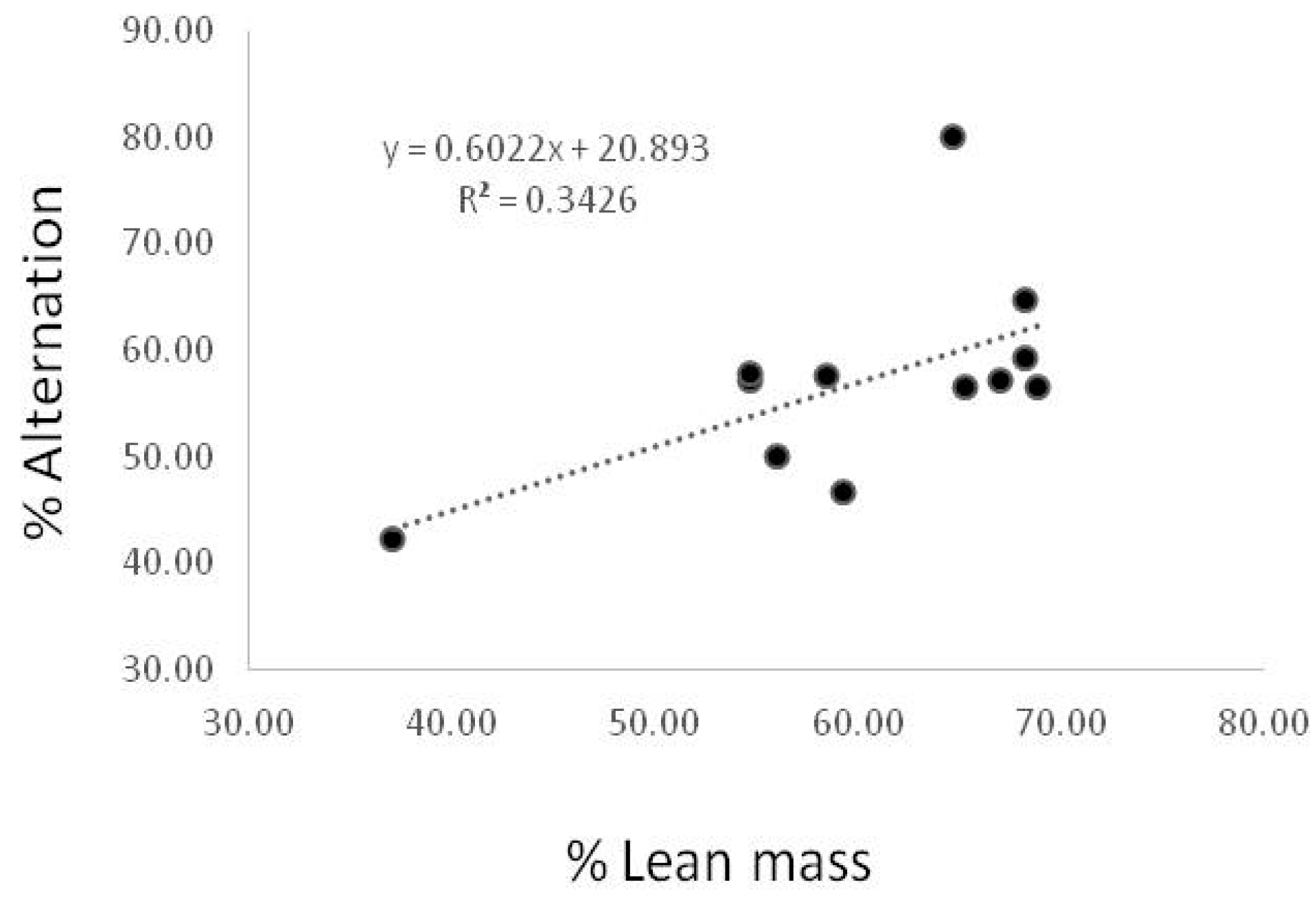
3.5. Correlation between % Lean Mass and Spontaneous Alternation Behavior
3.6. Morphological Changes of Skeletal Muscle in T2DM Mice
3.7. Morphological Changes of Brain in T2DM Mice
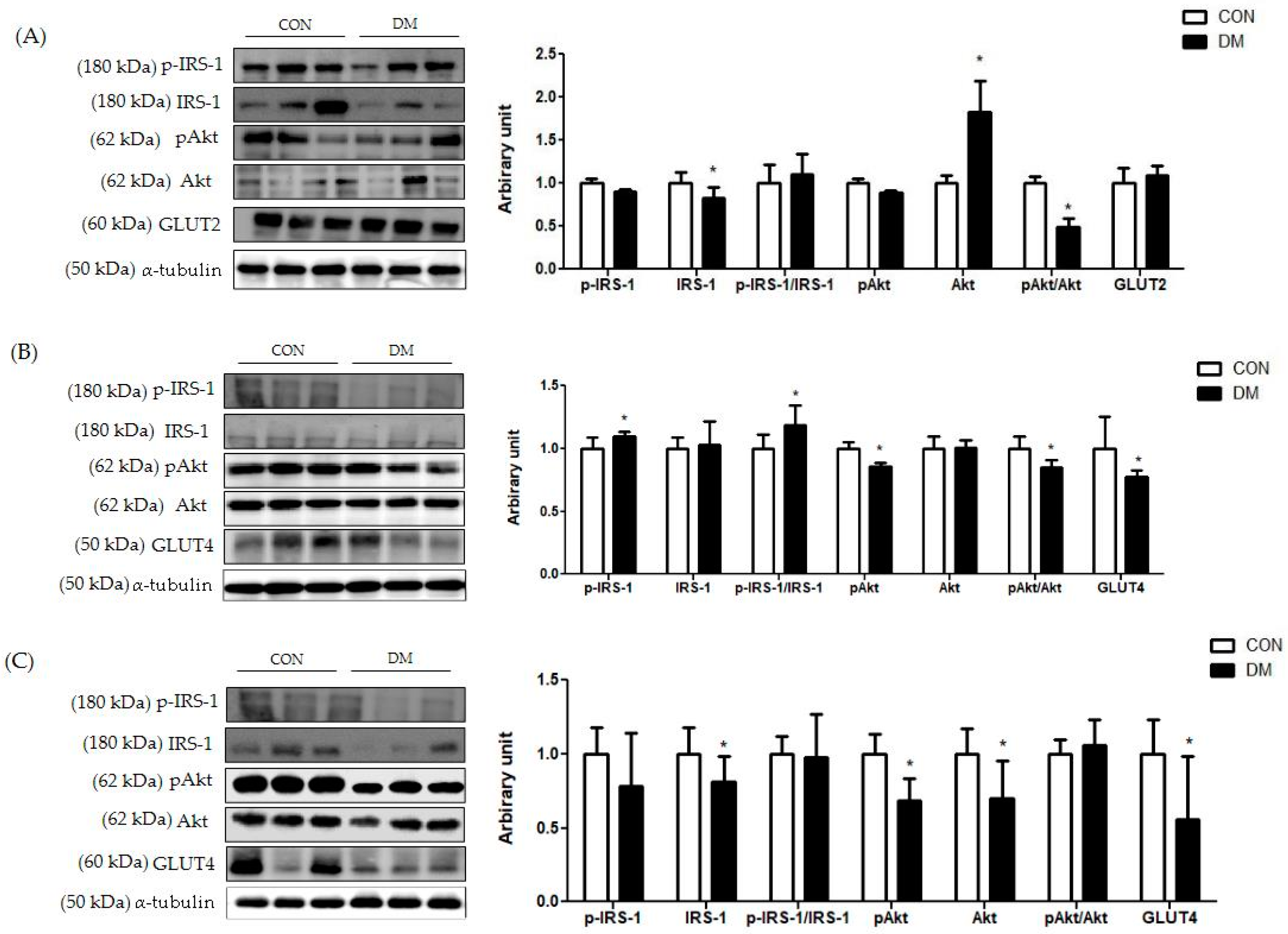
3.8. Insulin Signaling Related Markers in T2DM Mice
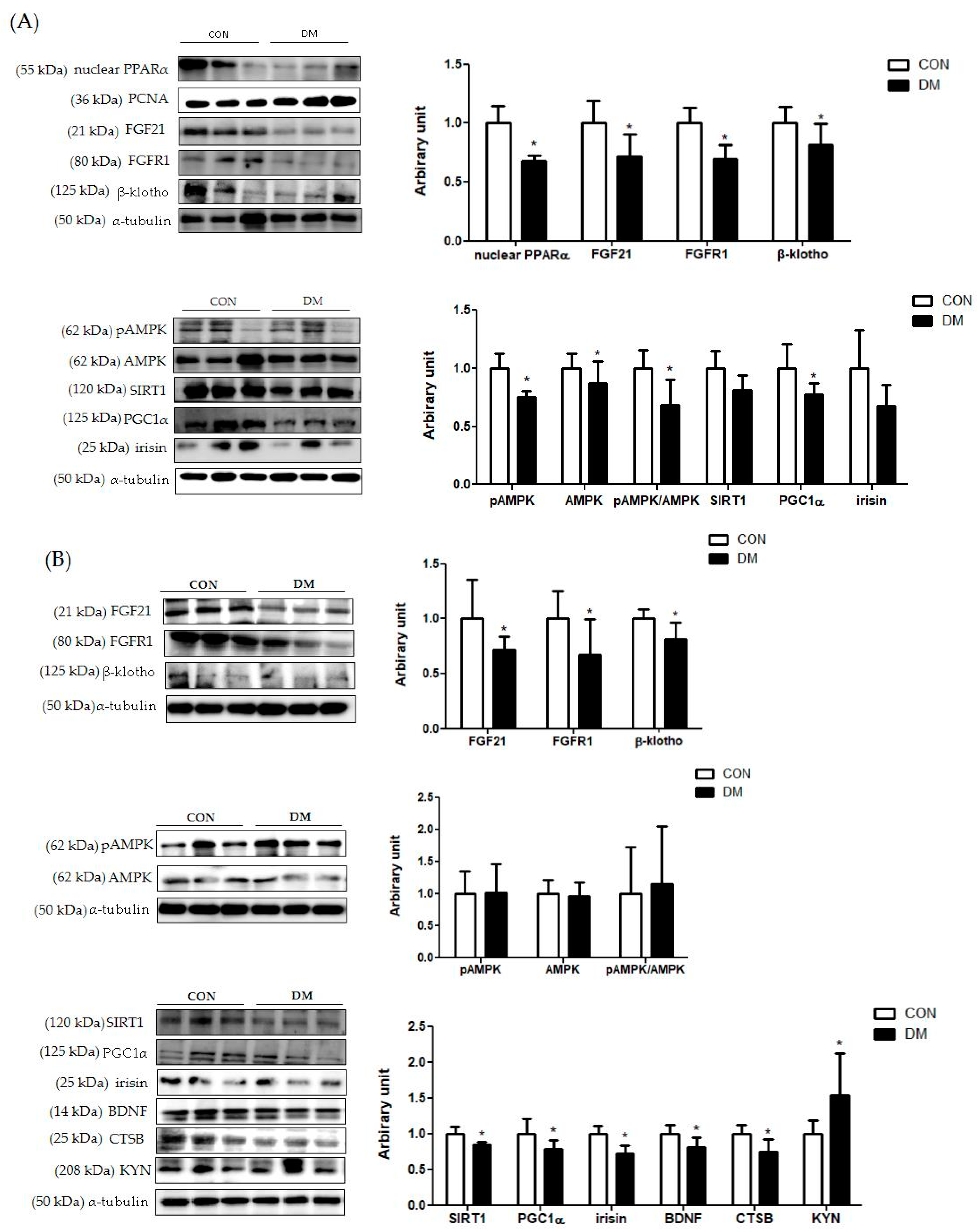
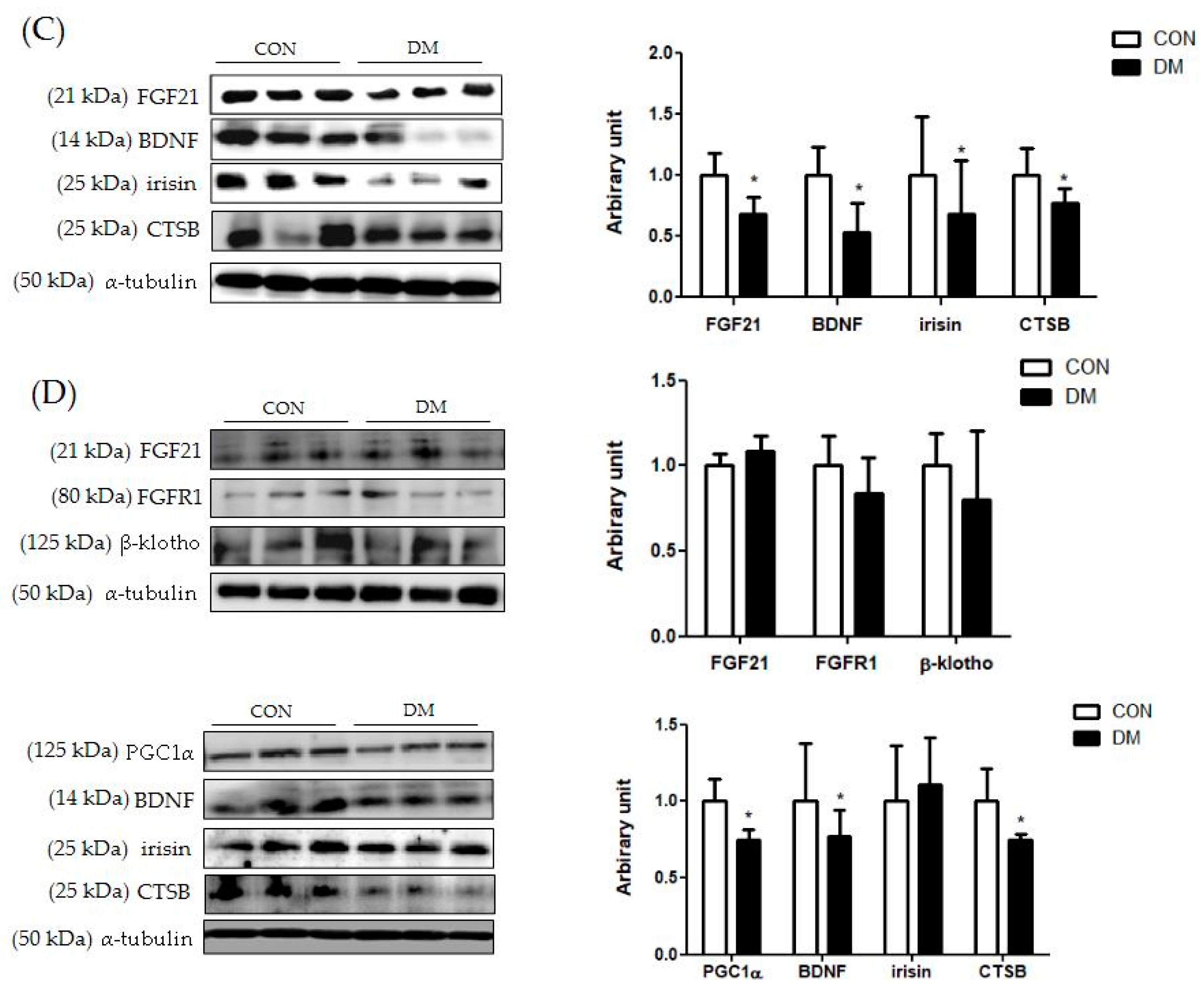
3.9. Energy Metabolism Related Markers with Myokines/Hepatokines and Neuronal Markers in T2DM Mice
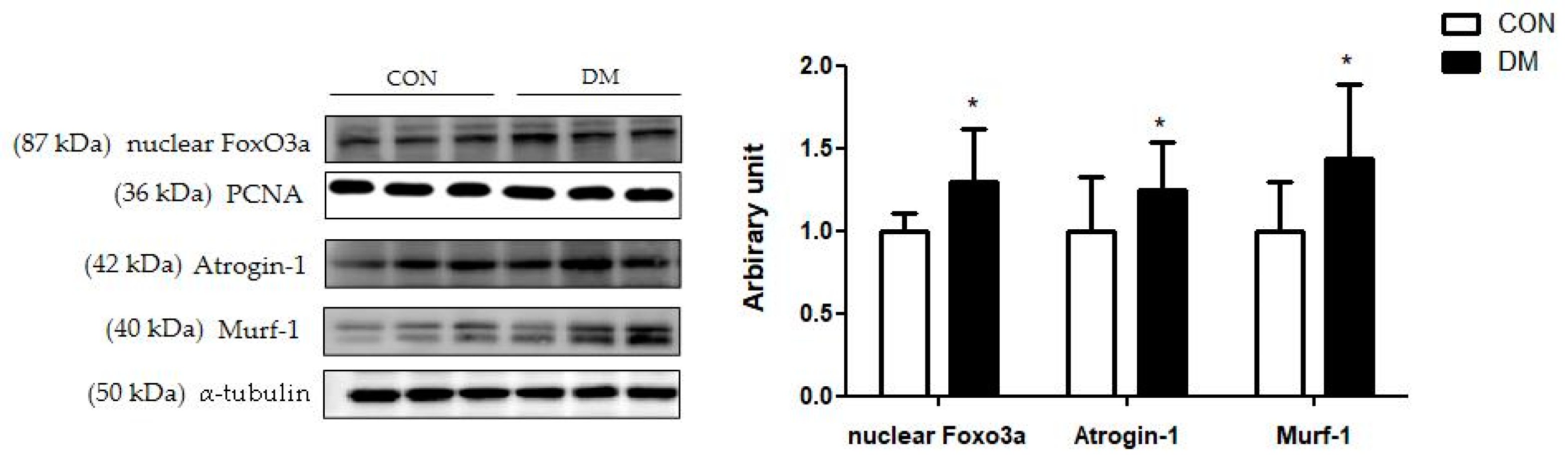
3.10. Skeletal Muscle Degradation Related Markers in T2DM Mice
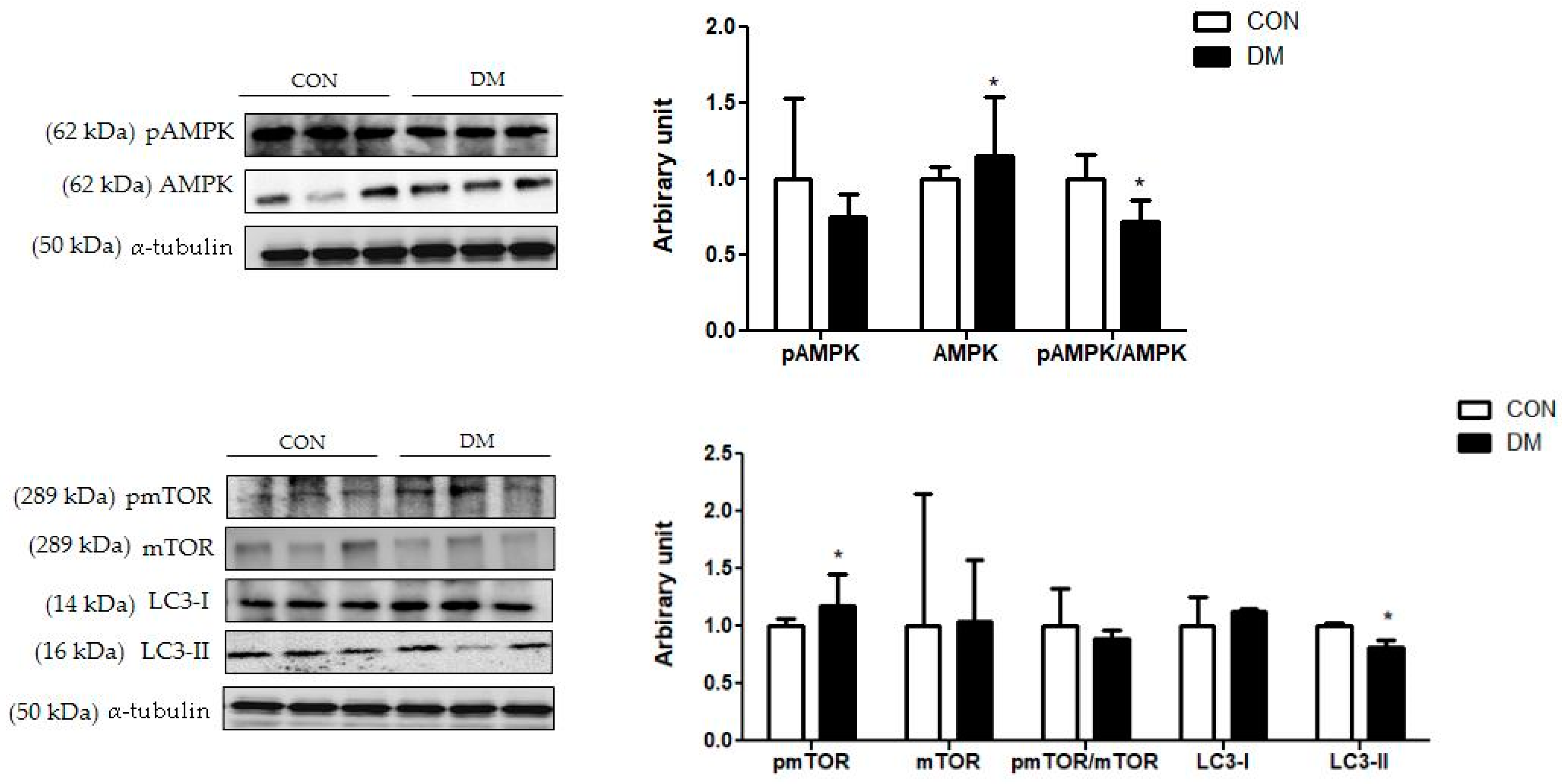
3.11. Hippocampal Autophagy Related Markers in T2DM Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maggio, C.A.; Pi-Sunyer, F.X. Obesity and type 2 diabetes. Endocrinol. Metab. Clin. N. Am. 2003, 32, 805. [Google Scholar] [CrossRef]
- Peng, T.-C.; Chen, W.-L.; Wu, L.-W.; Chang, Y.-W.; Kao, T.-W. Sarcopenia and cognitive impairment: A systematic review and meta-analysis. Clin. Nutr. 2019, 39, 2695–2701. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-V.; Hsu, T.-H.; Wu, W.-T.; Huang, K.-C.; Han, D.-S. Association Between Sarcopenia and Cognitive Impairment: A Systematic Review and Meta-Analysis. J. Am. Med Dir. Assoc. 2016, 17, 1164.e7–1164.e15. [Google Scholar] [CrossRef] [PubMed]
- Biessels, G.J.; Nobili, F.; Teunissen, E.C.; Simó, R.; Scheltens, P. Understanding multifactorial brain changes in type 2 diabetes: A biomarker perspective. Lancet Neurol. 2020, 19, 699–710. [Google Scholar] [CrossRef]
