β-Glucosidase Activity of Lactiplantibacillus plantarum UNQLp 11 in Different Malolactic Fermentations Conditions: Effect of pH and Ethanol Content

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
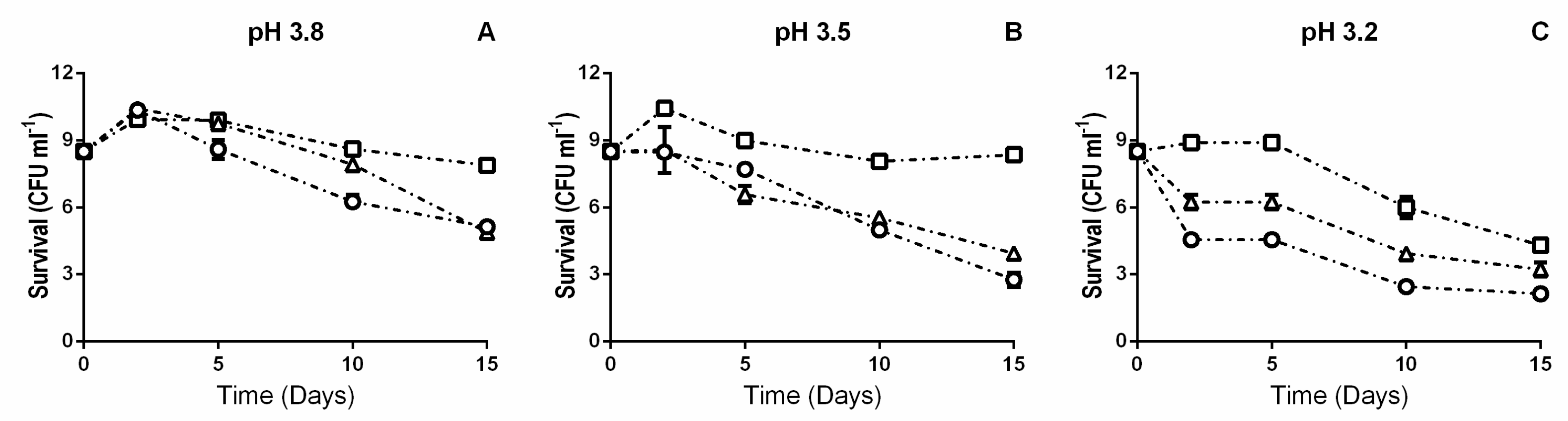
2.1. Growth, Acclimation, and Wine Inoculation
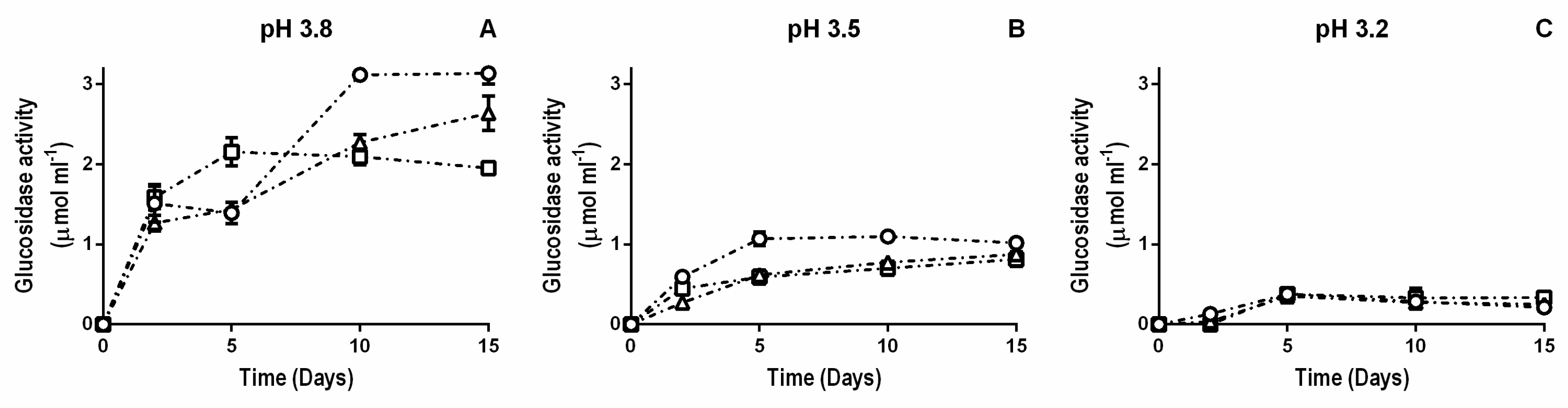
2.2. β-Glucosidase Activity during Fermentation in Synthetic Wine
