Comparison of the Glycolytic and Alcoholic Fermentation Pathways of Hanseniaspora vineae with Saccharomyces cerevisiae Wine Yeasts


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Fermentation in Natural Grape Must
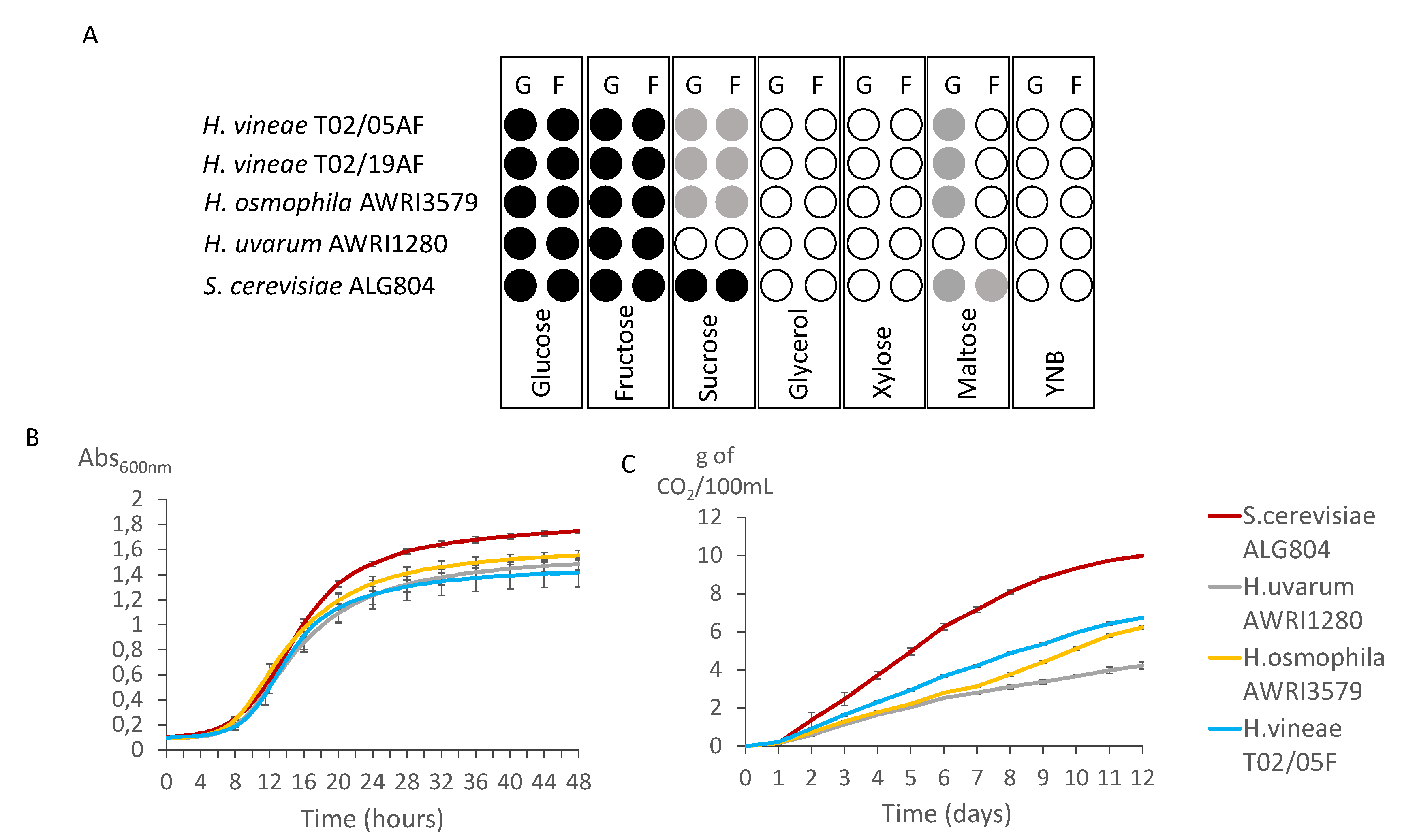
2.3. Growth Kinetics in Different Types of Media
2.4. Fermentation Ability in Different Carbon Sources
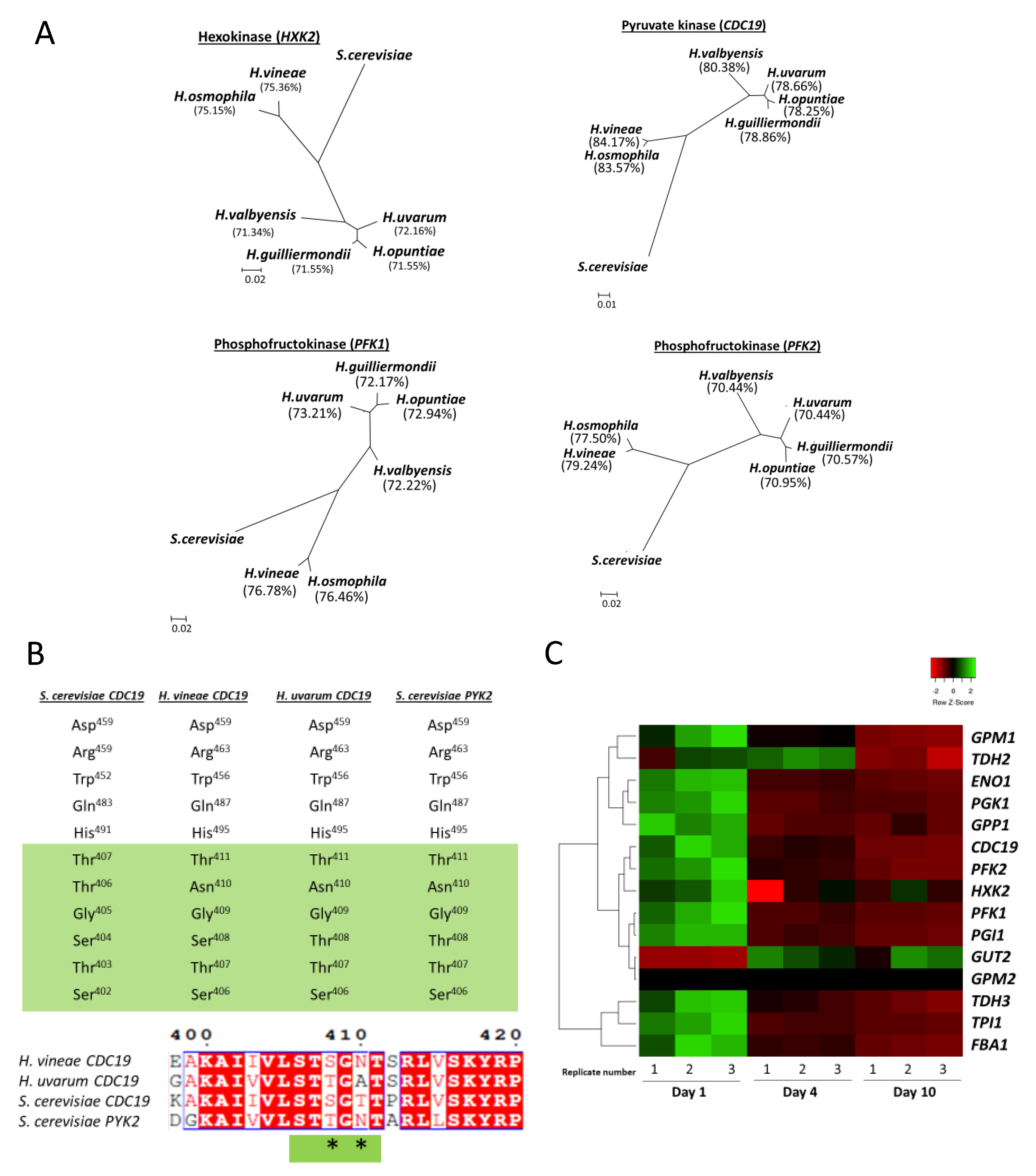
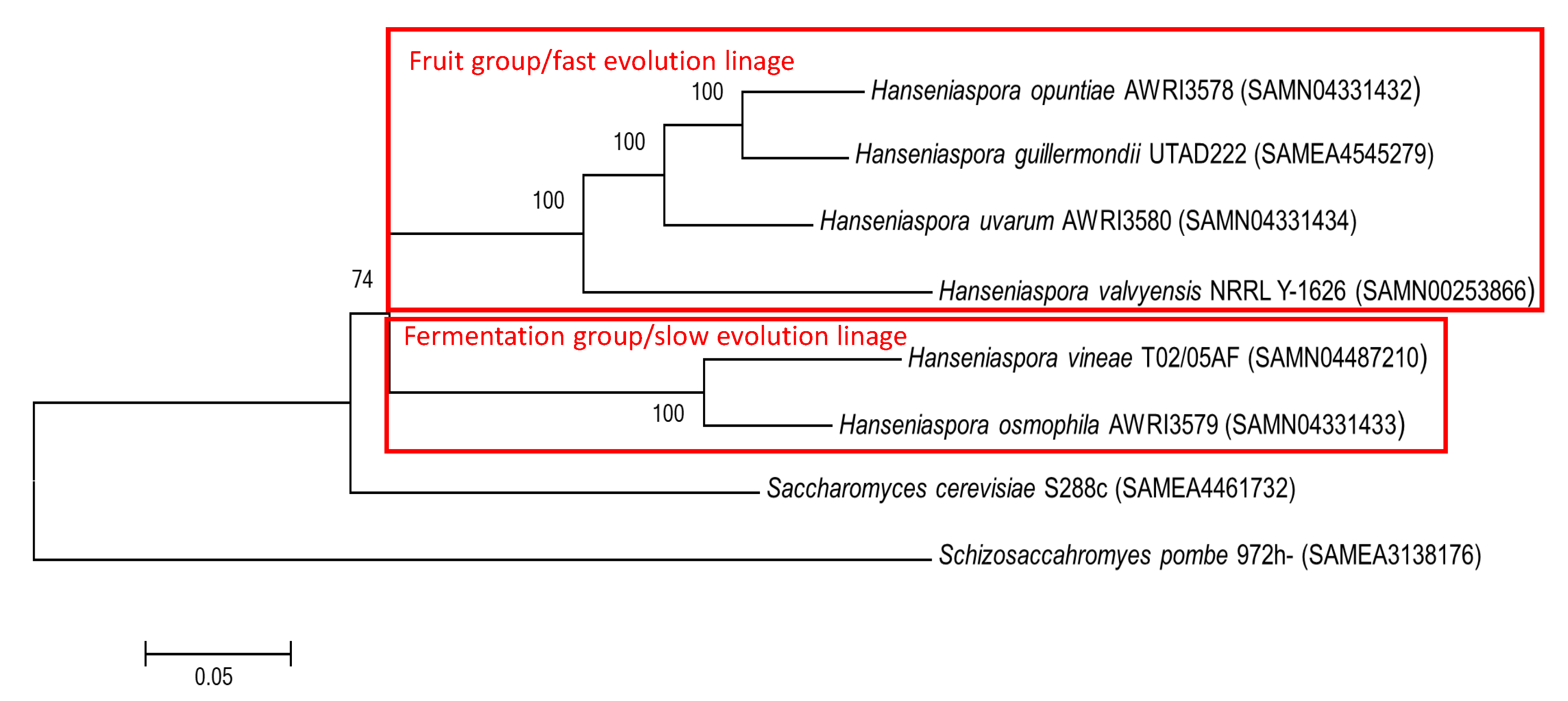
2.5. Genomic Analysis
2.6. Transcriptomic Analysis
2.7. Statistical Analysis
3. Results and Discussion
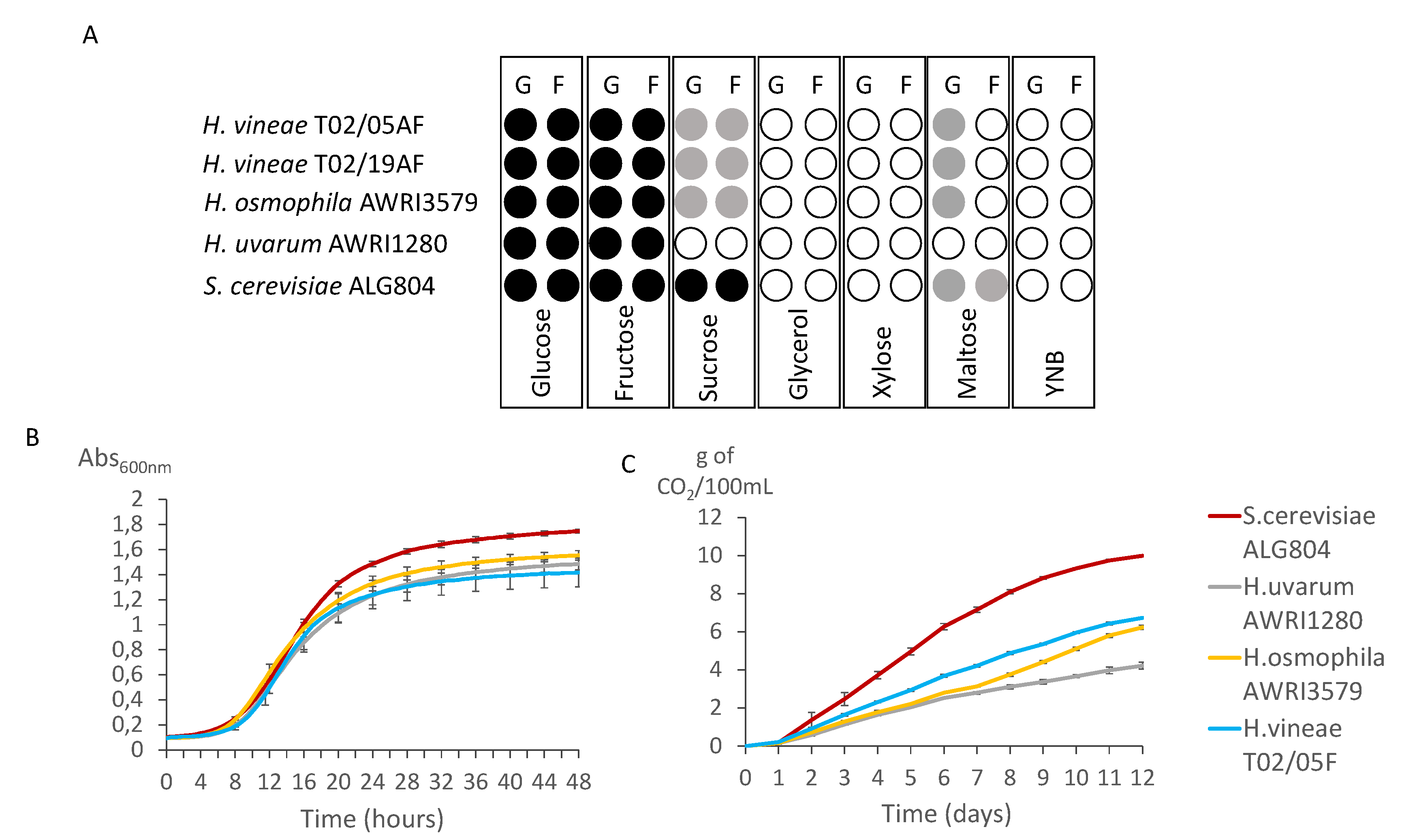
3.1. Fermentative Capacity of H. vineae in Different Media
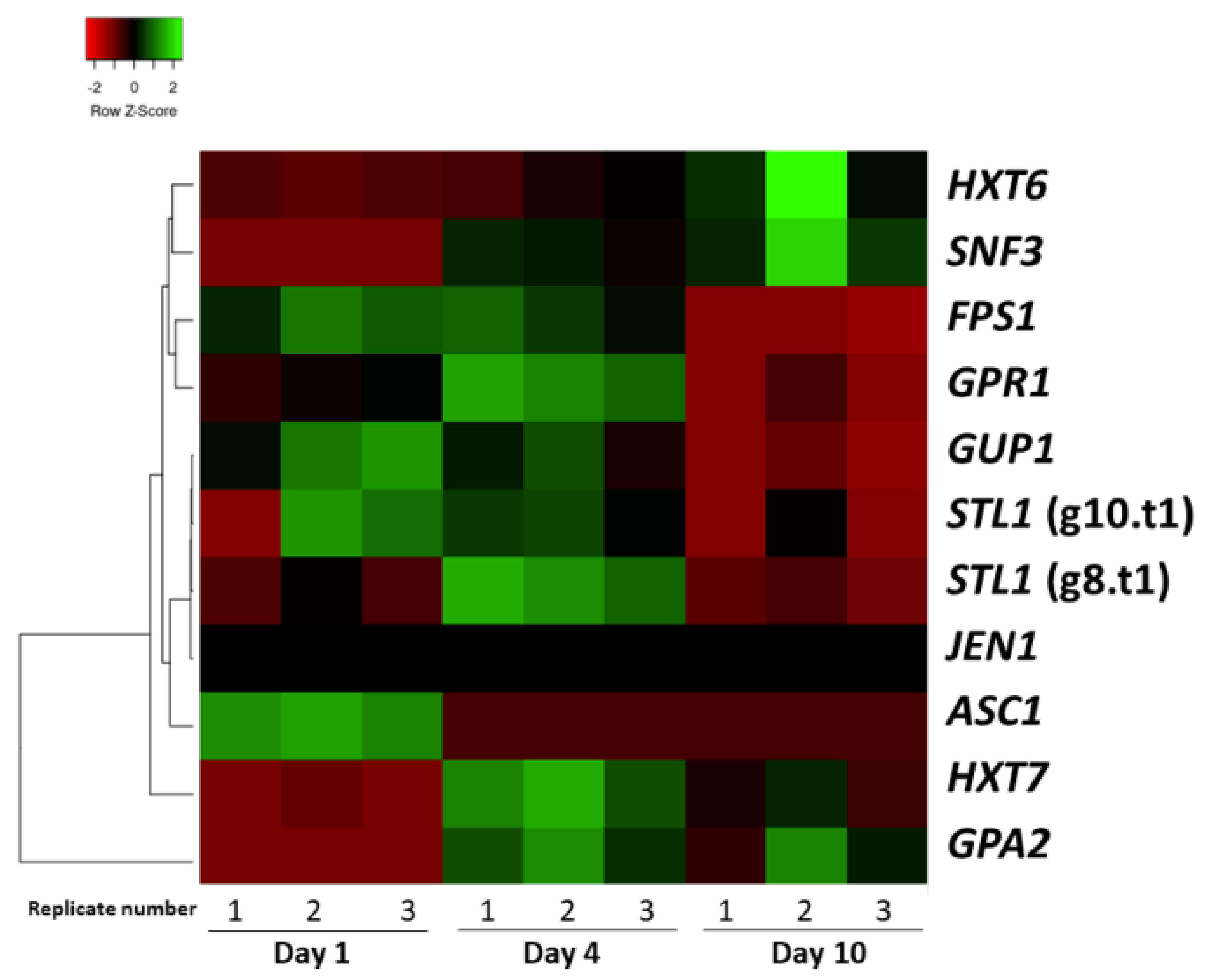
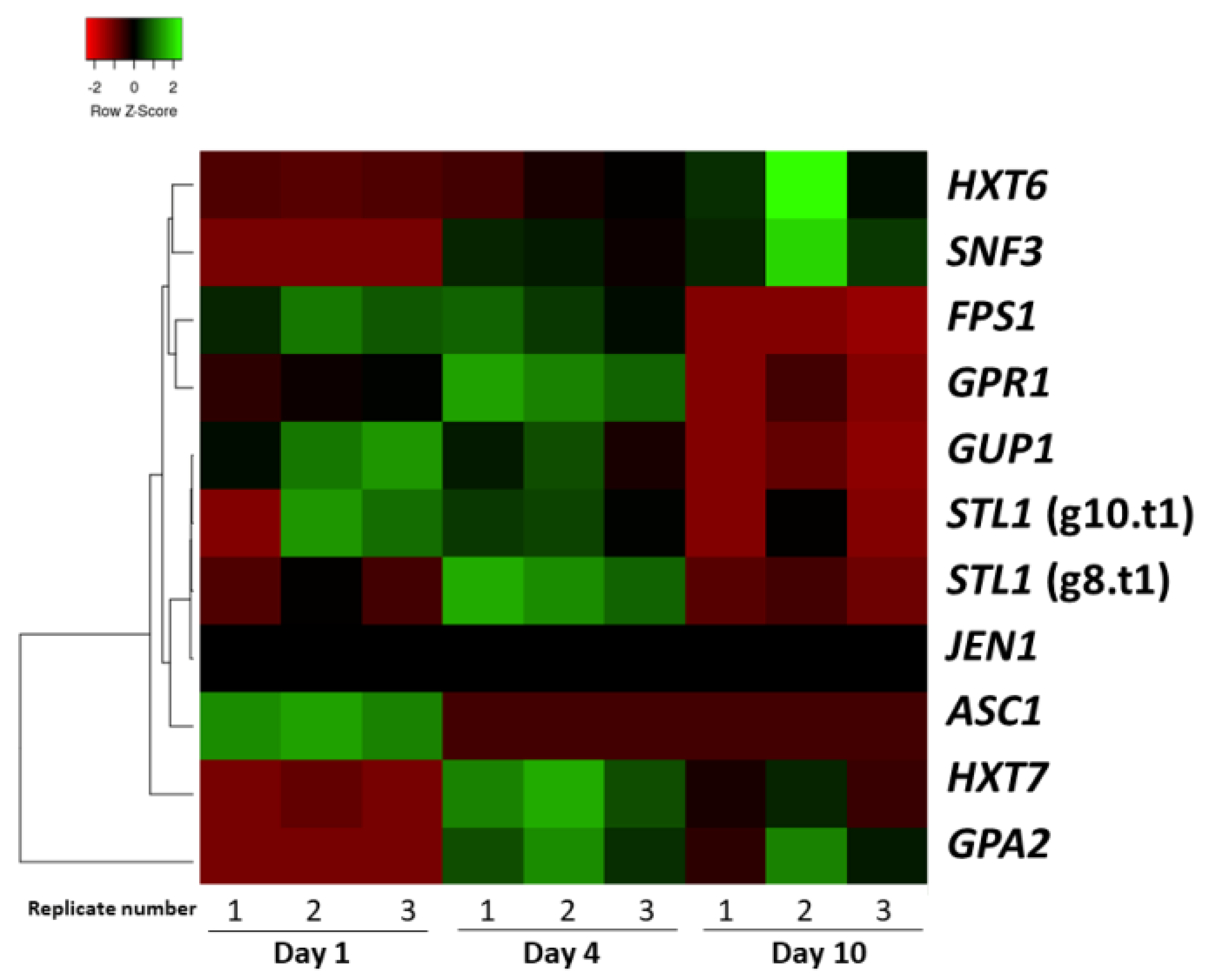
3.2. Sugar Transport
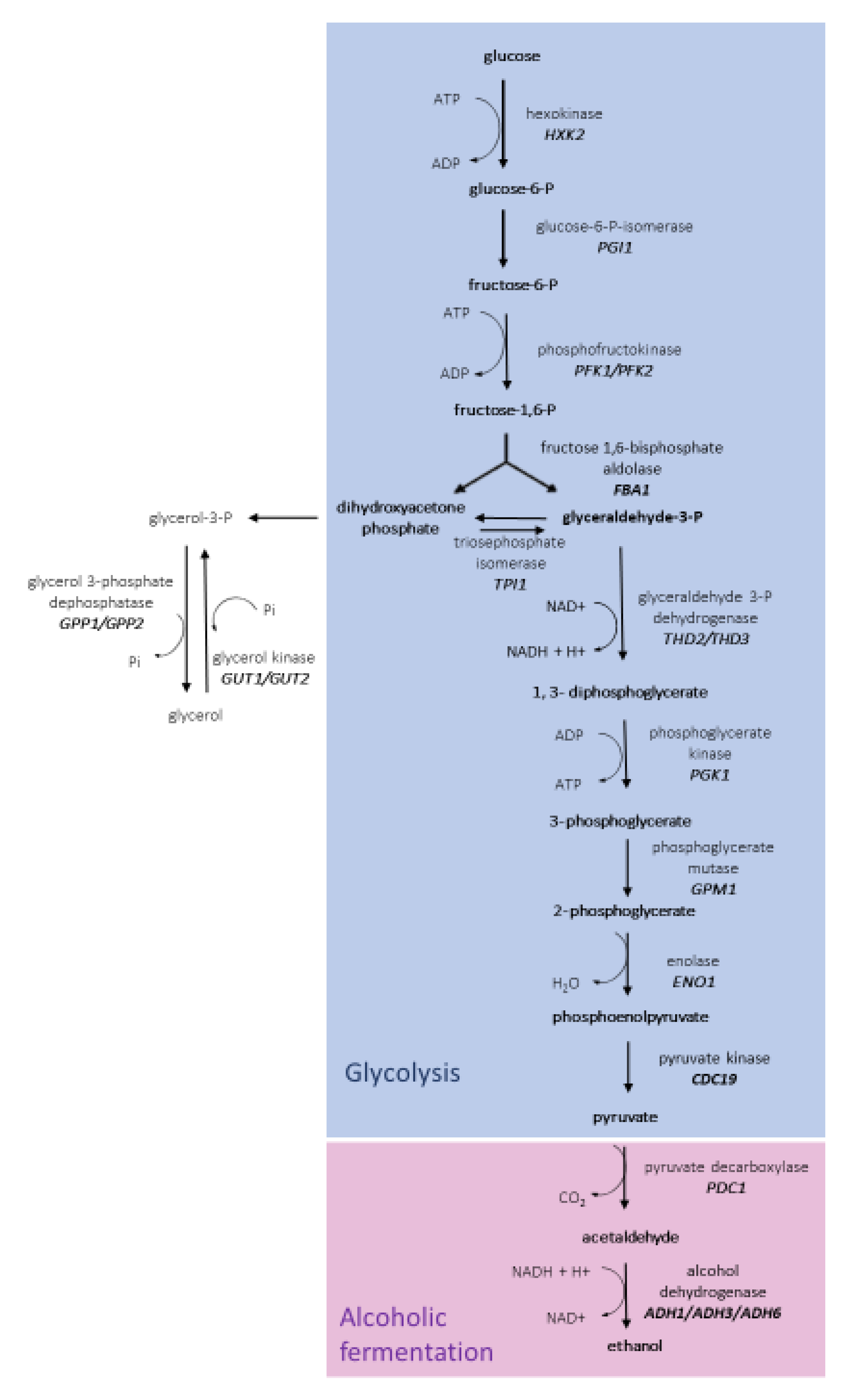
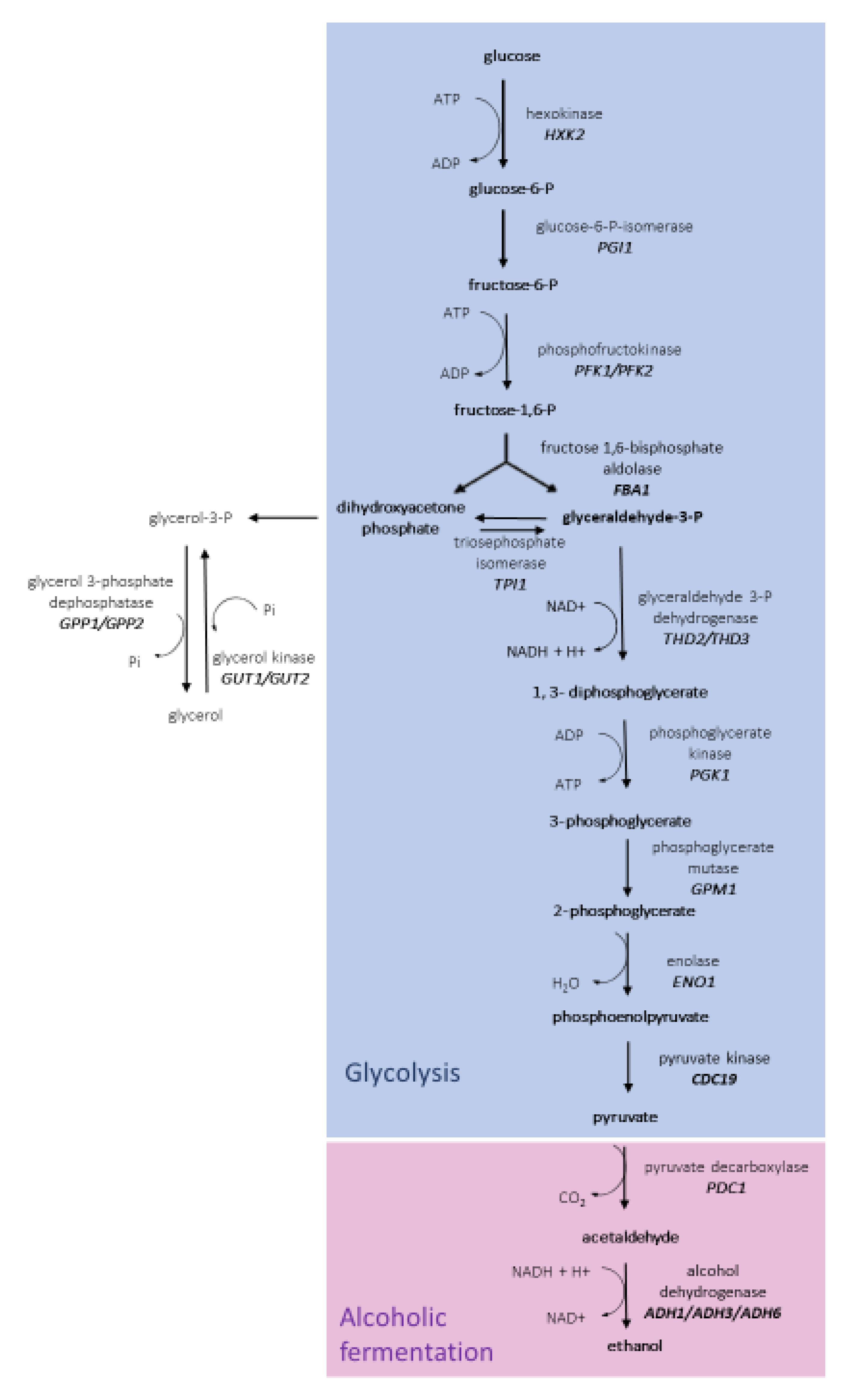
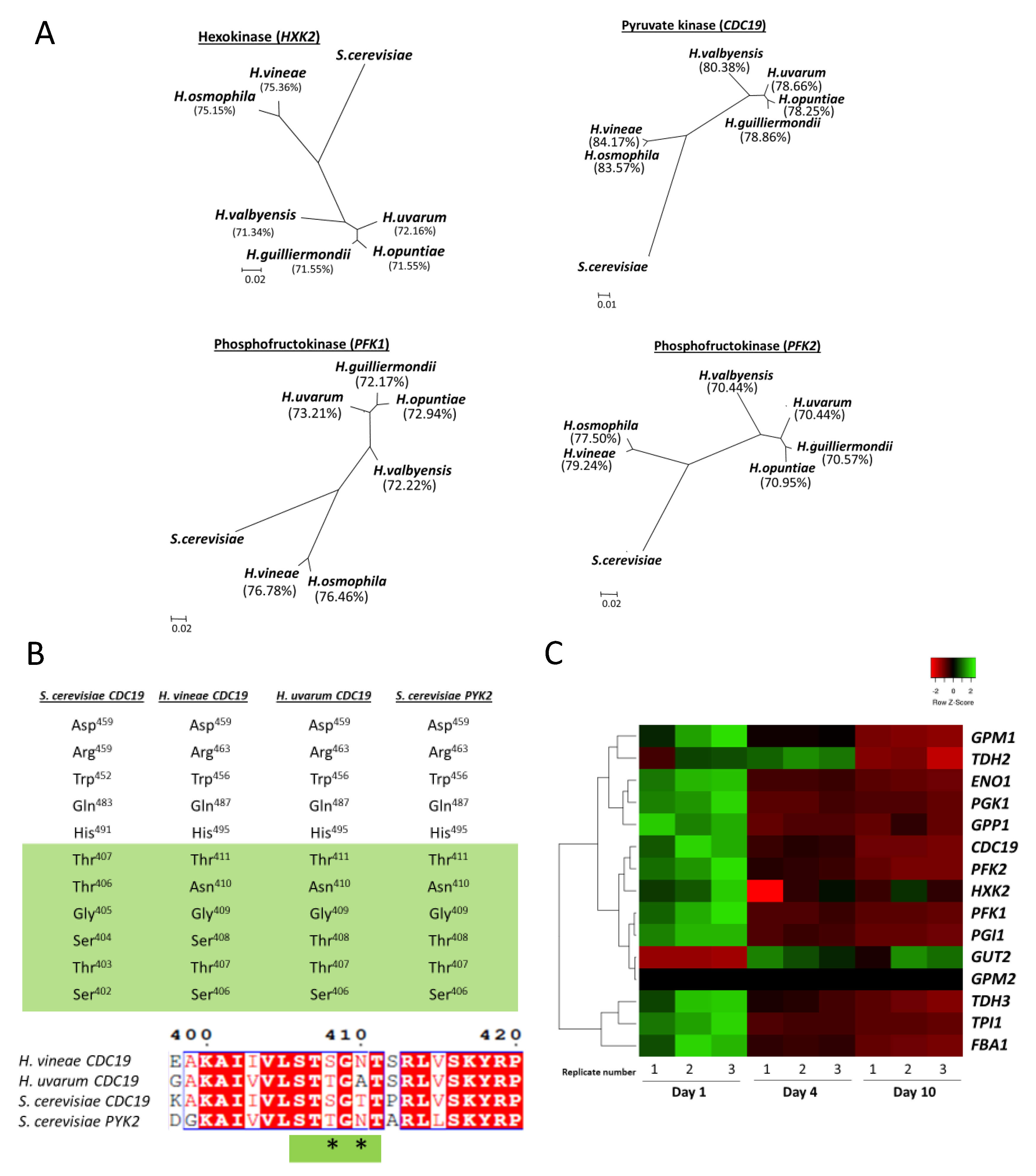
3.3. Glycolytic Pathway in H. vineae Strains
3.4. Alcoholic Fermentation in H. vineae Strains
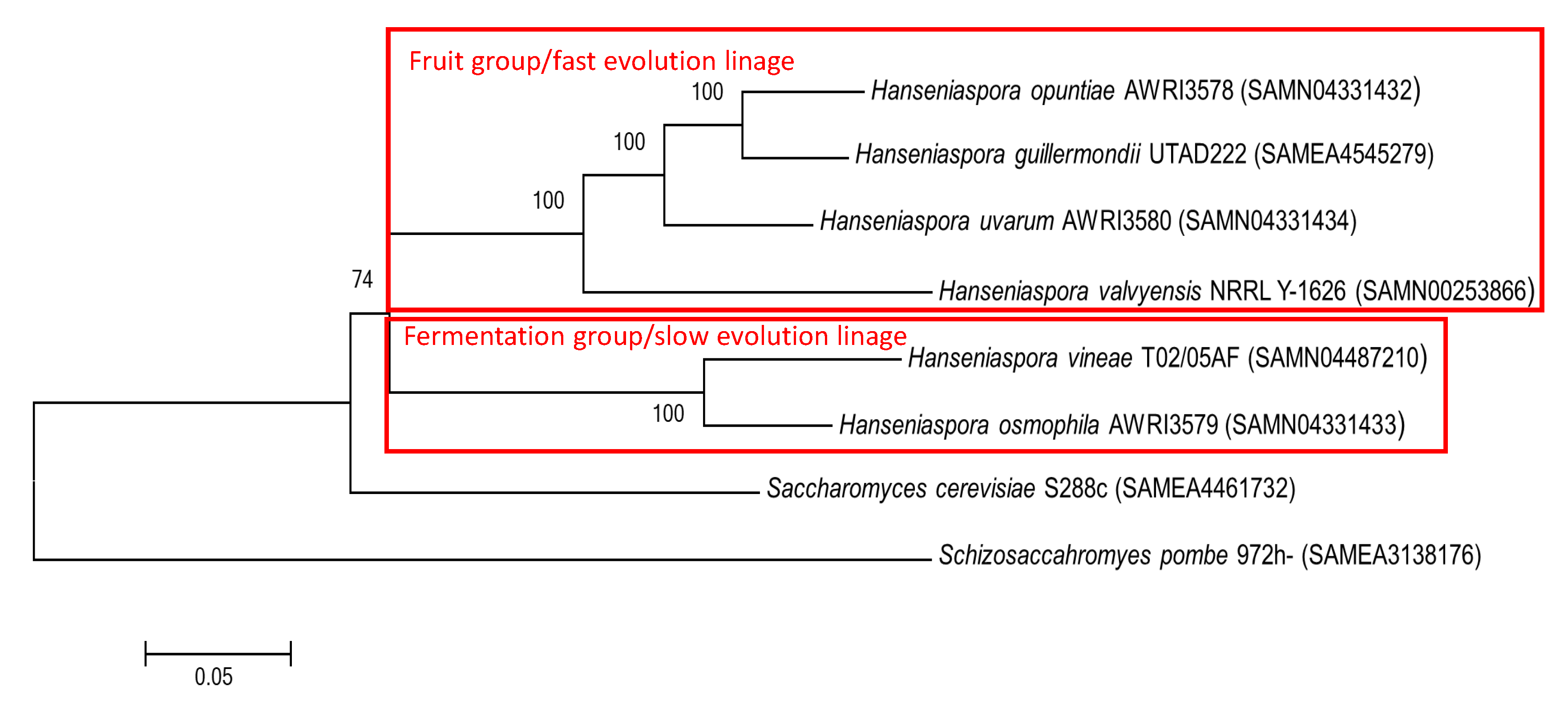
3.5. Hanseniaspora Genus as an Evolution Model for Alcoholic Fermentation Adaptations
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rainieri, S.; Pretorius, I.S. Selection and improvement of wine yeasts. Ann. Microbiol. 2000, 50, 15–31. [Google Scholar]
- Fleet, G.H. Wine yeasts for the future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas, A.; Guillamón, J.M.; Beltran, G. Non-conventional Yeast in the Wine Industry. FEMS Yeast Res. 2016, 7, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Carrau, F.; Gaggero, C.; Aguilar, P.S. Yeast diversity and native vigor for flavor phenotypes. Trends Biotechnol. 2015, 33, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Lleixà, J.; Manzano, M.; Mas, A.; Portillo, M.d.C. Saccharomyces and non-Saccharomyces competition during microvinification under different sugar and nitrogen conditions. Front. Microbiol. 2016, 7, 1959. [Google Scholar] [CrossRef]