- Izzo, A.; Massimino, E.; Riccardi, G.; Della Pepa, G. A Narrative Review on Sarcopenia in Type 2 Diabetes Mellitus: Prevalence and Associated Factors. Nutrients 2021, 13, 183. [Google Scholar] [CrossRef]
- Chung, H.S.; Choi, K.M. Organokines in disease. Adv. Clin. Chem. 2020, 94, 261–321. [Google Scholar] [CrossRef]
- Della Guardia, L.; Codella, R. Exercise tolls the bell for key mediators of low-grade inflammation in dysmetabolic conditions. Cytokine Growth Factor Rev. 2021, 62, 83–93. [Google Scholar] [CrossRef]
- Itoh, N. FGF21 as a Hepatokine, Adipokine, and Myokine in Metabolism and Diseases. Front. Endocrinol. 2014, 5, 107. [Google Scholar] [CrossRef] [Green Version]
- Yen, H.-H.; Hsieh, S.-T.; Chen, C.-L.; Yang, W.-S.; Lee, P.-C.; Lin, M.-T.; Chen, C.-N.; Yang, P.-J. Circulating Diabetic Candidate Neurotrophic Factors, Brain-Derived Neurotrophic Factor and Fibroblast Growth Factor 21, in Sleeve Gastrectomy. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, B.; Wen, M.; Hu, M.; Peng, J.-G.; Wang, Y.; Fan, L.-L.; Tang, L. ZG02 Improved Hepatic Glucose Metabolism and Insulin Sensitivity via Activation of AMPK/Sirt1 Signaling Pathways in a High-fat Diet/Streptozotocin-induced Type 2 Diabetes Model. Diabetes Metab. Syndr. Obesity Targets Ther. 2020, 13, 4333–4339. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipido. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.K. Physical activity and muscle–brain crosstalk. Nat. Rev. Endocrinol. 