
β-Glucan Production by Levilactobacillus brevis and Pediococcus claussenii for In Situ Enriched Rye and Wheat Sourdough Breads


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Materials
2.2. Sourdough Fermentation
2.3. Cell Count, pH, and Analysis of Sourdough Microbiota
2.4. Sourdough Bread Preparation and Baking
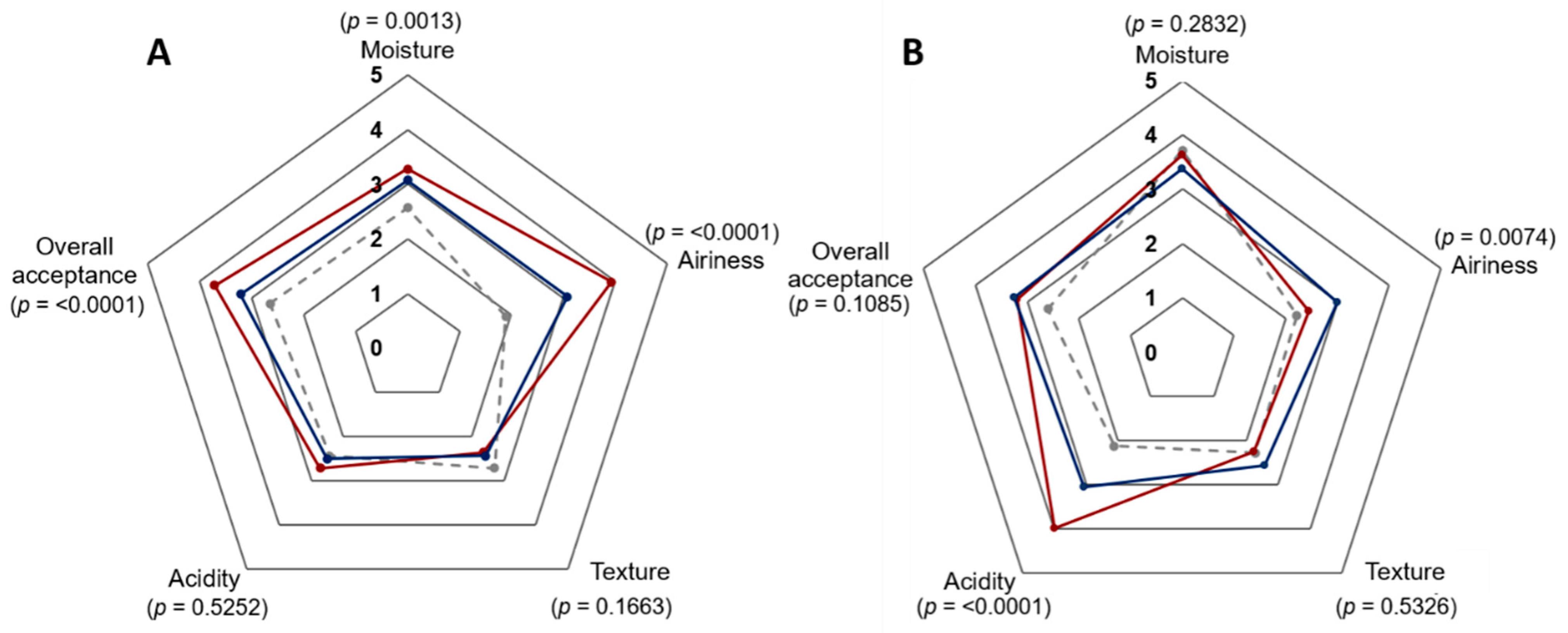
2.5. Sensory Analysis
2.6. Isolation and Purification of β-Glucan
2.7. Quantification of the Bacterial β-Glucan
2.7.1. Quantification by HPLC
2.7.2. Quantification by ELISA
2.8. Statistical Analysis
3. Results
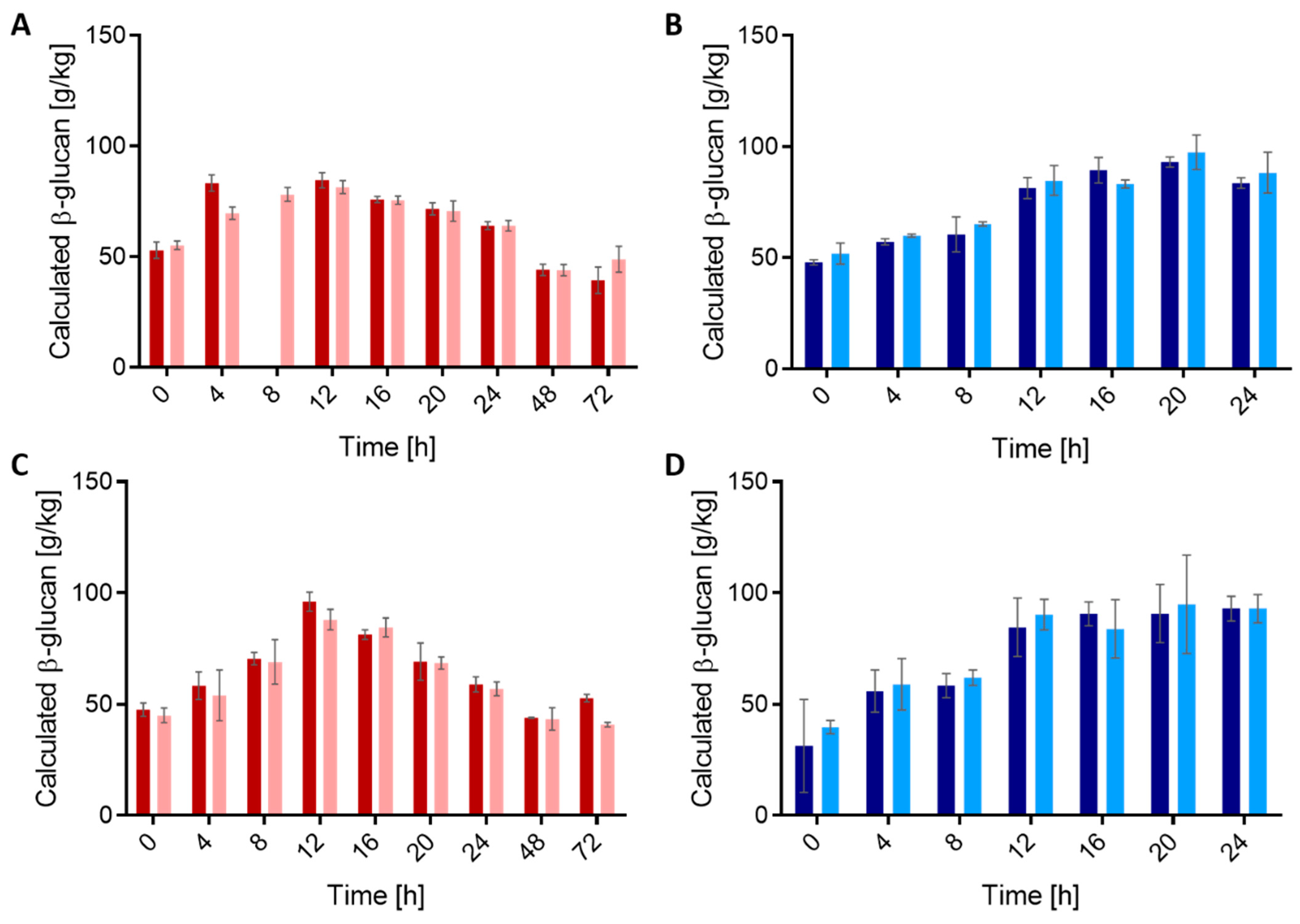
3.1. Growth Characteristics of Strains in Wheat and Rye Sourdoughs
3.1.1. Growth Characteristics of Strains Co-Cultivated with La. plantarum TMW 1.2330
3.1.2. Determining Plasmid Stability
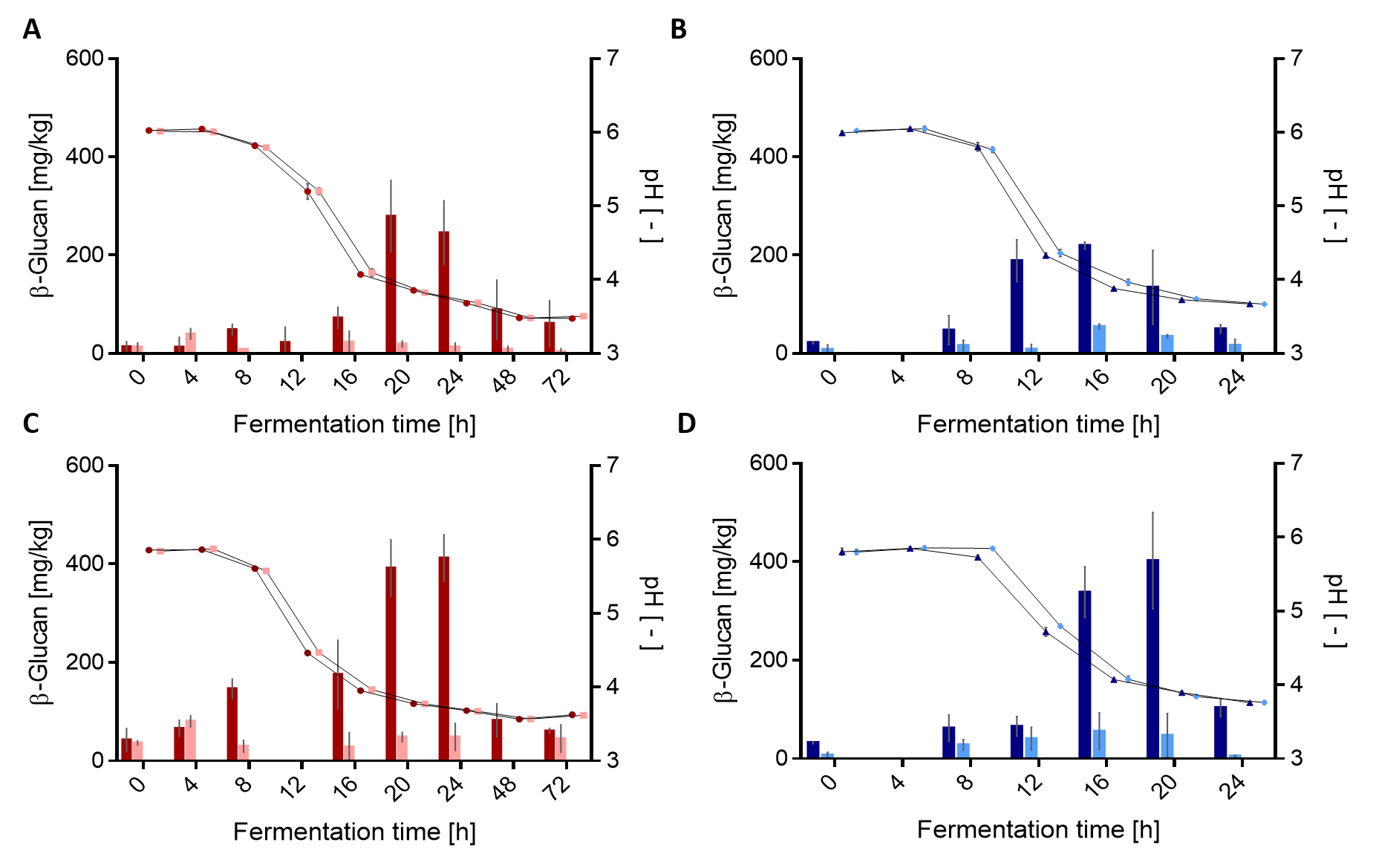
3.2. Quantification of β-Glucan in Fermented Sourdoughs
3.2.1. Quantification by HPLC
3.2.2. Quantification by ELISA
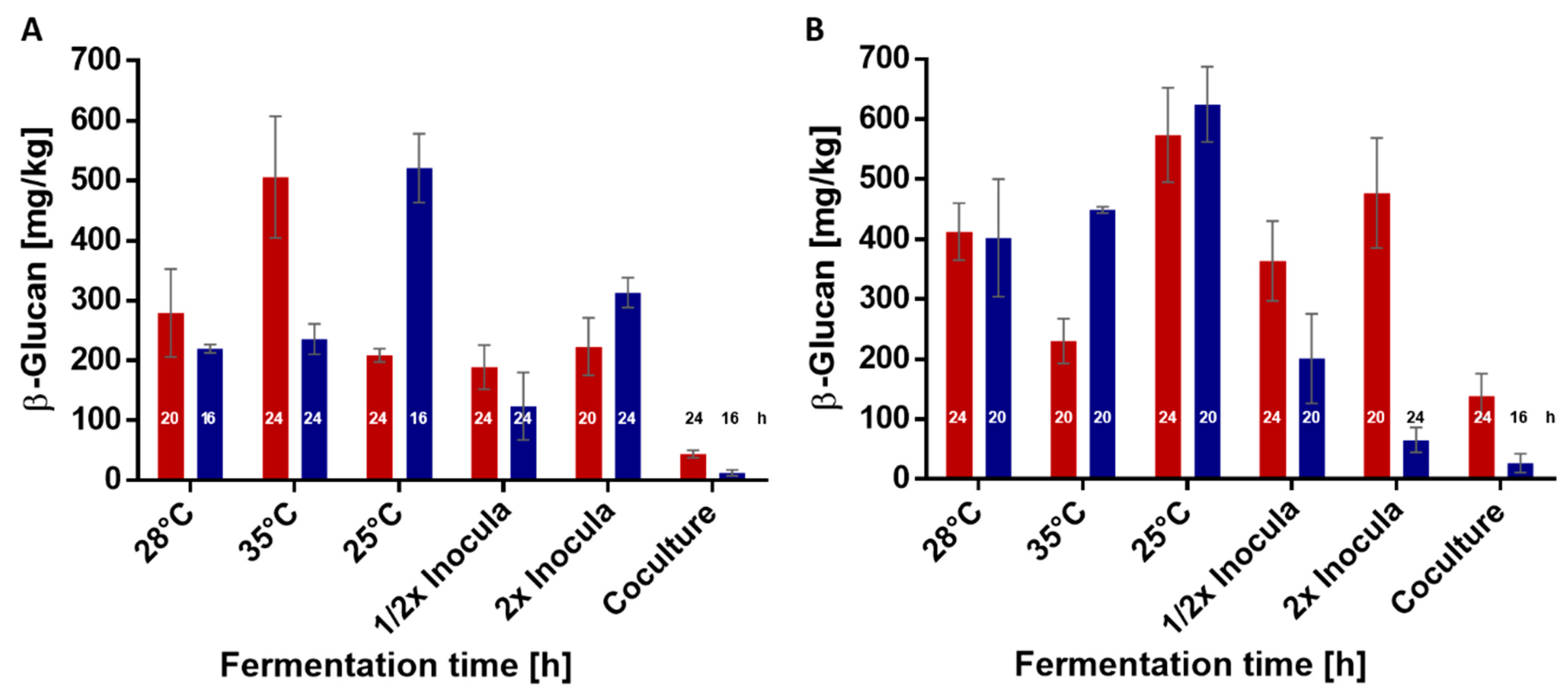
3.3. Parameters Influencing In Situ Formation of β-Glucan in Wheat and Rye Sourdoughs
3.4. Analysis of β-Glucan-Enriched Bread Characterisics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Tech. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Ray, B. Food Biopreservatives of Microbial Origin; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Katina, K.; Heiniö, R.-L.; Autio, K.; Poutanen, K. Optimization of sourdough process for improved sensory profile and texture of wheat bread. LWT Food Sci. Technol. 2006, 39, 1189–1202. [Google Scholar] [CrossRef]
- Arendt, E.K.; Ryan, L.A.; Dal Bello, F. Impact of sourdough on the texture of bread. Food Microbiol. 2007, 24, 165–174. [Google Scholar] [CrossRef]
- Gerez, C.L.; Torino, M.I.; Rollán, G.; de Valdez, G.F. Prevention of bread mould spoilage by using lactic acid bacteria with antifungal properties. Food Control. 2009, 20, 144–148. [Google Scholar] [CrossRef]
- Galle, S.; Arendt, E.K. Exopolysaccharides from sourdough lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2014, 54, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Rühmkorf, C.; Rübsam, H.; Becker, T.; Bork, C.; Voiges, K.; Mischnick, P.; Brandt, M.J.; Vogel, R.F. Effect of structurally different microbial homoexopolysaccharides on the quality of gluten-free bread. Eur. Food Res. Technol. 2012, 235, 139–146. [Google Scholar] [CrossRef]
- Chen, X.Y.; Levy, C.; Ganzle, M.G. Structure-function relationships of bacterial and enzymatically produced reuterans and dextran in sourdough bread baking application. Int. J. Food Microbiol. 2016, 239, 95–102. [Google Scholar] [CrossRef]