- Masneuf-Pomarede, I.; Bely, M.; Marullo, P.; Albertin, W. The genetics of non-conventional wine yeasts: Current knowledge and future challenges. Front. Microbiol. 2016, 6, 1563. [Google Scholar] [CrossRef]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Gostíncar, A.; Bobet, R.; Uruburu, F.; Querol, A. Selection and molecular characterization of wine yeasts isolated from the “El Penedes” area (Spain). Food Microbiol. 2000, 17, 553–562. [Google Scholar] [CrossRef]
- Degre, R. Selection and Commercial Cultivation of Wine Yeast and Bacteria. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Taylor and Francis: New York, NY, USA, 1993. [Google Scholar]
- Romano, P.; Capece, A.; Jespersen, L. Taxonomic and ecological diversity of food and beverage yeasts. In Yeasts in Foods and Beverages; Springer: Berlin/Heidelberg, Germany, 2006; pp. 13–53. [Google Scholar]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Effect of Non-Saccharomyces Yeasts on Fermentation and Wine Quality. S. Afr. J. Enol. Vitic. 2003, 24, 55–62. [Google Scholar] [CrossRef]
- Zott, K.; Miot-Sertier, C.; Claisse, O.; Lonvaud-Funel, A.; Masneuf-Pomarede, I. Dynamics and diversity of non-Saccharomyces yeasts during the early stages in winemaking. Int. J. Food Microbiol. 2008, 125, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Pina, C.; Santos, C.; Couto, J.A.; Hogg, T. Ethanol tolerance of five non-Saccharomyces wine yeasts in comparison with a strain of Saccharomyces cerevisiae—Influence of different culture conditions. Food Microbiol. 2004, 21, 439–447. [Google Scholar] [CrossRef]
- Albergaria, H.; Francisco, D.; Gori, K.; Arneborg, N.; Gírio, F. Saccharomyces cerevisiae CCMI 885 secretes peptides that inhibit the growth of some non-Saccharomyces wine-related strains. Appl. Microbiol. Biotechnol. 2010, 86, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Dang, W.; Xie, J.; Zhu, R.; Sun, M.; Jia, F.; Zhao, Y.; An, X.; Qiu, S.; Li, X.; et al. Antimicrobial peptide protonectin disturbs the membrane integrity and induces ROS production in yeast cells. Biochim. Biophys. Acta Biomembr. 2015, 1848, 2365–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, N.; Pina, C.; Mendes, F.; Couto, J.A.; Hogg, T.; Vasconcelos, I. Volatile compounds contribution of Hanseniaspora guilliermondii and Hanseniaspora uvarum during red wine vinifications. Food Control 2011, 22, 662–667. [Google Scholar] [CrossRef]
