
Microbial Diversity and Quality-Related Physicochemical Properties of Spicy Cabbage in Northeastern China and Their Correlation Analysis


Abstract
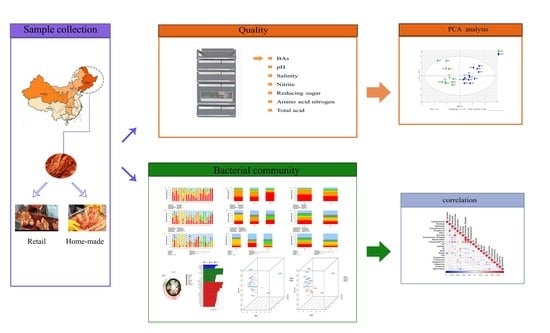
:
1. Introduction
2. Material and Methods
2.1. CSC Samples
2.2. Physicochemical Analysis
2.2.1. pH and TA Measurement
2.2.2. AAN and RS Measurement
2.2.3. Salinity and Nitrite Measurement
2.2.4. BAs Measurement
2.3. DNA Extraction and PCR Amplification for 16S rRNA Sequence
2.4. Sequence Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Physicochemical Analysis of CSC
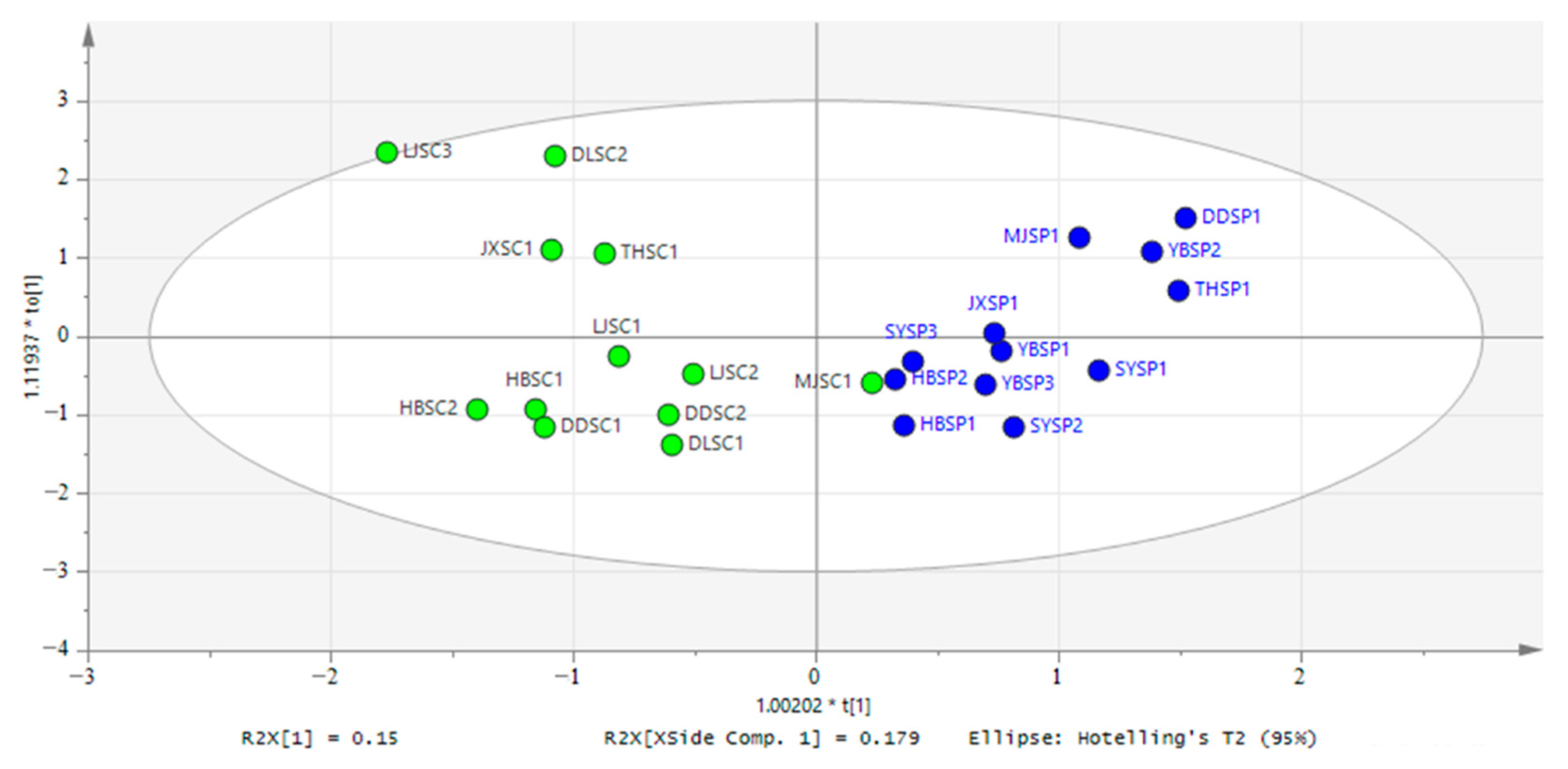
3.2. OPLS-DA of Physicochemical of CSC
3.3. Analysis of α Diversity in CSC
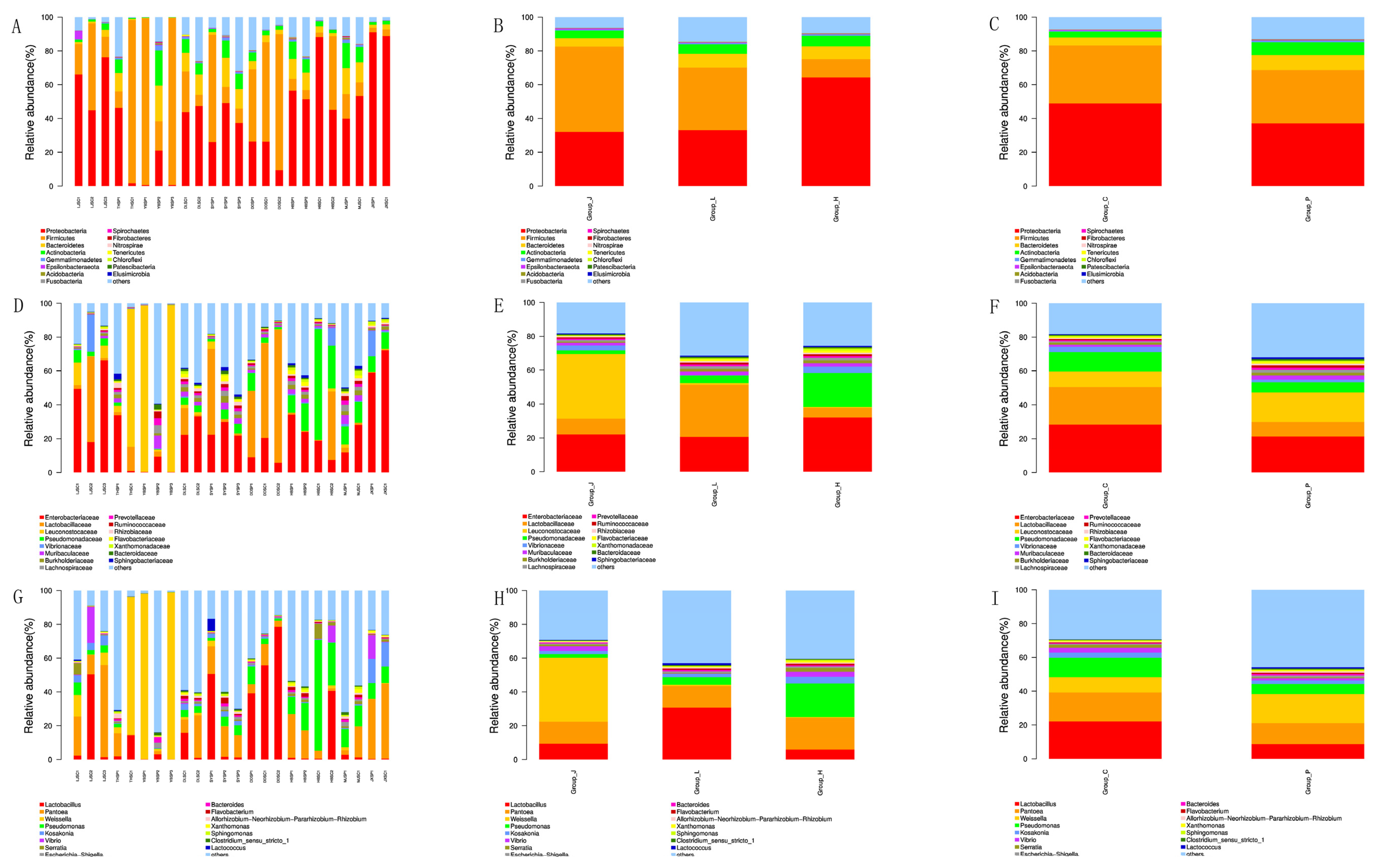
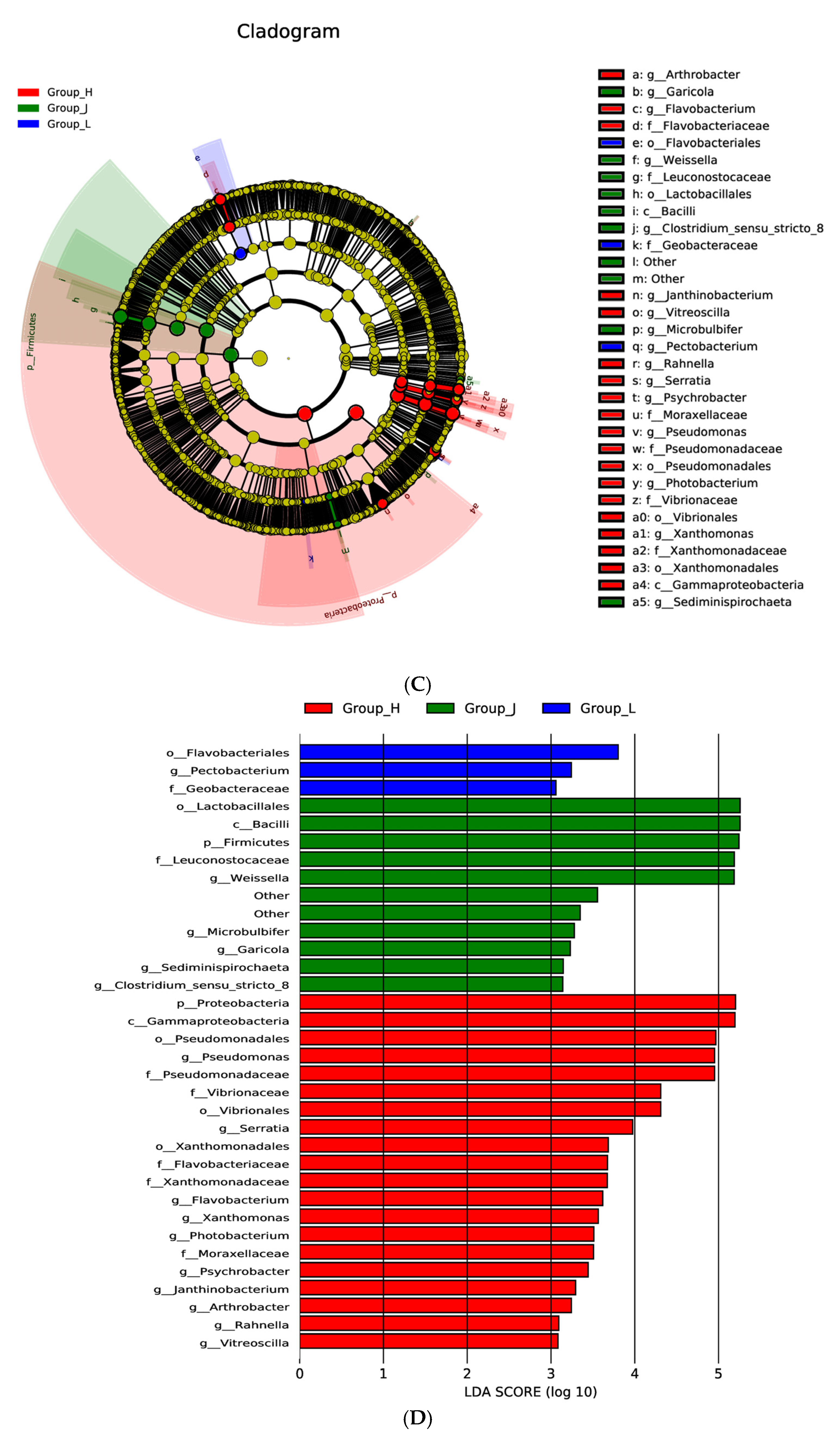
3.4. Bacterial Community of CSC
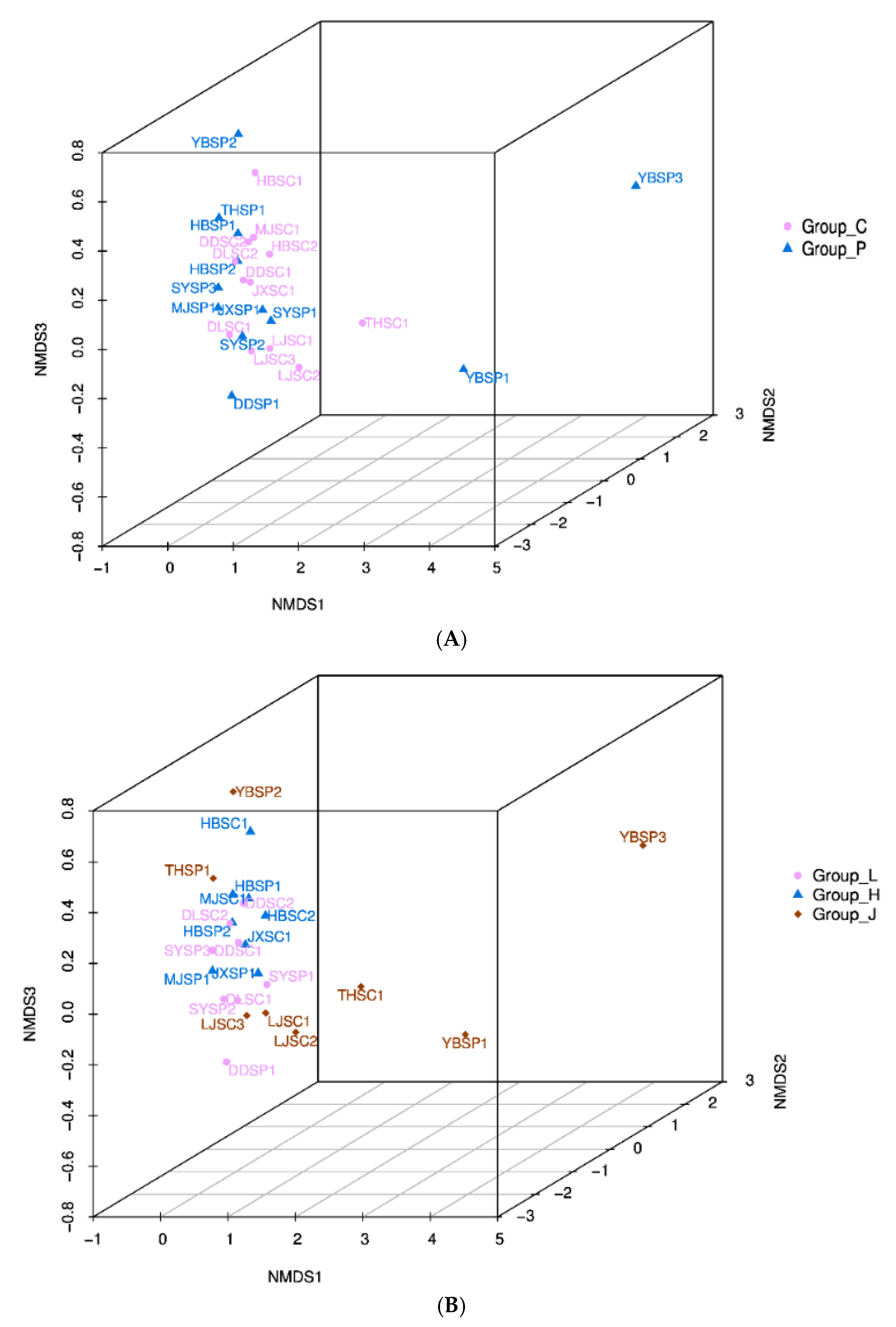
3.5. Multivariate Analysis of the CSC Bacterial Community
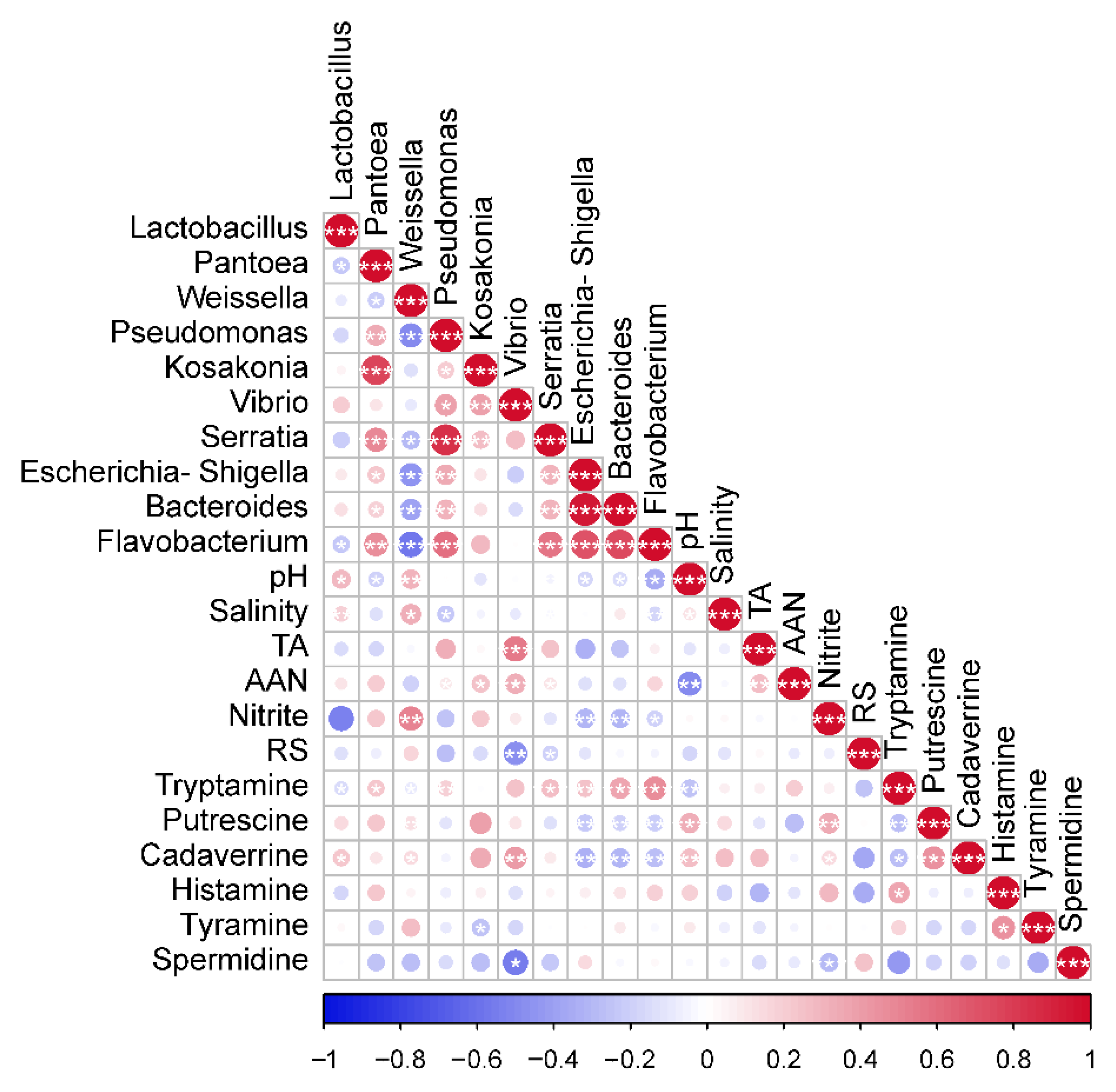
3.6. Association between Bacteria and Physicochemical Factors in CSC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, Z.; Peng, Z.; Huang, T.; Xiao, Y.; Li, J.; Xie, M.; Xiong, T. Comparison of bacterial diversity in traditionally homemade paocai and Chinese spicy cabbage. Food Microbiol. 2019, 83, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Lee, D.; Yang, C.; Jeon, J.; Kim, J.; Han, H. Microbial population dynamics of kimchi, a fermented cabbage product. FEMS Microbiol. Lett. 2006, 257, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; He, Z.; Wang, X.; Song, G.; Chen, H.; Lin, X.; Ji, C.; Zhang, S. Bacterial profiles and volatile flavor compounds in commercial Suancai with varying salt concentration from Northeastern China. Food Res. Int. 2020, 137, 109384. [Google Scholar] [CrossRef]
- Song, H.S.; Whon, T.W.; Kim, J.; Lee, S.H.; Kim, J.Y.; Kim, Y.B.; Choi, H.-J.; Rhee, J.-K.; Roh, S.W. Microbial niches in raw ingredients determine microbial community assembly during kimchi fermentation. Food Chem. 2020, 318, 126481. [Google Scholar] [CrossRef] [PubMed]
- Song, H.S.; Lee, S.H.; Ahn, S.W.; Kim, J.Y.; Rhee, J.K.; Roh, S.W. Effects of the main ingredients of the fermented food, kimchi, on bacterial composition and metabolite profile. Food Res. Int. 2021, 149, 110668. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Du, X.; Feng, L.; Mu, G.; Tuo, Y. The microbial community, biogenic amines content of soybean paste, and the degradation of biogenic amines by Lactobacillus plantarum HM24. Food Sci. Nutr. 2021, 9, 6458–6470. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hu, W.; Xiu, Z.; Jiang, A.; Yang, X.; Ji, Y.; Guan, Y.; Feng, K. Comparison of northeast sauerkraut fermentation between single lactic acid bacteria strains and traditional fermentation. Food Res. Int. 2020, 137, 109553. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hu, W.; Jiang, A.; Xiu, Z.; Ji, Y.; Guan, Y.; Yang, X. Effect of salt concentration on quality of Chinese northeast sauerkraut fermented by Leuconostoc mesenteroides and Lactobacillus plantarum. Food Biosci. 2019, 30, 100421. [Google Scholar] [CrossRef]