- Galle, S.; Schwab, C.; Arendt, E.K.; Ganzle, M.G. Structural and rheological characterisation of heteropolysaccharides produced by lactic acid bacteria in wheat and sorghum sourdough. Food Microbiol. 2011, 28, 547–553. [Google Scholar] [CrossRef]
- Poutanen, K.; Flander, L.; Katina, K. Sourdough and cereal fermentation in a nutritional perspective. Food Microbiol. 2009, 26, 693–699. [Google Scholar] [CrossRef]
- Russo, P.; Lopez, P.; Capozzi, V.; de Palencia, P.F.; Duenas, M.T.; Spano, G.; Fiocco, D. Beta-glucans improve growth, viability and colonization of probiotic microorganisms. Int. J. Mol. Sci. 2012, 13, 6026–6039. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramos, A.; Mohedano, M.L.; López, P.; Spano, G.; Fiocco, D.; Russo, P.; Capozzi, V. In situ β-glucan fortification of cereal-based matrices by Pediococcus parvulus 2.6: Technological aspects and prebiotic potential. Int. J. Mol. Sci. 2017, 18, 1588. [Google Scholar] [CrossRef] [Green Version]
- Salazar, N.; Gueimonde, M.; De Los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Exopolysaccharides produced by lactic acid bacteria and bifidobacteria as fermentable substrates by the intestinal microbiota. Crit. Rev. Food Sci. Nutr. 2016, 56, 1440–1453. [Google Scholar] [CrossRef] [PubMed]
- Welman, A.D.; Maddox, I.S. Exopolysaccharides from lactic acid bacteria: Perspectives and challenges. Trends Biotechnol. 2003, 21, 269–274. [Google Scholar] [CrossRef]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsu, W.H.; Pan, T.M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory effects triggered by lactic acid bacteria exopolysaccharides: New insights into molecular interactions with host cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- Notararigo, S.; Nacher-Vazquez, M.; Ibarburu, I.; Werning, M.L.; de Palencia, P.F.; Duenas, M.T.; Aznar, R.; Lopez, P.; Prieto, A. Comparative analysis of production and purification of homo- and hetero-polysaccharides produced by lactic acid bacteria. Carbohydr. Polym. 2013, 93, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Galle, S.; Schwab, C.; Arendt, E.; Ganzle, M. Exopolysaccharide-forming Weissella strains as starter cultures for sorghum and wheat sourdoughs. J. Agric. Food Chem. 2010, 58, 5834–5841. [Google Scholar] [CrossRef]
- van Hijum, S.A.; Kralj, S.; Ozimek, L.K.; Dijkhuizen, L.; van Geel-Schutten, I.G. Structure-function relationships of glucansucrase and fructansucrase enzymes from lactic acid bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 157–176. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; De Vin, F.; Vaningelgem, F.; Degeest, B. Recent developments in the biosynthesis and applications of heteropolysaccharides from lactic acid bacteria. Int. Dairy J. 2001, 11, 687–707. [Google Scholar] [CrossRef]