- Martin, V.; Valera, M.J.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Giorello, F.; Valera, M.J.; Martin, V.; Parada, A.; Salzman, V.; Camesasca, L.; Fariña, L.; Boido, E.; Medina, K.; Dellacassa, E.; et al. Genomic and transcriptomic basis of Hanseniaspora vineae’s impact on flavor diversity and wine quality. Appl. Environ. Microbiol. 2019, 75, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Valera, M.J.; Boido, E.; Ramos, J.C.; Manta, E.; Radi, R.; Dellacassa, E.; Carrau, F. The mandelate pathway, an alternative to the PAL pathway for the synthesis of benzenoids in yeast. Appl. Environ. Microbiol. 2020, 86, 701–720. [Google Scholar] [CrossRef] [PubMed]
- Giorello, F.M.; Berna, L.; Greif, G.; Camesasca, L.; Salzman, V.; Medina, K.; Robello, C.; Gaggero, C.; Aguilar, P.S.; Carrau, F. Genome Sequence of the Native Apiculate Wine Yeast Hanseniaspora vineae T02/19AF. Genome Announc. 2014, 2, e00530-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternes, P.R.; Lee, D.; Kutyna, D.R.; Borneman, A.R. Genome Sequences of Three Species of Hanseniaspora Isolated from Spontaneous Wine Fermentations. Genome Announc. 2016, 4, e01287-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, R.; Haridas, S.; Wolfe, K.H.; Lopes, M.R.; Hittinger, C.T.; Göker, M.; Salamov, A.A.; Wisecaver, J.H.; Long, T.M.; Calvey, C.H.; et al. Comparative genomics of biotechnologically important yeasts. Proc. Natl. Acad. Sci. USA 2016, 113, 9882–9887. [Google Scholar] [CrossRef] [Green Version]
- Seixas, I.; Barbosa, C.; Mendes-Faia, A.; Güldener, U.; Tenreiro, R.; Mendes-Ferreira, A.; Mira, N.P. Genome sequence of the non-conventional wine yeast Hanseniaspora guilliermondii UTAD222 unveils relevant traits of this species and of the Hanseniaspora genus in the context of wine fermentation. DNA Res. 2019, 26, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.E.; Nguyen, T.D.; Liccioli, T.; Schmid, F.; Kalatzis, N.; Sundstrom, J.F.; Gardner, J.M.; Jiranek, V. Genome-wide identification of the Fermentome; genes required for successful and timely completion of wine-like fermentation by Saccharomyces cerevisiae. BMC Genom. 2014, 15, 552. [Google Scholar] [CrossRef] [Green Version]