- Zhao, N.; Lai, H.; Wang, Y.; Huang, Y.; Shi, Q.; He, W.; Zhu, S.; Li, Y.; Zhu, Y.; Li, H. Assessment of biogenic amine and nitrite production in low-salt Paocai during fermentation as affected by reused brine and fresh brine. Food Biosci. 2021, 41, 100958. [Google Scholar] [CrossRef]
- Wang, Z.; Shao, Y. Effects of microbial diversity on nitrite concentration in pao cai, a naturally fermented cabbage product from China. Food Microbiol. 2018, 72, 185–192. [Google Scholar] [CrossRef]
- Yu, Y.; Li, L.; Xu, Y.; An, K.; Shi, Q.; Yu, Y.; Xu, Z. Evaluation of the Relationship among Biogenic Amines, Nitrite and Microbial Diversity in Fermented Mustard. Molecules 2021, 26, 6173. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-E.; Seo, S.-H.; Kim, E.-J.; Byun, S.; Na, C.-S.; Son, H.-S. Changes of microbial community and metabolite in kimchi inoculated with different microbial community starters. Food Chem. 2019, 274, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; He, Z.; Wang, X.; Zhao, M.; Cao, X.; Lin, X.; Ji, C.; Zhang, S.; Liang, H. Improving the quality of Suancai by inoculating with Lactobacillus plantarum and Pediococcus pentosaceus. Food Res. Int. 2021, 148, 110581. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yang, J.; Hou, Q.; Xu, H.; Zheng, Y.; Zhang, H.; Zhang, L. Assessment of bacterial profiles in aged, home-made Sichuan paocai brine with varying titratable acidity by PacBio SMRT sequencing technology. Food Control 2017, 78, 14–23. [Google Scholar] [CrossRef]
- Wang, X.; Song, G.; He, Z.; Zhao, M.; Cao, X.; Lin, X.; Ji, C.; Zhang, S.; Liang, H. Effects of salt concentration on the quality of paocai, a fermented vegetable product from China. J. Sci. Food Agric. 2021, 101, 6202–6210. [Google Scholar] [CrossRef] [PubMed]
- Satora, P.; Skotniczny, M.; Strnad, S.; Piechowicz, W. Chemical composition and sensory quality of sauerkraut produced from different cabbage varieties. LWT-Food Sci. Technol. 2021, 136, 110325. [Google Scholar] [CrossRef]
- Li, Q.; Kang, J.; Ma, Z.; Li, X.; Liu, L.; Hu, X. Microbial succession and metabolite changes during traditional serofluid dish fermentation. LWT-Food Sci. Technol. 2017, 84, 771–779. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, T.; Huang, C.; Hardie, J.; Peng, Z.; Xie, M.; Xiong, T. The microbial communities and flavour compounds of Jiangxi yancai, Sichuan paocai and Dongbei suancai: Three major types of traditional Chinese fermented vegetables. LWT-Food Sci. Technol. 2020, 121, 108865. [Google Scholar] [CrossRef]
- Stoll, D.A.; Müller, A.; Meinhardt, A.-K.; Dötsch, A.; Greiner, R.; Kulling, S.E.; Huch, M. Influence of salt concentration and iodized table salt on the microbiota of fermented cucumbers. Food Microbiol. 2020, 92, 103552. [Google Scholar] [CrossRef]
- Kim, E.-J.; Seo, S.-H.; Park, S.-E.; Lim, Y.-W.; Roh, S.W.; Son, H.-S. Initial storage of kimchi at room temperature alters its microbial and metabolite profiles. LWT-Food Sci. Technol. 2020, 134, 110160. [Google Scholar] [CrossRef]