2019, 15, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Isaac, A.R.; Lima-Filho, R.A.S.; Lourenco, M.V. How does the skeletal muscle communicate with the brain in health and disease? Neuropharmacology 2021, 1, 108744. [Google Scholar] [CrossRef]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle–Organ Crosstalk: The Emerging Roles of Myokines. Endocr. Rev. 2020, 41, 594–609. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Valaris, S.; Young, M.F.; Haley, E.B.; Luo, R.; Bond, S.F.; Mazuera, S.; Kitchen, R.R.; Caldarone, B.J.; Bettio, L.E.B.; et al. Exercise hormone irisin is a critical regulator of cognitive function. Nat. Metab. 2021, 3, 1058–1070. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and Synaptic Plasticity, Cognitive Function, and Dysfunction. Neurotrophic Factors 2014, 220, 223–250. [Google Scholar] [CrossRef]
- Jin, W. Regulation of BDNF-TrkB Signaling and Potential Therapeutic Strategies for Parkinson’s Disease. J. Clin. Med. 2020, 17, 257. [Google Scholar] [CrossRef] [Green Version]
- Mueller-Steiner, S.; Zhou, Y.; Arai, H.; Roberson, E.D.; Sun, B.; Chen, J.; Wang, X.; Yu, G.; Esposito, L.; Mucke, L.; et al. Antiamyloidogenic and neuroprotective functions of cathepsin B: Implications for Alzheimer’s disease. Neuron 2006, 21, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Embury, C.; Dyavarshetty, B.; Lu, Y.; Wiederin, J.L.; Ciborowski, P.; Gendelman, E.H.; Kiyota, T. Cathepsin B Improves ß-Amyloidosis and Learning and Memory in Models of Alzheimer’s Disease. J. Neuroimmune Pharmacol. 2016, 12, 340–352. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Lv, X.-Y.; Li, J.; Xu, Z.