- Fraunhofer, M.E.; Jakob, F.; Vogel, R.F. Influence of Different Sugars and Initial pH on β-Glucan Formation by Lactobacillus brevis TMW 1.2112. Curr. Microbiol. 2018, 75, 794–802. [Google Scholar] [CrossRef]
- Tieking, M.; Korakli, M.; Ehrmann, M.A.; Gänzle, M.G.; Vogel, R.F. In situ production of exopolysaccharides during sourdough fermentation by cereal and intestinal isolates of lactic acid bacteria. Appl. Environ. Microbiol. 2003, 69, 945–952. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Degeest, B. Heteropolysaccharides from lactic acid bacteria. FEMS Microbiol. Rev. 1999, 23, 153–177. [Google Scholar] [CrossRef]
- Fraunhofer, M. Characterization of EPS-Producing Brewery-Associated Lactobacilli. Ph.D. Thesis, Technical University of Munich, Munich, Germany, 2018. [Google Scholar]
- Patel, S.; Majumder, A.; Goyal, A. Potentials of exopolysaccharides from lactic Acid bacteria. Indian J. Microbiol. 2012, 52, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M. The sourdough microflora: Interactions of lactic acid bacteria and yeasts. Trends Food Sci. Technol. 1998, 9, 267–274. [Google Scholar] [CrossRef]
- KučeroVá, J. Effects of location and year on technological quality and pentosan content in rye. Czech J. Food Sci. 2009, 27, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Anaya, M.A. Enzymes and bread flavor. J. Agric. Food Chem. 1996, 44, 2469–2480. [Google Scholar] [CrossRef]
- Bockwoldt, J.A.; Stahl, L.; Ehrmann, M.A.; Vogel, R.F.; Jakob, F. Persistence and β-glucan formation of beer-spoiling lactic acid bacteria in wheat and rye sourdoughs. Food Microbiol. 2020, 91, 103539. [Google Scholar] [CrossRef] [PubMed]
- Korakli, M.; Rossmann, A.; Ganzle, M.G.; Vogel, R.F. Sucrose metabolism and exopolysaccharide production in wheat and rye sourdoughs by Lactobacillus sanfranciscensis. J. Agric. Food Chem. 2001, 49, 5194–5200. [Google Scholar] [CrossRef] [PubMed]
- Korakli, M.; Vogel, R.F. Structure/function relationship of homopolysaccharide producing glycansucrases and therapeutic potential of their synthesised glycans. Appl. Microbiol. Biotechnol. 2006, 71, 790–803. [Google Scholar] [CrossRef]
- Bounaix, M.S.; Gabriel, V.; Morel, S.; Robert, H.; Rabier, P.; Remaud-Simeon, M.; Gabriel, B.; Fontagne-Faucher, C. Biodiversity of exopolysaccharides produced from sucrose by sourdough lactic acid bacteria. J. Agric. Food Chem. 2009, 57, 10889–10897. [Google Scholar] [CrossRef] [PubMed]