- Yang, X.; Hu, W.; Xiu, Z.; Jiang, A.; Yang, X.; Ji, Y.; Guan, Y.; Feng, K. Microbial dynamics and volatilome profiles during the fermentation of Chinese northeast sauerkraut by Leuconostoc mesenteroides ORC 2 and Lactobacillus plantarum HBUAS 51041 under different salt concentrations. Food Res. Int. 2020, 130, 108926. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, Y.; Li, T.; Yang, Y.; Zeng, F.; Wang, H.; Suo, H.; Song, J.; Zhang, Y. Microbial composition and correlation between microbiota and quality-related physiochemical characteristics in chongqing radish paocai. Food Chem. 2022, 369, 130897. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Cho, Y.; Lee, Y.; Han, S.-K.; Kim, C.-G.; Choo, D.-W.; Kim, Y.-R.; Kim, H.-Y. A proteomic approach for rapid identification of Weissella species isolated from Korean fermented foods on MALDI-TOF MS supplemented with an in-house database. Int. J. Food Microbiol. 2017, 243, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Cho, E.-J.; Yang, S.-M.; Kim, M.-J.; Kim, H.-Y. Novel approaches for the identification of microbial communities in kimchi: MALDI-TOF MS analysis and high-throughput sequencing. Food Microbiol. 2021, 94, 103641. [Google Scholar] [CrossRef] [PubMed]
- Adewumi, G.A.; Oguntoyinbo, F.A.; Keisam, S.; Jeyaram, K. Combination of culture-independent and culture-dependent molecular methods for the determination of bacterial community of iru, a fermented Parkia biglobosa seeds. Front. Microbiol. 2013, 3, 436. [Google Scholar] [CrossRef] [Green Version]
- Ahmadsah, L.; Kim, E.; Jung, Y.; Kim, H. Identification by PCR-DGGE, SDS-PAGE, and MALDI-TOF MS of LAB and fungi in laru, a fermented starter. J. Microbiol. Biotechnol. 2018, 28, 32–39. [Google Scholar] [CrossRef]
- Ahmadsah, L.S.; Min, S.-G.; Han, S.-K.; Hong, Y.; Kim, H.-Y. Effect of low salt concentrations on microbial changes during kimchi fermentation monitored by PCR-DGGE and their sensory acceptance. J. Microbiol. Biotechnol. 2015, 25, 2049–2057. [Google Scholar] [CrossRef]
- Lee, M.E.; Jang, J.-Y.; Lee, J.H.; Park, H.W.; Choi, H.J.; Kim, T.W. Starter cultures for kimchi fermentation. J. Microbiol. Biotechnol. 2015, 25, 559–568. [Google Scholar] [CrossRef]
- Lee, M.; Song, J.H.; Lee, S.H.; Jung, M.Y.; Chang, J.Y. Effect of seasonal production on bacterial communities in Korean industrial kimchi fermentation. Food Control 2018, 91, 381–389. [Google Scholar] [CrossRef]
- Lee, M.; Song, J.H.; Park, J.M.; Chang, J.Y. Bacterial diversity in Korean temple kimchi fermentation. Food Res. Int. 2019, 126, 108592. [Google Scholar] [CrossRef]
- Paik, H.-D.; Lee, J.Y. Investigation of reduction and tolerance capability of lactic acid bacteria isolated from kimchi against nitrate and nitrite in fermented sausage condition. Meat Sci. 2014, 97, 609–614. [Google Scholar] [CrossRef]
- Lu, L.; Xiaoxue, W.; Zhiyou, W.; Shouwen, C.; Ling, W.; Xuetuan, W. Evaluation of the Biogenic Amines Formation and Degradation Abilities of Lactobacillus curvatus From Chinese Bacon. Front. Microbiol. 2018, 9, 1015. [Google Scholar]