- Langenberg, A.; Bink, F.J.; Wolff, L.; Walter, S.; Grossmann, M.; Schmitz, H. Glycolytic Functions Are Conserved in the Genome of the Wine Yeast. Appl. Environ. Microbiol. 2017, 83, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Stanke, M.; Keller, O.; Gunduz, I.; Hayes, A.; Waack, S.; Morgenstern, B. AUGUSTUS: Ab initio prediction of alternative transcripts. Nucleic Acids Res. 2006, 34, W435–W439. [Google Scholar] [CrossRef] [Green Version]
- Cherry, J.M.; Hong, E.L.; Amundsen, C.; Balakrishnan, R.; Binkley, G.; Chan, E.T.; Christie, K.R.; Costanzo, M.C.; Dwight, S.S.; Engel, S.R.; et al. Saccharomyces Genome Database: The genomics resource of budding yeast. Nucleic Acids Res. 2012, 40, D700–D706. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Carrau, F.M.; Medina, K.; Boido, E.; Farina, L.; Gaggero, C.; Dellacassa, E.; Versini, G.; Henschke, P.A. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 2005, 243, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henschke, P.A.; Jiranek, V. Yeast: Metabolism of nitrogen compounds. In Wine Microbiology and Biotechnology; Taylor and Francis: New York, NY, USA, 1993; ISBN 0-415-27850-3. [Google Scholar]
- Steenwyk, J.L.; Opulente, D.A.; Kominek, J.; Shen, X.X.; Zhou, X.; Labella, A.L.; Bradley, N.P.; Eichman, B.F.; Čadež, N.; Libkind, D.; et al. Extensive loss of cell-cycle and DNA repair genes in an ancient lineage of bipolar budding yeasts. PLoS Biol. 2019, 17, e3000255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410.e16. [Google Scholar] [CrossRef] [Green Version]
- Albertin, W.; Setati, M.E.; Miot-Sertier, C.; Mostert, T.T.; Colonna-Ceccaldi, B.; Coulon, J.; Girard, P.; Moine, V.; Pillet, M.; Salin, F.; et al. Hanseniaspora uvarum from winemaking environments show spatial and temporal genetic clustering. Front. Microbiol. 2016, 6, 1569. [Google Scholar] [CrossRef]