- Kaditzky, S.; Seitter, M.; Hertel, C.; Vogel, R.F. Performance of Lactobacillus sanfranciscensis TMW 1.392 and its levansucrase deletion mutant in wheat dough and comparison of their impact on bread quality. Eur. Food Res. Technol. 2007, 227, 433–442. [Google Scholar] [CrossRef]
- Mårtensson, O.; Chasco, M.-D.; Irastorza, A.; Holst, O.; Rudling, M.; Norin, E.; Midtvedt, T.; Öste, R. Effects of fermented, ropy, non-dairy, oat-based products on serum lipids and the faecal excretion of cholesterol and short chain fatty acids in germfree and conventional rats. Nutr. Res. 2002, 22, 1461–1473. [Google Scholar] [CrossRef]
- Notararigo, S.; de Las Casas-Engel, M.; de Palencia, P.F.; Corbi, A.L.; Lopez, P. Immunomodulation of human macrophages and myeloid cells by 2-substituted (1-3)-β-D-glucan from P. parvulus 2.6. Carbohydr. Polym. 2014, 112, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Fraunhofer, M.E.; Geissler, A.J.; Wefers, D.; Bunzel, M.; Jakob, F.; Vogel, R.F. Characterization of β-glucan formation by Lactobacillus brevis TMW 1.2112 isolated from slimy spoiled beer. Int. J. Biol. Macromol. 2018, 107, 874–881. [Google Scholar] [CrossRef]
- Fraunhofer, M.E.; Geissler, A.J.; Jakob, F.; Vogel, R.F. Multiple Genome Sequences of Exopolysaccharide-Producing, Brewery-Associated Lactobacillus brevis Strains. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K. 125th Anniversary Review: Microbiological Instability of Beer Caused by Spoilage Bacteria. J. Inst. Brew. 2011, 117, 131–155. [Google Scholar] [CrossRef]
- Schurr, B.C.; Behr, J.; Vogel, R.F. Role of the GAD system in hop tolerance of Lactobacillus brevis. Eur. Food Res. Technol. 2013, 237, 199–207. [Google Scholar] [CrossRef]
- Duenas-Chasco, M.T.; Rodriguez-Carvajal, M.A.; Mateo, P.T.; Franco-Rodriguez, G.; Espartero, J.L.; Irastorza-Iribas, A.; Gil-Serrano, A.M. Structural analysis of the exopolysaccharide produced by Pediococcus damnosus 2.6. Carbohydr. Res. 1997, 303, 453–458. [Google Scholar] [CrossRef]
- Kern, C.C.; Usbeck, J.C.; Vogel, R.F.; Behr, J. Optimization of Matrix-Assisted-Laser-Desorption-Ionization-Time-Of-Flight Mass Spectrometry for the identification of bacterial contaminants in beverages. J. Microbiol. Methods 2013, 93, 185–191. [Google Scholar] [CrossRef]
- Kupetz, M.; Geißinger, C.; Gastl, M.; Becker, T. Comparison of Dumas and Kjeldahl method for nitrogen determination in malt, wort and beer. Brew. Sci. 2018, 71, 18–23. [Google Scholar]