- Hao, Y.; Sun, B. Analysis of bacterial diversity and biogenic amines content during fermentation of farmhouse sauce from Northeast China–ScienceDirect. Food Control 2020, 108, 106861. [Google Scholar] [CrossRef]
- Luo, F.; Yang, Z.; Zhong, K.; Huang, C.; Yu, Z.; Peng, Z.; Wu, Y.; Bu, Q.; Gao, H. Effects of Bacillus megaterium L222 on quality and bacterial diversity of Sichuan paocai. Food Res. Int. 2021, 140, 109994. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Jung, J.Y.; Jeon, C.O. Effects of temperature on microbial succession and metabolite change during saeu-jeot fermentation. Food Microbiol. 2014, 38, 16–25. [Google Scholar] [CrossRef]
- Hoang, N.X.; Ferng, S.; Ting, C.-H.; Lu, Y.-C.; Yeh, Y.-F.; Lai, Y.-R.; Chiou, R.Y.-Y.; Hwang, J.-Y.; Hsu, C.-K. Effect of initial 5 days fermentation under low salt condition on the quality of soy sauce. LWT-Food Sci. Technol. 2018, 92, 234–241. [Google Scholar] [CrossRef]
- ten Brink, B.; Damink, C.; Joosten, H.; In’t Veld, J.H. Occurrence and formation of biologically active amines in foods. Int. J. Food Microbiol. 1990, 11, 73–84. [Google Scholar] [CrossRef]
- Santos, M.S. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, K.S. Tyramine production among lactic acid bacteria and other species isolated from kimchi. LWT-Food Sci. Technol. 2014, 56, 406–413. [Google Scholar] [CrossRef]
- Jeong, D.-W.; Lee, J.-H. Antibiotic resistance, hemolysis and biogenic amine production assessments of Leuconostoc and Weissella isolates for kimchi starter development. LWT-Food Sci. Technol. 2015, 64, 1078–1084. [Google Scholar] [CrossRef]
- Kim, S.Y.; Dang, Y.M.; Ha, J.H. Effect of various seasoning ingredients on the accumulation of biogenic amines in kimchi during fermentation. Food Chem. 2022, 380, 132214. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Lee, J.H.; Mah, J.-H. Occurrence and reduction of biogenic amines in kimchi and korean fermented seafood products. Foods 2019, 8, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.-H.; Kung, H.-F.; Lin, Q.-L.; Hwang, J.-H.; Cheng, S.-H.; Wei, C.-I.; Hwang, D.-F. Occurrence of histamine and histamine-forming bacteria in kimchi products in Taiwan. Food Chem. 2005, 90, 635–641. [Google Scholar] [CrossRef]
- Meng, L.; Zhu, X.; Tuo, Y.; Zhang, H.; Li, Y.; Xu, C.; Mu, G.; Jiang, S. Reducing antigenicity of β-lactoglobulin, probiotic properties and safety evaluation of Lactobacillus plantarum AHQ-14 and Lactobacillus bulgaricus BD0390. Food Biosci. 2021, 42, 101137. [Google Scholar] [CrossRef]
- Xian, S.; Zhong, H.; Yi, B.; Liu, X.; Shen, G.; Li, M.; Zhang, Z.; Luo, Q.; Li, S.; Zhou, M.; et al. Identification of pellicle formation related microorganisms in traditional Sichuan paocai through metagenomic sequence and the effects of Baijiu/Salt on pellicle and volatile components. Food Res. Int. 2022, in press. [CrossRef]
- Guan, Q.; Zheng, W.; Huang, T.; Xiao, Y.; Liu, Z.; Peng, Z.; Gong, D.; Xie, M.; Xiong, T. Comparison of microbial communities and physiochemical characteristics of two traditionally fermented vegetables. Food Res. Int. 2020, 128, 108755. [Google Scholar] [CrossRef]
