
Formation of Giant Lipid Vesicles in the Presence of Nonelectrolytes—Glucose, Sucrose, Sorbitol and Ethanol

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
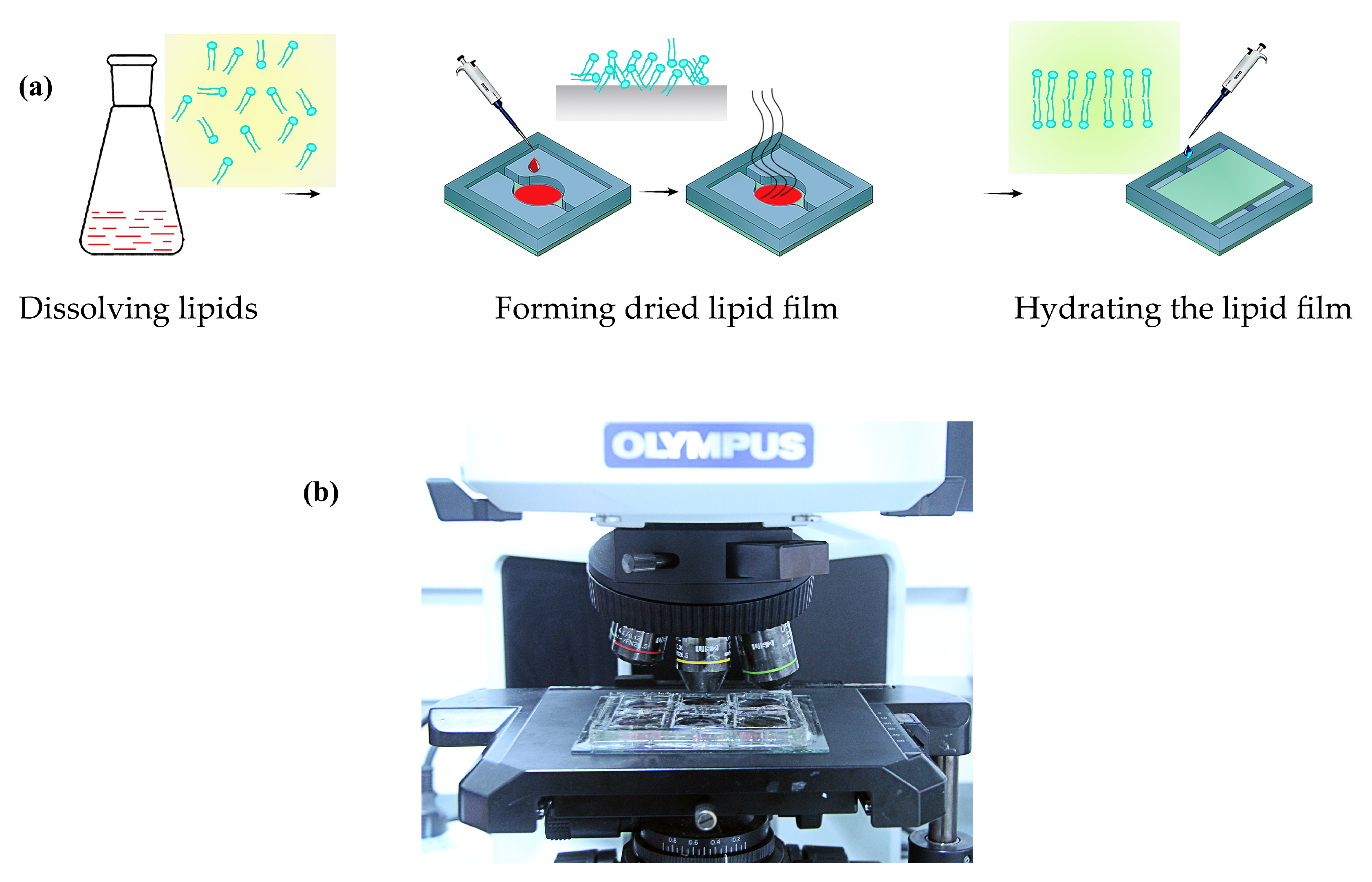
2.2. The Experimental Method
2.3. Calculation of the Relative Fluorescence Intensity and the Relative Membrane Continuity
3. Results
3.1. The Effects of Sucrose, Glucose and Sorbitol
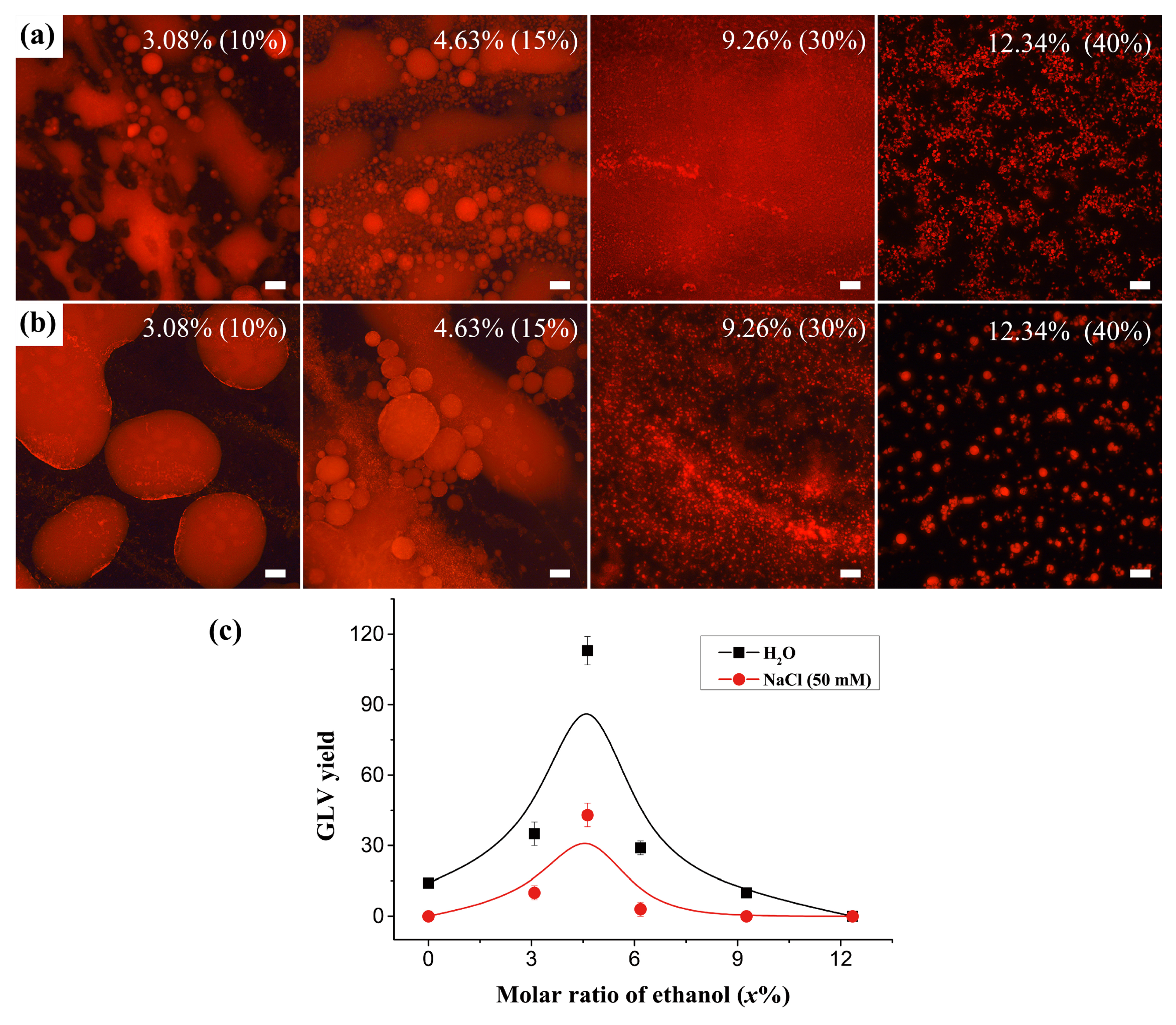
3.2. The Effect of Ethanol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stein, H.; Spindler, S.; Bonakdar, N.; Wang, C.; Sandoghdar, V. Production of Isolated Giant Unilamellar Vesicles under High Salt Concentrations. Front. Physiol. 2017, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Dickey, A.N.; Faller, R. How Alcohol Chain-Length and Concentration Modulate Hydrogen Bond Formation in a Lipid Bilayer. Biophys. J. 2007, 92, 2366–2376. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, I.L.; Kemmer, G.C.; Pomorski, T.G. Membrane protein reconstitution into giant unilamellar vesicles: A review on current techniques. Eur. Biophys. J. 2017, 46, 103–119. [Google Scholar] [CrossRef]
- Fenz, S.F.; Sengupta, K. Giant vesicles as cell models. Integr. Biol. 2012, 4, 982–995. [Google Scholar] [CrossRef]
- Saha, A.; Akhtar, N.; Kumar, V.; Kumar, S.; Srivastava, H.K.; Kumar, S.; Manna, D. pH-Regulated anion transport activities of bis(iminourea) derivatives across the cell and vesicle membrane. Org. Biomol. Chem. 2019, 17, 5779–5788. [Google Scholar] [CrossRef]
- Jenkins, E.; Santos, A.M.; O’Brien-Ball, C.; Felce, J.H.; Wilcock, M.J.; Hatherley, D.; Dustin, M.L.; Davis, S.J.; Eggeling, C.; Sezgin, E. Reconstitution of immune cell interactions in free-standing membranes. J. Cell Sci. 2019, 132, jcs219709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pott, T.; Bouvrais, H.; Méléard, P. Giant unilamellar vesicle formation under physiologically relevant conditions. Chem. Phys. Lipids 2008, 154, 115–119. [Google Scholar] [CrossRef]