- Forsberg, H.; Ljungdahl, P.O. Sensors of extracellular nutrients in Saccharomyces cerevisiae. Curr. Genet. 2001, 40, 91–109. [Google Scholar] [CrossRef]
- Jordan, P.; Choe, J.Y.; Boles, E.; Oreb, M. Hxt13, Hxt15, Hxt16 and Hxt17 from Saccharomyces cerevisiae represent a novel type of polyol transporters. Sci. Rep. 2016, 6, 23502. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, A.; De La Cera, T.; Herrero, P.; Moreno, F. The hexokinase 2 protein regulates the expression of the GLK1, HXK1 and HXK2 genes of Saccharomyces cerevisiae. Biochem. J. 2001, 355, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Reifenberger, E.; Freidel, K.; Ciriacy, M. Identification of novel HXT genes in Saccharomyces cerevisiae reveals the impact of individual hexose transporters on qlycolytic flux. Mol. Microbiol. 1995, 16, 157–167. [Google Scholar] [CrossRef]
- Tondini, F.; Lang, T.; Chen, L.; Herderich, M.; Jiranek, V. Linking gene expression and oenological traits: Comparison between Torulaspora delbrueckii and Saccharomyces cerevisiae strains. Int. J. Food Microbiol. 2019, 294, 42–49. [Google Scholar] [CrossRef]
- Ozcan, S.; Johnston, M. Function and regulation of yeast hexose transporters. Microbiol. Mol. Biol. Rev. 1999, 63, 554–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, C.; Van Voorst, F.; Martins, A.; Neves, L.; Oliveira, R.; Kielland-Brandt, M.C.; Lucas, C.; Brandt, A. A member of the sugar transporter family, Stl1p is the glycerol/H + symporter in Saccharomyces cerevisiae. Mol. Biol. Cell 2005, 16, 2068–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casal, M.; Paiva, S.; Andrade, R.P.; Gancedo, C.; Leão, C. The lactate-proton symport of Saccharomyces cerevisiae is encoded by JEN1. J. Bacteriol. 1999, 181, 2620–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akita, O.; Nishimori, C.; Shimamoto, T.; Fujii, T.; Iefuji, H. Transport of pyruvate in Saccharomyces cerevisiae and cloning of the gene encoded pyruvate permease. Biosci. Biotechnol. Biochem. 