- Jung, M.Y.; Kim, T.-W.; Lee, C.; Kim, J.Y.; Song, H.S.; Kim, Y.B.; Ahn, S.W.; Kim, J.S.; Roh, S.W.; Lee, S.H. Role of jeotgal, a Korean traditional fermented fish sauce, in microbial dynamics and metabolite profiles during kimchi fermentation. Food Chem. 2018, 265, 135–143. [Google Scholar] [CrossRef]
- Luo S-m Liu S-l Du X-h Chen, G.; Yan, Z. Study on microbial communities in Sichuan pickle and the correlation between salinity or acidity and microbial communities. Food Ferment. Ind. 2013, 39, 29–34. [Google Scholar]
- Chung, Y.B.; Lee, H.; Hwang, S.; Seo, H.Y.; Suh, H.J.; Jo, K. Effect of capsaicinoids in hot pepper powder on microbial community and free sugar during kimchi fermentation. J. Food Sci. 2021, 86, 3195–3204. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Jeon, C.O. Microbial succession and metabolite changes during fermentation of saeu-jeot: Traditional Korean salted seafood. Food Microbiol. 2013, 34, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Yang, H.S.; Li, J.; Han, S.K.; Chang, H.C.; Kim, H.Y. Identification of lactic acid bacteria in salted Chinese cabbage by SDS-PAGE and PCR-DGGE. J. Sci. Food Agric. 2014, 94, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.D.; Fletcher, G.C.; Paturi, G.; Hedderley, D.I. Influence of farming methods and seawater depth on Vibrio species in New Zealand Pacific oysters. Int. J. Food Microbiol. 2020, 325, 108644. [Google Scholar] [CrossRef]
- Cilli, F.; Nazli-Zeka, A.; Arda, B.; Sipahi, O.R.; Aksit-Barik, S.; Kepeli, N.; Ozinel, M.A.; Gulay, Z.; Ulusoy, S. Serratia marcescens sepsis outbreak caused by contaminated propofol. Am. J. Infect. Control 2019, 47, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Ersoz, G.; Uguz, M.; Aslan, G.; Horasan, E.; Kaya, A. Outbreak of meningitis due to Serratia marcescens after spinal anaesthesia. J. Hosp. Infect. 2014, 87, 122–125. [Google Scholar] [CrossRef]
- Devi, K.R.; Srinivasan, S.; Ravi, A.V. Inhibition of quorum sensing-mediated virulence in Serratia marcescens by Bacillus subtilis R-18. Microb. Pathog. 2018, 120, 166–175. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Smollan, G.; Tal, I.; Zade, N.P.; Haviv, Y.; Nudelman, V.; Gal-Mor, O.; Jaber, H.; Zimlichman, E.; Keller, N. Sink traps as the source of transmission of OXA-48–producing Serratia marcescens in an intensive care unit. Infect. Control Hosp. Epidemiol. 2018, 39, 1307–1315. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Luo, F.; Zhong, K.; Huang, C.; Yu, Z.; Peng, Z.; Wu, Y.; Bu, Q.; Gao, H. Effect of Bacillus subtilis Y61 inoculation on bacterial community and metabolic profile of sichuan paocai fermentation. LWT-Food Sci. Technol. 2021, 137, 110393. [Google Scholar] [CrossRef]
- He, Z.; Chen, H.; Wang, X.; Lin, X.; Ji, C.; Li, S.; Liang, H. Effects of different temperatures on bacterial diversity and volatile flavor compounds during the fermentation of suancai, a traditional fermented vegetable food from northeastern China. LWT-Food Sci. Technol. 2020, 118, 108773. [Google Scholar] [CrossRef]