- Montes, L.-R.; Alonso, A.; Goñi, F.M.; Bagatolli, L.A. Giant Unilamellar Vesicles Electroformed from Native Membranes and Organic Lipid Mixtures under Physiological Conditions. Biophys. J. 2007, 93, 3548–3554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, H.; Yang, B.; Wang, L.; Cao, W.; Han, X. Electroformation of giant unilamellar vesicles using interdigitated ITO electrodes. J. Mater. Chem. A 2013, 1, 7125–7130. [Google Scholar] [CrossRef]
- Tsuji, G.; Sunami, T.; Ichihashi, N. Production of giant unilamellar vesicles by the water-in-oil emulsion-transfer method without high internal concentrations of sugars. J. Biosci. Bioeng. 2018, 126, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Oka, M.; Tanaka, T.; Yamazaki, M. A new method for the preparation of giant liposomes in high salt concentrations and growth of protein microcrystals in them. Biochim. Biophys. Acta (BBA) Biomembr. 2002, 1561, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Angelova, M.I. Liposome Electroformation. Perspect. Supramol. Chem. 2007, 81, 26–36. [Google Scholar] [CrossRef]
- Angelova, M.; Dimitrov, D.S. A mechanism of liposome electroformation. Trends Colloid Interface Sci. II 2007, 59–67. [Google Scholar] [CrossRef]
- Wang, Q.; Li, W.; Hu, N.; Chen, X.; Fan, T.; Wang, Z.; Yang, Z.; Cheney, M.A.; Yang, J. Ion concentration effect (Na+ and Cl−) on lipid vesicle formation. Colloids Surf. B: Biointerfaces 2017, 155, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Wang, Q.; Hu, N.; Liao, Y.; Chen, X.; Wang, Z.; Yang, Z.; Yang, J.; Qian, S. Preparation of giant lipid vesicles with controllable sizes by a modified hydrophilic polydimethylsiloxane microarray chip. J. Colloid Interface Sci. 2019, 536, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, A.S.; Crowe, R.J.H. Membrane stabilization during freezing: The role of two natural cryoprotectants, trehalose and proline. Cryobiology 1985, 22, 367–377. [Google Scholar] [CrossRef]
- Konov, K.B.; Leonov, D.V.; Isaev, N.P.; Fedotov, K.Y.; Voronkova, V.K.; Dzuba, S.A. Membrane–Sugar Interactions Probed by Pulsed Electron Paramagnetic Resonance of Spin Labels. J. Phys. Chem. B 2015, 119, 10261–10266. [Google Scholar] [CrossRef]