2000, 64, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Cortassa, S.; Aon, J.C.; Aon, M.A. Fluxes of carbon, phosphorylation, and redox intermediates during growth of Saccharomyces cerevisiae on different carbon sources. Biotechnol. Bioeng. 1995, 47, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, A.; Zimmermann, F.K. Isolation and molecular analysis of the phosphoglucose isomerase structural gene of Saccharomyces cerevisiae. MGG Mol. Gen. Genet. 1986, 202, 83–89. [Google Scholar] [CrossRef]
- Aguilera, A. Deletion of the phosphoglucose isomerase structural gene makes growth and sporulation glucose dependent in Saccharomyces cerevisiae. MGG Mol. Gen. Genet. 1986, 204, 310–316. [Google Scholar] [CrossRef]
- McAlister, L.; Holland, M.J. Differential expression of the three yeast glyceraldehyde-3-phosphate dehydrogenase genes. J. Biol. Chem. 1985, 260, 15019–15027. [Google Scholar]
- McAlister, L.; Holland, M.J. Isolation and characterization of yeast strains carrying mutations in the glyceraldehyde-3-phosphate dehydrogenase genes. J. Biol. Chem. 1985, 260, 15013–15018. [Google Scholar]
- Grauslund, M.; Rønnow, B. Carbon source-dependent transcriptional regulation of the mitochondrial glycerol-3-phosphate dehydrogenase gene, GUT2, from Saccharomyces cerevisiae. Can. J. Microbiol. 2000, 46, 1096–1100. [Google Scholar] [CrossRef]
- Boles, E.; Hollenberg, C.P. The molecular genetics of hexose transport in yeasts. FEMS Microbiol. Rev. 1997, 21, 85–111. [Google Scholar] [CrossRef] [PubMed]
- Jurica, M.S.; Mesecar, A.; Heath, P.J.; Shi, W.; Nowak, T.; Stoddard, B.L. The allosteric regulation of pyruvate kinase by fructose-1,6-bisphosphate. Structure 1998, 6, 195–210. [Google Scholar] [CrossRef] [Green Version]
- Kellermann, E.; Seeboth, P.G.; Hollenberg, C.P. Analysis of the primary structure and promoter function of a pyruvate decarboxylase gene (PDCI) from Saccharomyces cerevisiae. Nucleic Acids Res. 1986, 14, 8963–8977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, S.; Cederberg, H. Autoregulation may control the expression of yeast pyruvate decarboxylase structural genes PDC1 and PDC5. Eur. J. Biochem. 1990, 188, 615–621. [Google Scholar] [CrossRef]