- Huang, T.-T.; Wu, Z.-Y.; Zhang, W.-X. Effects of garlic addition on bacterial communities and the conversions of nitrate and nitrite in a simulated pickle fermentation system. Food Control 2020, 113, 107215. [Google Scholar] [CrossRef]
- Semenova, E.M.; Ershov, A.P.; Sokolova, D.S.; Tourova, T.P.; Nazina, T.N. Diversity and Biotechnological Potential of Nitrate-Reducing Bacteria from Heavy-Oil Reservoirs (Russia). Microbiology 2020, 89, 685–696. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Type | Sampling Province | Sampling Location | pH | Salinity (g/100 g) | AAN (g/100 g) | TA (g/100 g) | Nitrite (mg/kg) | RS (mg/g) |
|---|---|---|---|---|---|---|---|---|---|
| LJSC1 | homemade | Jilin | Linjiang | 5.36 ± 0.03 c | 4.32 ± 0.06 a | 0.136 ± 0.007 e,f,g | 0.21 ± 0.01 i | 2.00 ± 0.18 f | 3.24 ± 0.16 k,l |
| LJSC2 | homemade | Jilin | Linjiang | 4.75 ± 0.03 g | 2.43 ± 0.24 h | 0.150 ± 0.01 d,e | 0.23 ± 0.02 g,h | 2.79 ± 0.12 e | 1.16 ± 0.02 o |
| LJSC3 | homemade | Jilin | Linjiang | 5.87 ± 0.02 a | 2.98 ± 0.01 e,f | 0.053 ± 0.001 l | 0.09 ± 0.01 k | 4.61 ± 0.09 a | 7.72 ± 0.24 b |
| THSC1 | homemade | Jilin | Linjiang | 5.08 ± 0.03 e | 1.96 ± 0.02 m | 0.057 ± 0.006 j,k | 0.19 ± 0.01 i | 0.57 ± 0.04 k | 0.47 ± 0.011 p |
| THSP1 | retail | Jilin | Tonghua | 4.81 ± 0.03 f | 1.99 ± 0.02 l,m | 0.095 ± 0.000 i | 0.32 ± 0.01 f | 4.04 ± 0.14 k,l | 3.35 ± 0.08 k,l |
| YBSP1 | retail | Jilin | Yanji | 4.13 ± 0.03 m | 1.93 ± 0.06 m,n | 0.127 ± 0.000 g | 0.14 ± 0.001 j | 0.72 ± 0.11 i,j,k | 6.54 ± 0.04 d |
| YBSP2 | retail | Jilin | Yanji | 4.65 ± 0.06 i | 1.73 ± 0.03 o,p | 0.061 ± 0.001 k,l | 0.12 ± 0 j,k | 0.63 ± 0.05 j,k,l | 4.05 ± 0.02 g,h |
| YBSP3 | retail | Jilin | Yanji | 4.33 ± 0.03 k | 2.08 ± 0.01 j,k,l,m | 0.160 ± 0.004 i,j | 0.25 ± 0.01 g | 0.95 ± 0.04 h,i | 3.70 ± 0.15 i,j |
| DLSC1 | homemade | Liaoning | Dandong | 3.93 ±0.02 n | 3.31 ± 0.05 c | 0.074 ± 0.001 j,k | 0.47 ± 0.09 c | 1.96 ± 0.15 f | 5.05 ± 0.07 f |
| DLSC2 | homemade | Liaoning | Dandong | 4.74 ± 0.06 g,h | 1.65 ± 0.02 p | 0.092 ± 0.005 i | 0.34 ± 0.03 e,f | 2.88 ± 0.21 e | 6.77 ± 0.08 c |
| DDSC1 | homemade | Liaoning | Dalian | 4.27 ± 0.04 l | 1.81 ± 0.02 n,o | 0.137 ± 0.005 i | 0.52 ± 0.037 b | 4.30 ± 0.16 b | 3.18 ± 0.21 l |
| DDSC2 | homemade | Liaoning | Dalian | 4.15 ± 0.03 m | 1.70 ± 0.03 o,p | 0.131 ± 0.003 i,j | 0.44 ± 0.01 c | 3.44 ± 0.01 d | 4.01 ± 0.08 g,h |
| SYSP1 | retail | Liaoning | Shenyang | 4.24 ± 0.02 l | 2.14 ± 0.01 j,k,l | 0.098 ± 0.011 h,i | 0.39 ± 0.02 d | 0.62 ± 0.08 j,k,l | 4.84 ± 0.06 f |
| SYSP2 | retail | Liaoning | Shenyang | 4.39 ±0.02 j,k | 3.18 ± 0.01 c,d | 0.085 ± 0.003 e,f,g | 0.60 ± 0.03 a | 0.56 ± 0.09 k,l | 3.30 ± 0.16 k,l |
| SYSP3 | retail | Liaoning | Shenyang | 4.69 ± 0.02 h,i | 2.92 ± 0.01 f | 0.112 ± 0.008 g | 0.37 ± 0.03 d,e | 1.01 ± 0.02 h | 4.11 ± 0.04 g |
| DDSP1 | retail | Liaoning | Dandong | 5.19 ± 0.08 d | 2.29 ± 0.17 i | 0.057 ± 0006 l | 0.19 ± 0.01 i | 0.42 ± 0.04 l | 3.86 ± 0.27 h,i |