- Koynova, R.; Brankov, J.; Tenchov, B. Modulation of lipid phase behavior by kosmotropic and chaotropic solutes. Eur. Biophys. J. 1997, 25, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.D.; Wang, C.; Arleth, L.; Peters, G.H.; Westh, P. Reconciliation of opposing views on membrane–sugar interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 1874–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Ohmae, H.; Hatta, I. Trehalose-induced destabilization of interdigitated gel phase in dihexadecylphosphatidylcholine. Biophys. J. 1997, 73, 3030–3038. [Google Scholar] [CrossRef] [Green Version]
- Caffrey, M.; Fonseca, V.; Leopold, A.C. Lipid-Sugar Interactions: Relevance to Anhydrous Biology. Plant Physiol. Mar. 1988, 86, 754–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikelj, M.; Praper, T.; Demič, R.; Hodnik, V.; Turk, T.; Anderluh, G. Electroformation of giant unilamellar vesicles from erythrocyte membranes under low-salt conditions. Anal. Biochem. 2013, 435, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Nagle, J.F.; Jablin, M.S.; Tristram-Nagle, S. Sugar does not affect the bending and tilt moduli of simple lipid bilayers. Chem. Phys. Lipids 2016, 196, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Marcus, Y. Effect of Ions on the Structure of Water: Structure Making and Breaking. Chem. Rev. 2009, 109, 1346–1370. [Google Scholar] [CrossRef]
- Mancinelli, R.; Botti, A.; Bruni, A.F.; Ricci, M.A.; Soper, A.K. Hydration of Sodium, Potassium, and Chloride Ions in Solution and the Concept of Structure Maker/Breaker. J. Phys. Chem. B 2007, 111, 13570–13577. [Google Scholar] [CrossRef] [PubMed]
- Imberti, S.; Botti, A.; Bruni, F.; Cappa, G.; Ricci, M.A.; Soper, A.K. Ions in water: The microscopic structure of concentrated hydroxide solutions. J. Chem. Phys. 2005, 122, 194509. [Google Scholar] [CrossRef]
- Botti, A.; Bruni, F.; Imberti, S.; Ricci, M.A.; Soper, A.K. Ions in water: The microscopic structure of concentrated NaOH solutions. J. Chem. Phys. 2004, 120, 10154. [Google Scholar] [CrossRef]