-G.; Chen, L. The Characterization of High-Fat Diet and Multiple Low-Dose Streptozotocin Induced Type 2 Diabetes Rat Model. Exp. Diabetes Res. 2008, 2008, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cole, C.L.; Beck, C.A.; Robinson, D.; Ye, J.; Mills, B.; Gerber, S.A.; Schwarz, E.M.; Linehan, D. Dual Energy X-ray Absorptiometry (DEXA) as a longitudinal outcome measure of cancer-related muscle wasting in mice. PLoS ONE 2020, 15, e0230695. [Google Scholar] [CrossRef]
- Winzell, M.S.; Ahrén, B. The high-fat diet-fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53, S215–S219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Lim, Y. Tocotrienol-rich fraction supplementation reduces hyperglycemia-induced skeletal muscle damage through regulation of insulin signaling and oxidative stress in type 2 diabetic mice. J. Nutr. Biochem. 2018, 57, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Bano, G. Glucose homeostasis, obesity and diabetes. Best Pract. Res. Clin. Obstet. Gynaecol. 2013, 27, 715–726. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, H.; Kim, S.Y.; Lim, Y. Effects of Lespedeza Bicolor Extract on Regulation of AMPK Associated Hepatic Lipid Metabolism in Type 2 Diabetic Mice. Antioxidants 2019, 8, 599. [Google Scholar] [CrossRef] [Green Version]
- Freeman, O.J.; Unwin, R.D.; Dowsey, A.W.; Begley, P.; Ali, S.; Hollywood, K.A.; Rustogi, N.; Petersen, R.S.; Dunn, W.B.; Cooper, G.J.; et al. Metabolic Dysfunction Is Restricted to the Sciatic Nerve in Experimental Diabetic Neuropathy. Diabetes 2015, 65, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Heurling, K.; Johansson, E.; Leuzy, A. Disturbances in brain energy metabolism in insulin resistance and diabetes and Alzheimer’s disease—Learnings from brain imaging biomarkers. Int. Rev. Neurobiol. 