| HBSC1 | homemade | Heilongjiang | Haerbin | 4.37 ± 0.03 j,k | 3.56 ± 0.15 c | 0.091 ± 0.010 h | 0.12 ± 0.05 j,k | 2.19 ± 0.06 f | 3.97 ± 0.12 g,h |
| HBSC2 | homemade | Heilongjiang | Haerbin | 4.41 ± 0.02 j | 3.27 ± 0.21 c | 0.083 ± 0.014 f | 0.14 ± 0.001 j | 3.82 ± 0.18 c | 2.11 ± 0.10 m |
| JXSC1 | homemade | Heilongjiang | Jixi | 5.04 ± 0.03 e | 2.03 ± 0.01 k,l,m | 0.230 ± 0.019 b | 0.15 ± 0.01 j | 2.67 ± 0.28 e | 15.35 ± 0.19 a |
| MJSC1 | homemade | Heilongjiang | Mudanjiang | 5.53 ± 0.04 b | 2.17 ± 0.02 i,j,k | 0.343 ± 0.004 a | 0.15 ± 0.01 j | 0.64 ± 0.03 j,k,l | 3.46 ± 0.19 j,k |
| JXSP1 | retail | Heilongjiang | Mudanjiang | 4.70 ± 0.02 g,h,i | 3.11 ± 0.01 d,e | 0.175 ± 0.003 c | 0.25 ± 0.02 g | 1.48 ± 0.04 g | 5.28 ± 0.10 e |
| MJSP1 | retail | Heilongjiang | Jixi | 5.09 ± 0.03 e | 2.21 ± 0.07 i,j | 0.061 ± 0.003 k,l | 0.14 ± 0.02 j | 1.26 ± 0.03 g | 1.85 ± 0.01 n |
| HBSP1 | retail | Heilongjiang | Haerbin | 4.35 ± 0.04 j,k | 2.69 ± 0.05 g | 0.145 ± 0.023 d,e,f | 0.42 ± 0.06 c | 1.44 ± 0.02 g | 3.45 ± 0.04 j,k |
| HBSP2 | retail | Heilongjiang | Haerbin | 4.66 ± 0.03 i | 2.75 ± 0.01 g | 0.133 ± 0.023 f,g | 0.35 ± 0.03 ef | 0.88 ± 0.04 h,i,g | 3.48 ± 0.06 j,k |
| Sample | Tryptamine | Phenethylamine | Putrescine | Cadaverrine | Histamine | Tyramine | Spermidine | Spermine | Total Amine |
|---|---|---|---|---|---|---|---|---|---|
| LJSC1 | ND | ND | ND | 0.76 ± 0.06 a,b | ND | ND | ND | ND | 0.76 ± 0.06 h,i |
| LJSC2 | ND | ND | 0.34 ± 0.03 b | 1.02 ± 0.37 a | ND | ND | ND | ND | 1.36 ± 0.40 f,g |
| LJSC3 | ND | ND | 0.74 ± 0.004 a | ND | ND | ND | ND | ND | 0.74 ± 0.00 h,i |
| THSC1 | 4.83 ± 0.11 c | ND | ND | ND | ND | 2.31 ± 0.2 b | ND | ND | 7.13 ± 0.14 c |
| THSP1 | ND | ND | ND | ND | ND | ND | 0.017 ± 0.00 c | ND | 0.02 ± 0.00 j |
| YBSP1 | ND | ND | ND | ND | ND | ND | 0.019 ± 0.00 c | ND | 0.02 ± 0.00 j |
| YBSP2 | 0.95 ± 0.49 e,f | ND | ND | ND | ND | ND | ND | ND | 0.95 ± 0.49 h |
| YBSP3 | ND | ND | ND | ND | ND | 1.41 ± 0.01 d | ND | ND | 1.41 ± 0.01 g,h |
| DLSC1 | 9.38 ± 0.247 b | ND | ND | ND | ND | ND | ND | ND | 9.38 ± 0.24 b |
| DLSC2 | 10.55 ± 1.44 a | ND | ND | ND | 36.53 ± 0.173 a | 3.07 ± 1.45 a | ND | ND | 50.15 ± 0.2 a |
| DDSC1 | 0.04 ± 0.024 f,g | ND | ND | ND | ND | ND | ND | ND | 0.04 ± 0.02 i |
| DDSC2 | ND | ND | ND | ND | ND | ND | 0.018 ± 0.00 c | ND | 0.02 ± 0.00 j |
| SYSP1 | ND | ND | ND | ND | ND | ND | 0.038 ± 0.01 a | ND | 0.04 ± 0.09 |
| SYSP2 | ND | ND | ND | ND | ND | ND | 0.016 ± 0.00 c | ND | 0.16 ± 0.00 j |
| SYSP3 | ND | ND | ND | ND | ND | 1.76 ± 0.03 b,c | ND | ND | 1.76 ± 0.03 e |
| DDSP1 | ND | ND | ND | ND | ND | ND | 0.032 ± 0.01 b | ND | 0.03 ± 0.01 j |
| HBSC1 | 0.82 ± 0.11 e,f | ND | ND | ND | ND | ND | ND | ND | 0.817 ± 0.01 h |
| HBSC2 | 0.02 ± 0.002 e | ND | ND | ND | ND | ND | 0.020 ± 0.03 c | ND | 0.04 ± 0.10 j |
| JXSC1 | 0.42 ± 0.04 f,g | ND | ND | ND | ND | ND | ND | ND | 0.42 ± 0.04 i |
| MJSC1 | 0.37 ± 0.06 f,g | ND | ND | ND | ND | 1.06 ± 0.01 d | ND | ND | 1.44 ± 0.01 e,f |
| JXSP1 | ND | ND | ND | ND | ND | 1.41 ± 0.11 c,d | ND | ND | 1.41 ± 0.11 f |
| MJSP1 | 2.27 ± 0.27 d | ND | ND | ND | ND | ND | ND | ND | 2.27 ± 0.27 d |