- Vácha, R.; Jurkiewicz, P.; Petrov, M.; Berkowitz, M.L.; Böckmann, R.A.; Barucha-Kraszewska, J.; Hof, M.; Jungwirth, P. Mechanism of Interaction of Monovalent Ions with Phosphatidylcholine Lipid Membranes. J. Phys. Chem. B 2010, 114, 9504–9509. [Google Scholar] [CrossRef] [PubMed]
- Vácha, R.; Siu, S.W.I.; Petrov, M.; Böckmann, R.; Barucha-Kraszewska, J.; Jurkiewicz, P.; Hof, M.; Berkowitz, M.L.; Jungwirth, P. Effects of Alkali Cations and Halide Anions on the DOPC Lipid Membrane. J. Phys. Chem. A 2009, 113, 7235–7243. [Google Scholar] [CrossRef]
- Skibinsky, A.; Venable, R.M.; Pastor, R.W. A Molecular Dynamics Study of the Response of Lipid Bilayers and Monolayers to Trehalose. Biophys. J. 2005, 89, 4111–4121. [Google Scholar] [CrossRef] [Green Version]
- Lambruschini, C.; Relini, A.; Ridi, A.; Cordone, L.; Gliozzi, A. Trehalose Interacts with Phospholipid Polar Heads in Langmuir Monolayers. Langmuir 2000, 16, 5467–5470. [Google Scholar] [CrossRef]
- Leekumjorn, S.; Sum, A.K. Molecular investigation of the interactions of trehalose with lipid bilayers of DPPC, DPPE and their mixture. Mol. Simul. 2006, 32, 219–230. [Google Scholar] [CrossRef]
- Villarreal, M.A.; Díaz, S.B.; Disalvo, E.A.; Montich, G.G. Molecular Dynamics Simulation Study of the Interaction of Trehalose with Lipid Membranes. Langmuir 2004, 20, 7844–7851. [Google Scholar] [CrossRef]
- Ma, C.D.; Wang, C.; Acevedo-Vélez, C.; Gellman, S.H.; Abbott, N.L. Modulation of hydrophobic interactions by proximally immobilized ions. Nat. Cell Biol. 2015, 517, 347–350. [Google Scholar] [CrossRef]
- Ricker, J.V.; Tsvetkova, N.M.; Wolkers, W.F.; Leidy, C.; Tablin, F.; Longo, M.; Crowe, J.H. Trehalose Maintains Phase Separation in an Air-Dried Binary Lipid Mixture. Biophys. J. 2003, 84, 3045–3051. [Google Scholar] [CrossRef] [Green Version]
- Fabrie, C.H.; De Kruijff, B.; De Gier, J. Protection by sugars against phase transition-induced leak in hydrated dimyristoylphosphatidylcholine liposomes. Biochim. Biophys. Acta (BBA) Biomembr. 1990, 1024, 380–384. [Google Scholar] [CrossRef]