2020, 154, 111–130. [Google Scholar]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Regulation of longevity by FGF21: Interaction between energy metabolism and stress responses. Ageing Res. Rev. 2017, 37, 79–93. [Google Scholar] [CrossRef]
- Adams, V.; Wunderlich, S.; Mangner, N.; Hommel, J.; Esefeld, K.; Gielen, S.; Halle, M.; Ellingsen, Ø.; Van Craenenbroeck, E.M.; Wisløff, U.; et al. Ubiquitin-proteasome-system and enzymes of energy metabolism in skeletal muscle of patients with HFpEF and HFrEF. ESC Heart Fail. 2021, 8, 2556–2568. [Google Scholar] [CrossRef]
- Milan, G.; Romanello, V.; Pescatore, F.; Armani, A.; Paik, J.-H.; Frasson, L.; Seydel, A.; Zhao, J.; Abraham, R.; Goldberg, A.L.; et al. Regulation of autophagy and the ubiquitin–proteasome system by the FoxO transcriptional network during muscle atrophy. Nat. Commun. 2015, 6, 6670. [Google Scholar] [CrossRef] [Green Version]
- Laurens, C.; Bergouignan, A.; Moro, C. Exercise-Released Myokines in the Control of Energy Metabolism. Front. Physiol. 2020, 11, 91. [Google Scholar] [CrossRef]
- Mengeste, A.M.; Rustan, A.C.; Lund, J. Skeletal muscle energy metabolism in obesity. Obesity 2021, 29, 1582–1595. [Google Scholar] [CrossRef]
- Tingstad, R.H.; Norheim, F.; Haugen, F.; Feng, Y.Z.; Tunsjø, H.S.; Thoresen, G.H.; Rustan, A.C.; Charnock, C.; Aas, V. The effect of toll-like receptor ligands on energy metabolism and myokine expression and secretion in cultured human skeletal muscle cells. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Giudice, J.; Taylor, J.M. Muscle as a paracrine and endocrine organ. Curr. Opin. Pharmacol. 2017, 34, 49–55. [Google Scholar] [CrossRef]