- Rühmkorf, C.; Jungkunz, S.; Wagner, M.; Vogel, R.F. Optimization of homoexopolysaccharide formation by lactobacilli in gluten-free sourdoughs. Food Microbiol. 2012, 32, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Influence of levan-producing acetic acid bacteria on buckwheat-sourdough breads. Food Microbiol. 2017, 65, 95–104. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; Codd, R. Measurement of (1→3),(1→4)-β-D-glucan in barley and oats: A streamlined enzymic procedure. J. Sci. Food Agric. 1991, 55, 303–312. [Google Scholar] [CrossRef]
- Werning, M.L.; Perez-Ramos, A.; de Palencia, P.F.; Mohedano, M.L.; Duenas, M.T.; Prieto, A.; Lopez, P. A specific immunological method to detect and quantify bacterial 2-substituted (1,3)-β-D-glucan. Carbohydr. Polym. 2014, 113, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Walling, E.; Gindreau, E.; Lonvaud-Funel, A. A putative glucan synthase gene dps detected in exopolysaccharide-producing Pediococcus damnosus and Oenococcus oeni strains isolated from wine and cider. Int. J. Food Microbiol. 2005, 98, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Comino, P.; Shelat, K.; Collins, H.; Lahnstein, J.; Gidley, M.J. Separation and purification of soluble polymers and cell wall fractions from wheat, rye and hull less barley endosperm flours for structure-nutrition studies. J. Agric. Food Chem. 2013, 61, 12111–12122. [Google Scholar] [CrossRef]
- Stolz, P.; Böcker, G.; Vogel, R.F.; Hammes, W.P. Utilisation of maltose and glucose by lactobacilli isolated from sourdough. FEMS Microbiol. Lett. 1993, 109, 237–242. [Google Scholar] [CrossRef]
- Pittet, V.; Abegunde, T.; Marfleet, T.; Haakensen, M.; Morrow, K.; Jayaprakash, T.; Schroeder, K.; Trost, B.; Byrns, S.; Bergsveinson, J.; et al. Complete genome sequence of the beer spoilage organism Pediococcus claussenii ATCC BAA-344T. J. Bacteriol. 2012, 194, 1271–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llull, D.; Munoz, R.; Lopez, R.; Garcia, E. A single gene (tts) located outside the cap locus directs the formation of Streptococcus pneumoniae type 37 capsular polysaccharide: Type 37 pneumococci are natural, genetically binary strains. J. Exp. Med. 1999, 190, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Stier, H.; Ebbeskotte, V.; Gruenwald, J. Immune-modulatory effects of dietary Yeast β-1,3/1,6-D-glucan. Nutr. J. 2014, 13, 38. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, T.; Carter, G. β-glucan biological activities: A review (condensed version). Glyco. Sci. Nutr. 2000, 1, 1–6. [Google Scholar]