| HBSP1 | 1.33 ± 0.09 e | ND | ND | ND | ND | ND | ND | ND | 1.33 ± 0.10 f,g |
| HBSP2 | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| Samples | Clean Tags | Valid Tags | OTU | Goods Coverage | Chao1 | Shannon | Observed Species | Simpson |
|---|---|---|---|---|---|---|---|---|
| LJSC1 | 73,883 | 66,102 | 948 | 1.00 | 1146.65 | 4.96 | 920.20 | 0.92 |
| LJSC2 | 73,442 | 67,242 | 790 | 1.00 | 1070.54 | 2.94 | 750.50 | 0.70 |
| LJSC3 | 68,440 | 63,119 | 1228 | 1.00 | 1349.10 | 4.30 | 1213.00 | 0.73 |
| THSC1 | 71,936 | 67,339 | 624 | 1.00 | 943.30 | 1.21 | 584.70 | 0.32 |
| THSP1 | 71,804 | 66,850 | 1383 | 1.00 | 1536.54 | 5.77 | 1360.9 | 0.9 |
| YBSP1 | 60,868 | 59,141 | 400 | 1.00 | 764.40 | 0.28 | 399.70 | 0.04 |
| YBSP2 | 71,639 | 65,163 | 1284 | 1.00 | 1401.83 | 8.46 | 1271.5 | 0.98 |
| YBSP3 | 73,123 | 71,361 | 346 | 1.00 | 682.16 | 0.20 | 308.50 | 0.03 |
| DLSC1 | 71,531 | 65,072 | 1413 | 1.00 | 1527.51 | 7.38 | 1398.70 | 0.97 |
| DLSC2 | 72,021 | 65,941 | 1270 | 1.00 | 1452.50 | 5.76 | 1251.50 | 0.89 |
| DDSC1 | 72,149 | 65,970 | 1291 | 0.99 | 1492.86 | 3.52 | 1259.00 | 0.67 |
| DDSC2 | 71,607 | 67,690 | 1100 | 1.00 | 1239.84 | 2.23 | 1070.60 | 0.38 |
| SYSP1 | 71,279 | 66,859 | 931 | 1.00 | 1098.90 | 3.34 | 901.40 | 0.72 |
| SYSP2 | 72,406 | 66,156 | 1294 | 1.00 | 1419.27 | 7.5 | 1278.30 | 0.96 |
| SYSP3 | 70,810 | 65,458 | 1406 | 1.00 | 1571.20 | 6.23 | 1387.20 | 0.88 |
| DDSP1 | 71,945 | 67,382 | 1160 | 1.00 | 1266.26 | 4.66 | 1141.60 | 0.81 |
| HBSC1 | 71,905 | 65,058 | 1041 | 1.00 | 1197.20 | 3.58 | 1021.10 | 0.66 |
| HBSC2 | 71,596 | 64,107 | 934 | 1.00 | 1122.21 | 3.40 | 915.60 | 0.77 |
| JXSC1 | 73,590 | 65,350 | 1157 | 0.99 | 1371.42 | 4.71 | 1127.30 | 0.83 |
| MJSC1 | 71,864 | 65,526 | 1309 | 1.00 | 1434.70 | 6.64 | 1294.80 | 0.94 |
| JXSP1 | 73,422 | 63,327 | 961 | 0.99 | 1233.36 | 4.65 | 938.30 | 0.87 |
| MJSP1 | 70,959 | 64,331 | 1298 | 1.00 | 1492.25 | 7.89 | 1285.30 | 0.98 |
| HBSP1 | 72,175 | 66,277 | 1420 | 1.00 | 1632.38 | 6.74 | 1396.60 | 0.94 |
| HBSP2 | 72,303 | 65,943 | 1349 | 1.00 | 1522.56 | 6.14 | 1330.50 | 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, L.; Mu, G.; Wang, M.; Zhao, T.; Tuo, Y.; Zhu, X.; Qian, F. Microbial Diversity and Quality-Related Physicochemical Properties of Spicy Cabbage in Northeastern China and Their Correlation Analysis. Foods 2022, 11, 1511. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101511
Chang L, Mu G, Wang M, Zhao T, Tuo Y, Zhu X, Qian F. Microbial Diversity and Quality-Related Physicochemical Properties of Spicy Cabbage in Northeastern China and Their Correlation Analysis. Foods. 2022; 11(10):1511. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101511
Chicago/Turabian StyleChang, Lixuan, Guangqing Mu, Mingxu Wang, Tong Zhao, Yanfeng Tuo, Xuemei Zhu, and Fang Qian. 2022. "Microbial Diversity and Quality-Related Physicochemical Properties of Spicy Cabbage in Northeastern China and Their Correlation Analysis" Foods 11, no. 10: 1511. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101511