- König, S. Subunit structure, function and organisation of pyruvate decarboxylases from various organisms. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1998, 1385, 271–286. [Google Scholar] [CrossRef]
- Muller, E.H.; Richards, E.J.; Norbeck, J.; Byrne, K.L.; Karlsson, K.A.; Pretorius, G.H.J.; Meacock, P.A.; Blomberg, A.; Hohmann, S. Thiamine repression and pyruvate decarboxylase autoregulation independently control the expression of the Saccharomyces cerevisiae PDC5 gene. FEBS Lett. 1999, 449, 248–250. [Google Scholar] [CrossRef] [Green Version]
- Eberhardt, I.; Cederberg, H.; Li, H.; König, S.; Jordan, F.; Hohmann, S. Autoregulation of yeast pyruvate decarboxylase gene expression requires the enzyme but not its catalytic activity. Eur. J. Biochem. 1999, 262, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Bataillon, M.; Rico, A.; Sablayrolles, J.M.; Salmon, J.M.; Barre, P. Early thiamin assimilation by yeasts under enological conditions: Impact on alcoholic fermentation kinetics. J. Ferment. Bioeng. 1996, 82, 145–150. [Google Scholar] [CrossRef]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef]
- Rossouw, D.; Bauer, F.F. Comparing the transcriptomes of wine yeast strains: Toward understanding the interaction between environment and transcriptome during fermentation. Appl. Microbiol. Biotechnol. 2009, 84, 937. [Google Scholar] [CrossRef]
- Gonçalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M.; Libkind, D.; Hutzler, M.; Gonçalves, P.; Sampaio, J.P. Distinct Domestication Trajectories in Top-Fermenting Beer Yeasts and Wine Yeasts. Curr. Biol. 2016, 26, 2750–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Source | Use |
|---|---|---|---|
| H. vineae | T02/19AF | Fermenting Tannat grape must (Uruguay) | Genomic, transcriptomic, phenotypic analysis |
| H. vineae | T02/05AF | Fermenting Tannat grape must (Uruguay) | Genomic and phenotypic analysis |
| H. osmophila | AWRI3579 | Fermenting Chardonnay grape must (Australia) | Genomic and phenotypic analysis |