- Boone, C.; Sommer, S.S.; Hensel, A.; Bussey, H. Yeast KRE genes provide evidence for a pathway of cell wall beta-glucan assembly. J. Cell Biol. 1990, 110, 1833–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, A.A.; Armö, E.; Grangeon, E.; Fredriksson, H.; Andersson, R.; Åman, P. Molecular weight and structure units of (1→3, 1→4)-β-glucans in dough and bread made from hull-less barley milling fractions. J. Cereal Sci. 2004, 40, 195–204. [Google Scholar] [CrossRef]
- Becker, S.; Tebben, J.; Coffinet, S.; Wiltshire, K.; Iversen, M.H.; Harder, T.; Hinrichs, K.U.; Hehemann, J.H. Laminarin is a major molecule in the marine carbon cycle. Proc. Natl. Acad. Sci. USA 2020, 117, 6599–6607. [Google Scholar] [CrossRef] [Green Version]
- Brandt, M.; Roth, K.; Hammes, W. Effect of an Exopolysaccharide Produced by Lactobacillus sanfranciscensis LTH1729 on Dough and Bread Quality. In Sourdough from Fundamentals to Applications; Vrije Universiteit Brussel: Brussels, Belgium, 2003; p. 80. [Google Scholar]
- Sutherland, I.W. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Christensen, S.K.; Mikkelsen, M.; Pedersen, K.; Gerdes, K. RelE, a global inhibitor of translation, is activated during nutritional stress. Proc. Natl. Acad. Sci. USA 2001, 98, 14328–14333. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, K.; Christensen, S.K.; Lobner-Olesen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef]
- Degeest, B.; De Vuyst, L. Correlation of Activities of the Enzymes α-Phosphoglucomutase, UDP-Galactose 4-Epimerase, and UDP-Glucose Pyrophosphorylase with Exopolysaccharide Biosynthesis by Streptococcus thermophilus LY03. Appl. Environ. Microbiol. 2000, 66, 3519–3527. [Google Scholar] [CrossRef] [Green Version]
- Escalante, A.; Wacher-Rodarte, C.; Garcia-Garibay, M.; Farres, A. Enzymes involved in carbohydrate metabolism and their role on exopolysaccharide production in Streptococcus thermophilus. J. Appl. Microbiol. 1998, 84, 108–114. [Google Scholar] [CrossRef]
- Grobben, G.; Smith, M.; Sikkema, J.; De Bont, J. Influence of fructose and glucose on the production of exopolysaccharides and the activities of enzymes involved in the sugar metabolism and the synthesis of sugar nucleotides in Lactobacillus delbrueckii subsp. bulgaricus NCFB 2772. Appl. Microbiol. Biotechnol. 1996, 46, 279–284. [Google Scholar] [CrossRef]
- Franz, C.M.; Vancanneyt, M.; Vandemeulebroecke, K.; De Wachter, M.; Cleenwerck, I.; Hoste, B.; Schillinger, U.; Holzapfel, W.H.; Swings, J. Pediococcus stilesii sp. nov., isolated from maize grains. Int. J. Syst. Evol. Microbiol. 2006, 56, 329–333. [Google Scholar] [CrossRef]