- Zavaglia, A.G.; Tymczyszyn, E.; De Antoni, G.; Disalvo, E.A. Action of trehalose on the preservation of Lactobacillus delbrueckii ssp. bulgaricus by heat and osmotic dehydration. J. Appl. Microbiol. 2003, 95, 1315–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Leopold, A.; Crowe, L.; Crowe, J. Stability of dry liposomes in sugar glasses. Biophys. J. 1996, 70, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Belton, P.S.; Gil, A.M. IR and Raman spectroscopic studies of the interaction of trehalose with hen egg white lysozyme. Biopolymers 1994, 34, 957–961. [Google Scholar] [CrossRef]
- Bryant, G.; Koster, K. Dehydration of solute–lipid systems: Hydration forces analysis. Colloids Surf. B Biointerfaces 2004, 35, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Lenné, T.; Garvey, C.J.; Koster, K.; Bryant, G. Effects of Sugars on Lipid Bilayers during Dehydration—SAXS/WAXS Measurements and Quantitative Model. J. Phys. Chem. B 2009, 113, 2486–2491. [Google Scholar] [CrossRef]
- Kapla, J.; Wohlert, J.; Stevensson, B.; Engström, O.; Widmalm, G.; Maliniak, A. Molecular Dynamics Simulations of Membrane–Sugar Interactions. J. Phys. Chem. B 2013, 117, 6667–6673. [Google Scholar] [CrossRef] [PubMed]
- Bogaart, G.V.D.; Hermans, N.; Krasnikov, V.; de Vries, A.H.; Poolman, B. On the Decrease in Lateral Mobility of Phospholipids by Sugars. Biophys. J. 2007, 92, 1598–1605. [Google Scholar] [CrossRef] [Green Version]
- Manca, M.L.; Castangia, I.; Matricardi, P.; Lampis, S.; Fernàndez-Busquets, X.; Fadda, A.M.; Manconi, M. Molecular arrangements and interconnected bilayer formation induced by alcohol or polyalcohol in phospholipid vesicles. Colloids Surf. B Biointerfaces 2014, 117, 360–367. [Google Scholar] [CrossRef]
- Feller, S.E.; Brown, C.A.; Nizza, D.T.; Gawrisch, K. Nuclear Overhauser Enhancement Spectroscopy Cross-Relaxation Rates and Ethanol Distribution across Membranes. Biophys. J. 2002, 82, 1396–1404. [Google Scholar] [CrossRef] [Green Version]
- Cevc, G.; Löbbecke, L.; Nagel, N.; Vierl, U. Phospholipid-Alcohol Interactions: Effects of Chain-Length and Headgroup Variations. Phosphorus Sulfur Silicon Relat. Elem. 1996, 109, 285–288. [Google Scholar] [CrossRef]