| H. uvarum | AWRI1280 | Fermenting Tannat grape must (Uruguay) | Phenotypic analysis |
| H. uvarum | AWRI3580 | Fermenting Chardonnay grape (Australia) | Genomic analysis |
| H. opuntiae | AWRI3578 | Fermenting Chardonnay grape (Australia) | Genomic analysis |
| H. valbyensis | NRRL Y-1626 | Soil (Denmark) | Genomic analysis |
| H. guilliermondii | UTAD222 | Grape must (Portugal) | Genomic analysis |
| S. cerevisiae | 288Sc | Laboratory strain | Genomic analysis |
| S. cerevisiae | ALG804 | Oenological yeast (Oenobrands®) | Phenotypic analysis |
| S. cerevisiae | H. vineae | |
|---|---|---|
| Sugar transport and sensors | HXT (x17); SNF3; RGT2; FPS1; GPR1; GUP1; GUP2; STL1; JEN1; ASC1; ASC2; GPA2 | HXT (x2); SNF3; GPR1; GUP1; STL1 (x2); JEN1; ASC1; GPA2 |
| Glycolysis | HXK1; HXK2; PGI1; PFK1; PFK2; FPBA1; TPI1; TDH1; TDH2; TDH3; PGK1; GPM1; ENO1; ENO2; CDC19; PYK2 | HXK2; PGI1; PFK1; PFK2; FPBA1; TPI1; TDH2; TDH3; PGK1; GPM1; ENO1; ENO2; CDC19 |
| Alcoholic fermentation | PDC1; PDC2; PDC5, PDC6; ADH (x8) | PDC1; ADH (x8) |
| Key genes of wine yeasts adaptations | SSU1; CUP1 (x2); SUC2; THI5; THI11, THI12, THI13; THI14; THI16; THI20; THI21; THI72; THI73; THI80; TPC1 | SSU1; SUC2 THI7; THI72; THI80; TPC1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valera, M.J.; Boido, E.; Dellacassa, E.; Carrau, F. Comparison of the Glycolytic and Alcoholic Fermentation Pathways of Hanseniaspora vineae with Saccharomyces cerevisiae Wine Yeasts. Fermentation 2020, 6, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030078
Valera MJ, Boido E, Dellacassa E, Carrau F. Comparison of the Glycolytic and Alcoholic Fermentation Pathways of Hanseniaspora vineae with Saccharomyces cerevisiae Wine Yeasts. Fermentation. 2020; 6(3):78. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030078
Chicago/Turabian StyleValera, María José, Eduardo Boido, Eduardo Dellacassa, and Francisco Carrau. 2020. "Comparison of the Glycolytic and Alcoholic Fermentation Pathways of Hanseniaspora vineae with Saccharomyces cerevisiae Wine Yeasts" Fermentation 6, no. 3: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030078