- Kim, S.; Choi, J.-Y.; Moon, S.; Park, D.-H.; Kwak, H.-B.; Kang, J.-H. Roles of myokines in exercise-induced improvement of neuropsychiatric function. Pflug. Arch. 2019, 471, 491–505. [Google Scholar] [CrossRef]
- Delezie, J.; Handschin, C. Endocrine Crosstalk Between Skeletal Muscle and the Brain. Front. Neurol. 2018, 9, 698–712. [Google Scholar] [CrossRef]
- Sakuma, K.; Yamaguchi, A. The recent understanding of the neurotrophin’s role in skeletal muscle adaptation. J. Biomed. Biotechnol. 2011, 2011, 201696. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-Y.; Wang, T.-Y.; Chen, S.-L.; Chang, Y.-H.; Chen, P.-S.; Huang, S.-Y.; Tzeng, N.-S.; Wang, L.-J.; Lee, I.H.; Chen, K.C.; et al. The correlation between plasma brain-derived neurotrophic factor and cognitive function in bipolar disorder is modulated by the BDNF Val66Met polymorphism. Sci. Rep. 2016, 6, 37950. [Google Scholar] [CrossRef]
- Yin, Q.; Zhang, L.; Han, X.; Zhang, H.; Wang, F.; Qin, X.; Zhuang, P.; Zhang, Y. Zi Shen Wan Fang regulates kynurenine metabolism to alleviate diabetes-associated cognitive impairment via activating the skeletal muscle PGC1α-PPARα signaling. Phytomedicine 2022, 99, 154000. [Google Scholar] [CrossRef]
- Rao, A.A. Views and opinion on BDNF as a target for diabetic cognitive dysfunction. Bioinformation 2013, 9, 551–554. [Google Scholar] [CrossRef]