- Orla-Jensen, S. The Lactic Acid Bacteria; Høst: Copenhagen, Denmark, 1919; Volume 3. [Google Scholar]
- Dobson, C.M.; Deneer, H.; Lee, S.; Hemmingsen, S.; Glaze, S.; Ziola, B. Phylogenetic analysis of the genus Pediococcus, including Pediococcus claussenii sp. nov., a novel lactic acid bacterium isolated from beer. Int. J. Syst. Evol. Microbiol. 2002, 52, 2003–2010. [Google Scholar]
- Russo, P.; de Chiara, M.L.V.; Capozzi, V.; Arena, M.P.; Amodio, M.L.; Rascón, A.; Dueñas, M.T.; López, P.; Spano, G. Lactobacillus plantarum strains for multifunctional oat-based foods. LWT Food Sci. Technol. 2016, 68, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Rieder, A.; Ballance, S.; Knutsen, S.H. Viscosity based quantification of endogenous β-glucanase activity in flour. Carbohydr. Polym. 2015, 115, 104–111. [Google Scholar] [CrossRef]
- Vuyst, D.; de Ven, V. Production by and isolation of exopolysaccharides from Streptococcus thermophilus grown in a milk medium and evidence for their growth-associated biosynthesis. J. Appl. Microbiol. 1998, 84, 1059–1068. [Google Scholar] [CrossRef]
- Cerning, J.; Bouillanne, C.; Landon, M.; Desmazeaud, M. Isolation and characterization of exopolysaccharides from slime-forming mesophilic lactic acid bacteria. J. Dairy Sci. 1992, 75, 692–699. [Google Scholar] [CrossRef]
- Ricciardi, A.; Parente, E.; Crudele, M.A.; Zanetti, F.; Scolari, G.; Mannazzu, I. Exopolysaccharide production by Streptococcus thermophilus SY: Production and preliminary characterization of the polymer. J. Appl. Microbiol. 2002, 92, 297–306. [Google Scholar] [CrossRef]
- Banu, I.; Vasilean, I.; Aprodu, I. Quality evaluation of the sourdough rye breads. Ann. Univ. Dunarea Jos Galati. Fascicle VI Food Technol. 2011, 35, 94–105. [Google Scholar]
- Lotong, V.; IV, E.C.; Chambers, D.H. Determination of the sensory attributes of wheat sourdough bread. J. Sens. Stud. 2000, 15, 309–326. [Google Scholar] [CrossRef]
- Messens, W.; De Vuyst, L. Inhibitory substances produced by Lactobacilli isolated from sourdoughs—A review. Int. J. Food Microbiol. 2002, 72, 31–43. [Google Scholar] [CrossRef]
- Korakli, M.; Pavlovic, M.; Ganzle, M.G.; Vogel, R.F. Exopolysaccharide and kestose production by Lactobacillus sanfranciscensis LTH2590. Appl. Environ. Microbiol. 2003, 69, 2073–2079. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Temperature | Inoculum Size | Fermentation Time [h] | ||
|---|---|---|---|---|---|
| 16 | 20 | 24 | |||
| L. brevis TMW 1.2112 | 25 °C | 1× | 35 ± 29.7 | 104.5 ± 15 | 208.5 ± 11.2 |
| 28 °C | 1× | 72.2 ± 21.5 | 279 ± 73.3 | 245.9 ± 65 | |