- Wanderlingh, U.; D’Angelo, G.; Nibali, V.C.; Crupi, C.; Rifici, S.; Corsaro, C.; Sabatino, G. Interaction of alcohol with phospholipid membrane: NMR and XRD investigations on DPPC–hexanol system. Spectroscopy 2010, 24, 375–380. [Google Scholar] [CrossRef]
- Spector, M.S.; Selinger, J.V.; Schnur, J.M. Thermodynamics of Phospholipid Tubules in Alcohol/Water Solutions. J. Am. Chem. Soc. 1997, 119, 8533–8539. [Google Scholar] [CrossRef]
- Cacela, C.; Hincha, D.K. Low Amounts of Sucrose Are Sufficient to Depress the Phase Transition Temperature of Dry Phosphatidylcholine, but Not for Lyoprotection of Liposomes. Biophys. J. 2006, 90, 2831–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziebert, F.; Lacoste, D. A Poisson–Boltzmann approach for a lipid membrane in an electric field. New J. Phys. 2010, 12, 095002. [Google Scholar] [CrossRef]
- Böckmann, R.; Hac, A.; Heimburg, T.; Grubmüller, H. Effect of Sodium Chloride on a Lipid Bilayer. Biophys. J. 2003, 85, 1647–1655. [Google Scholar] [CrossRef] [Green Version]
- Chong, Y.; Kleinhammes, A.; Tang, P.; Xu, Y.; Wu, Y. Dominant Alcohol–Protein Interaction via Hydration-Enabled Enthalpy-Driven Binding Mechanism. J. Phys. Chem. B 2015, 119, 5367–5375. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Hu, N.; Lei, J.; Qing, Q.; Huang, J.; Tao, K.; Zhao, S.; Sun, K.; Yang, J. Formation of Giant Lipid Vesicles in the Presence of Nonelectrolytes—Glucose, Sucrose, Sorbitol and Ethanol. Processes 2021, 9, 945. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9060945
Wang Q, Hu N, Lei J, Qing Q, Huang J, Tao K, Zhao S, Sun K, Yang J. Formation of Giant Lipid Vesicles in the Presence of Nonelectrolytes—Glucose, Sucrose, Sorbitol and Ethanol. Processes. 2021; 9(6):945. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9060945
Chicago/Turabian StyleWang, Qiong, Ning Hu, Jincan Lei, Qiurong Qing, Jing Huang, Ke Tao, Shixian Zhao, Ke Sun, and Jun Yang. 2021. "Formation of Giant Lipid Vesicles in the Presence of Nonelectrolytes—Glucose, Sucrose, Sorbitol and Ethanol" Processes 9, no. 6: 945. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9060945