- Wang, C.; Bomberg, E.; Billington, C.J.; Levine, A.S.; Kotz, C.M. Brain-derived neurotrophic factor (BDNF) in the hypothalamic ventromedial nucleus increases energy expenditure. Brain Res. 2010, 1336, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Milstein, J.L.; Ferris, H.A. The brain as an insulin-sensitive metabolic organ. Mol. Metab. 2021, 52, 101234. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, R.; Ma, Z. Autophagy and Energy Metabolism. Biol. Dis. 2019, 1206, 329–357. [Google Scholar] [CrossRef]
- Madhavi, Y.; Gaikwad, N.; Yerra, V.G.; Kalvala, A.K.; Nanduri, S.; Kumar, A. Targeting AMPK in Diabetes and Diabetic Complications: Energy Homeostasis, Autophagy and Mitochondrial Health. Curr. Med. Chem. 2019, 26, 5207–5229. [Google Scholar] [CrossRef]
- Seldin, M.M.; Koplev, S.; Rajbhandari, P.; Vergnes, L.; Rosenberg, G.M.; Meng, Y.; Pan, C.; Phuong, T.M.; Gharakhanian, R.; Che, N.; et al. A Strategy for Discovery of Endocrine Interactions with Application to Whole-Body Metabolism. Cell Metab. 2018, 27, 1138–1155.e6. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GROUP | CON | DM | ||
|---|---|---|---|---|
| Median (Min, Max) | Mean (SD) | Median (Min, Max) | Mean (SD) | |
| Body weight (g) | ||||
| Initial body weight | 26.25 (24.12, 30.09) | 26.77 (1.67) | 30.02 (27.25, 30.05) | 30.4 (2.24) * |
| Final body weight | 34.84 (31.7, 39.38) | 35.23 (2.52) | 41 (33.97, 45.414) | 40.55 (3.46) * |
| Gain | 3.26 (2.48, 3.06) | 3.63 (0.96) | 5.69 (1.94, 7.67) | 5.32 (1.92) |
| % Fat | 29.40 (28.05, 41.39) | 31.61 (5.0) | 40.34 (32.55, 59.3) | 42.40 (9.07) * |
| % Lean | 67.65 (54.67, 68.9) | 65.31 (5.42) | 57.24 (37.13, 65.35) | 55.16 (9.56) * |