| 35 °C | 1× | 131.1 ± 43.7 | 91.6 ± 48.7 | 506.2 ± 101.6 | |
| 28 °C | 1/2× | 60.6 ± 6 | 172.3 ± 12.6 | 188.7 ± 36.5 | |
| 28 °C | 2× | 21.1 ± 2.2 | 223.3 ± 47.7 | 111.1 ± 10.1 | |
| 28 °C | 1/2× L. brevis 1/2× La. plantarum | 46.5 ± 8.2 | 14.1 ± 3.5 | 43.8 ± 6 | |
| P. claussenii TMW 2.340 | 25 °C | 1× | 521.1 ± 57.4 | 222 ± 52.8 | 220.4 ±28.2 |
| 28 °C | 1× | 219.9 ± 7 | 134.5 ± 75.4 | 50.4 ± 8.5 | |
| 35 °C | 1× | 142.8 ± 18. | 224.9 ± 32.9 | 235.6 ± 25.4 | |
| 28 °C | 1/2× | 53.5 ± 27.9 | 64.5 ± 36.2 | 123.5 ± 56.4 | |
| 28 °C | 2× | 218.9 ± 31.1 | 178.4 ± 13.7 | 313.2 ± 24.9 | |
| 28 °C | 1/2× P. claussenii 1/2× La. plantarum | 12.5 ± 4.9 | 11.4 ± 11.9 | 1.7 ± 5.2 | |
| Strain | Temperature | Inoculum Size | Fermentation Time [h] | ||
|---|---|---|---|---|---|
| 16 | 20 | 24 | |||
| L. brevis TMW 1.2112 | 25 °C | 1× | 399.1 ± 39.2 | 422.4 ±56.4 | 573.6 ± 78.7 |
| 28 °C | 1× | 175.9 ± 69.4 | 391.7 ± 57.5 | 412.3 ± 47.4 | |
| 35 °C | 1× | 195.7 ± 40.9 | 230 ± 37.5 | 220.6 ±52.9 | |
| 28 °C | 1/2× | 191.3 ± 27.2 | 314.4 ± 34.6 | 363.6 ± 66.4 | |
| 28 °C | 2× | 41.2 ± 10.7 | 477 ± + 91.7 | 468.7 ± 90.2 | |
| 28 °C | 1/2× L. brevis 1/2× La. plantarum | 111.4 ± 8.8 | 99.3 ± 38.1 | 138.7 ± 37.1 | |
| P. claussenii TMW 2.340 | 25 °C | 1× | 493.6 ± 52.7 | 624.7 ± 62.8 | 623 ± 11.4 |
| 28 °C | 1× | 338.5 ± 51.5 | 402.2 ± 98.1 | 104 ± 18.1 | |
| 35 °C | 1× | 346 ± 36.3 | 514.1 ± 17.8 | 448.7 ± 5.2 | |
| 28 °C | 1/2× | 146.3 ± 78.1 | 200.8 ± 74.8 | 195.2 ± 121.1 | |
| 28 °C | 2× | 400.4 ± 117.3 | 328.8 ±71.9 | 429.5 ±129.8 | |
| 28 °C | 1/2× P. claussenii 1/2× La. plantarum | 29.8 ± 10.5 | 19.2 ± 21.1 | 26.4 ± 15.7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bockwoldt, J.A.; Fellermeier, J.; Steffens, E.; Vogel, R.F.; Ehrmann, M.A. β-Glucan Production by Levilactobacillus brevis and Pediococcus claussenii for In Situ Enriched Rye and Wheat Sourdough Breads. Foods 2021, 10, 547. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030547
Bockwoldt JA, Fellermeier J, Steffens E, Vogel RF, Ehrmann MA. β-Glucan Production by Levilactobacillus brevis and Pediococcus claussenii for In Situ Enriched Rye and Wheat Sourdough Breads. Foods. 2021; 10(3):547. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030547
Chicago/Turabian StyleBockwoldt, Julia A., Johanna Fellermeier, Emma Steffens, Rudi F. Vogel, and Matthias A. Ehrmann. 2021. "β-Glucan Production by Levilactobacillus brevis and Pediococcus claussenii for In Situ Enriched Rye and Wheat Sourdough Breads" Foods 10, no. 3: 547. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030547