| Gastrocnemius weight (g/kg BW) | 0.54 (0.48, 0.59) | 0.54 (0.03) | 0.43 (0.35, 0.48) | 0.42 (0.039) * |
| Quadriceps weight (g/kg BW) | 0.72 (0.59, 0.75) | 0.71 (0.05) | 0.55 (0.43, 0.61) | 0.53 (0.05) * |
| Food Intake (g/day) | 2.73 (2.32, 3.23) | 2.72 (0.26) | 2.53 (2.03, 3.29) | 2.53 (0.34) |
| Fasting blood glucose level (mg/dL) | 143 (119, 173) | 146.36 (15.09) | 263 (177,439) | 291.63 (89.45) * |
| Plasma insulin (μU/mL) | 8.66 (5.59, 18.88) | 10.0 (4.07) | 13.88 (7.3, 49.93) | 22.49 (16.96) * |
| HbA1c % | 4.95 (4.7, 5.4) | 5.0 (0.24) | 5.7 (3.2, 6.95) | 5.55 (1.15) * |
| HOMA-IR (mmol/L × μU/mL) | 2.91 (2.14, 7.5) | 3.65 (1.7) | 12.96 (5.06, 39.54) | 16.01 (11.39) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Lim, Y. The Potential Role of Myokines/Hepatokines in the Progression of Neuronal Damage in Streptozotocin and High-Fat Diet-Induced Type 2 Diabetes Mellitus Mice. Biomedicines 2022, 10, 1521. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10071521
Lee H, Lim Y. The Potential Role of Myokines/Hepatokines in the Progression of Neuronal Damage in Streptozotocin and High-Fat Diet-Induced Type 2 Diabetes Mellitus Mice. Biomedicines. 2022; 10(7):1521. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10071521
Chicago/Turabian StyleLee, Heaji, and Yunsook Lim. 2022. "The Potential Role of Myokines/Hepatokines in the Progression of Neuronal Damage in Streptozotocin and High-Fat Diet-Induced Type 2 Diabetes Mellitus Mice" Biomedicines 10, no. 7: 1521. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10071521