2.3. Hydrolysis of octyl β-D-glucopyranoside in Sterile Pinot Noir Wine
2.4. Statistical Analysis
3. Results
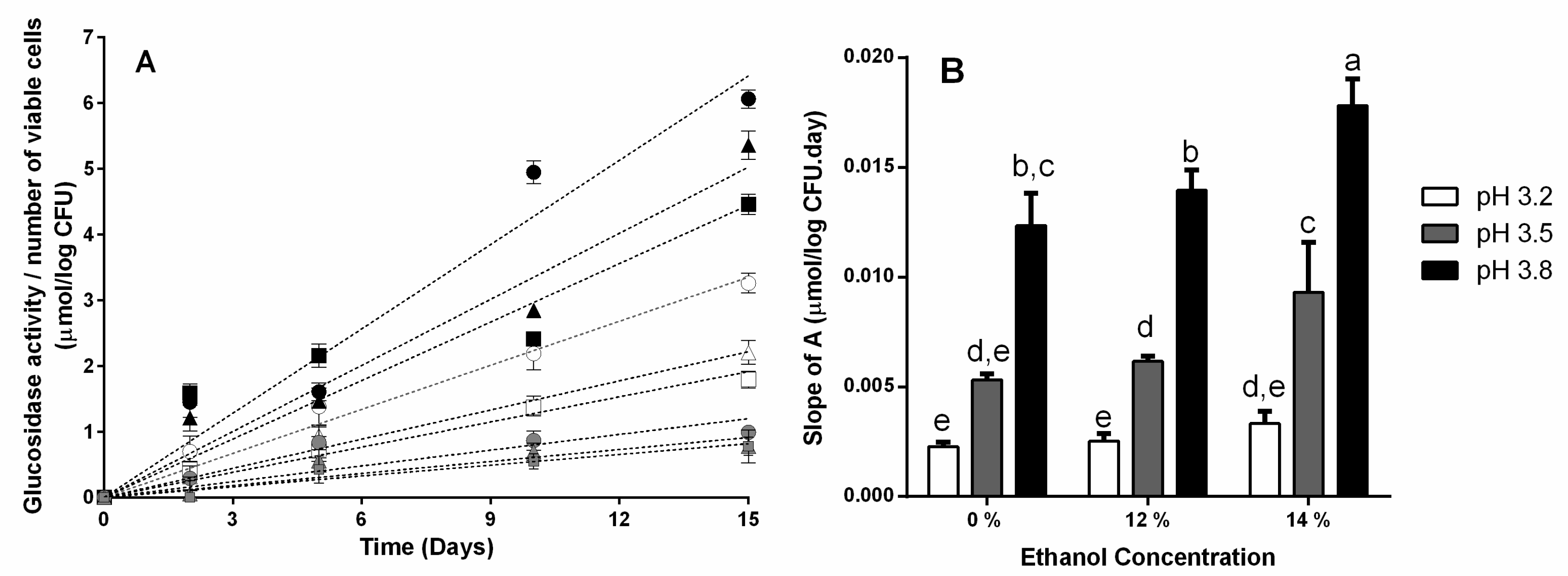
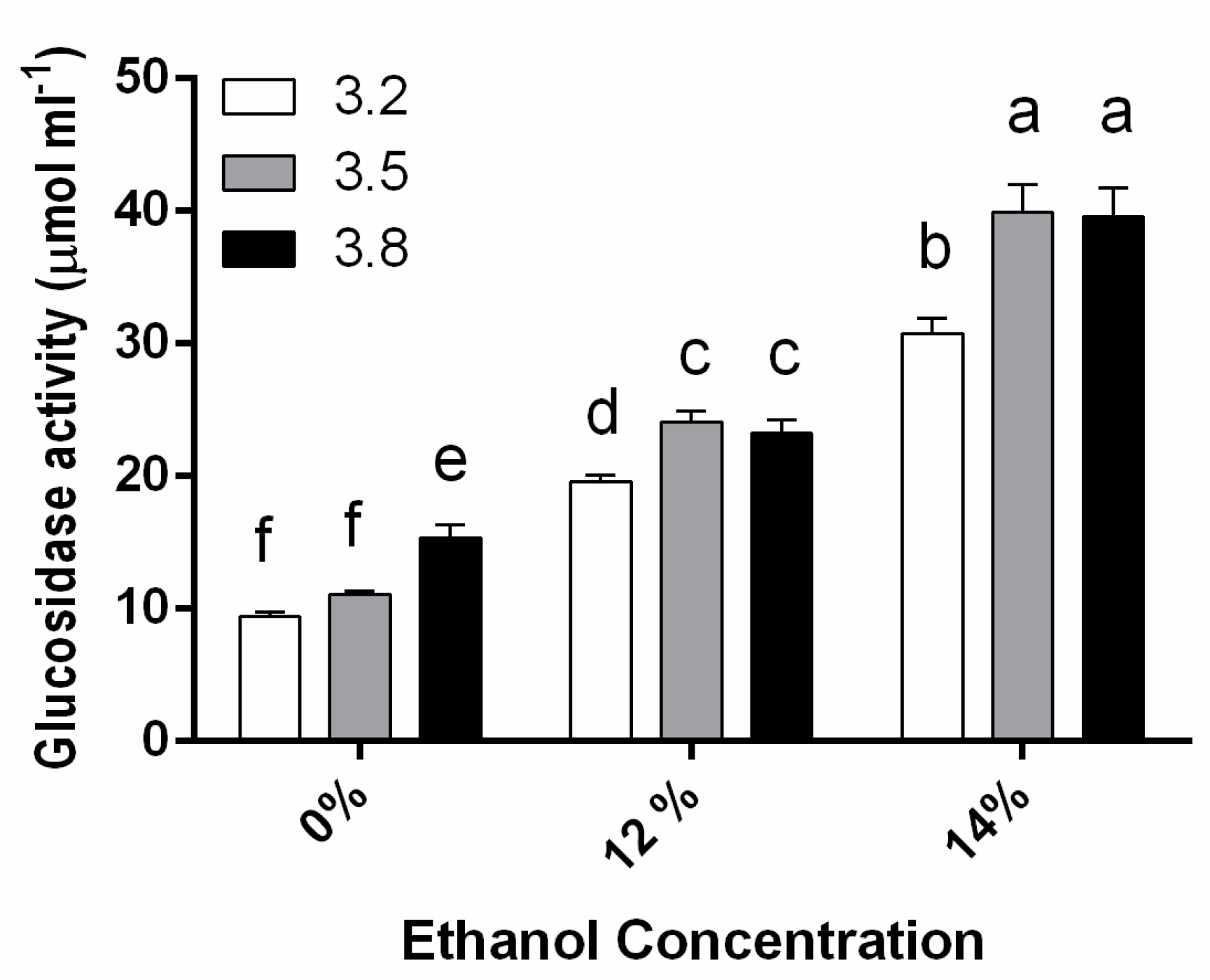
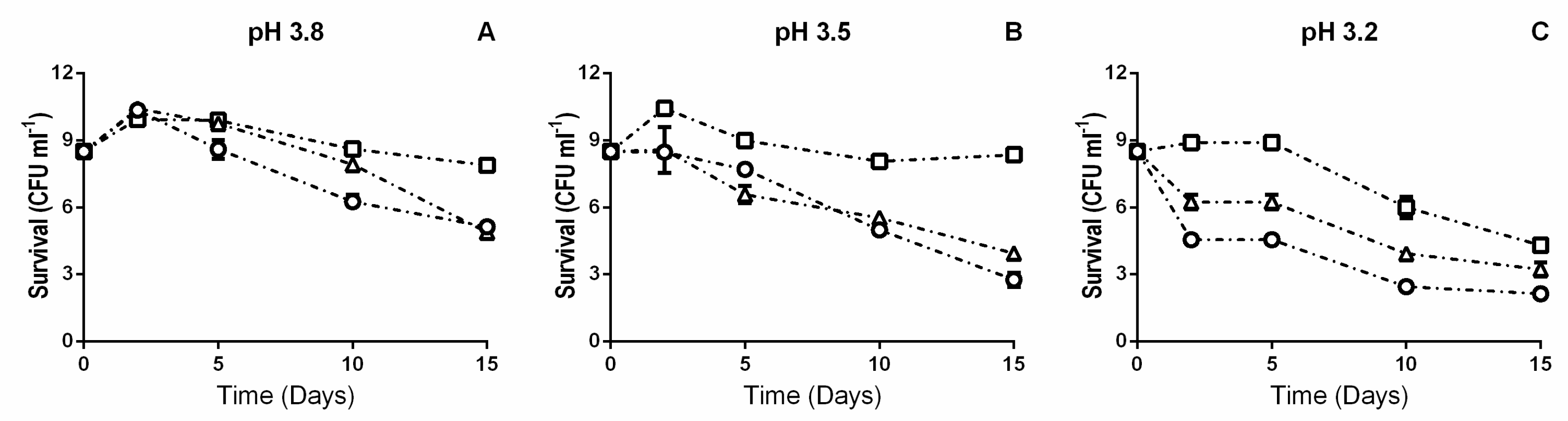
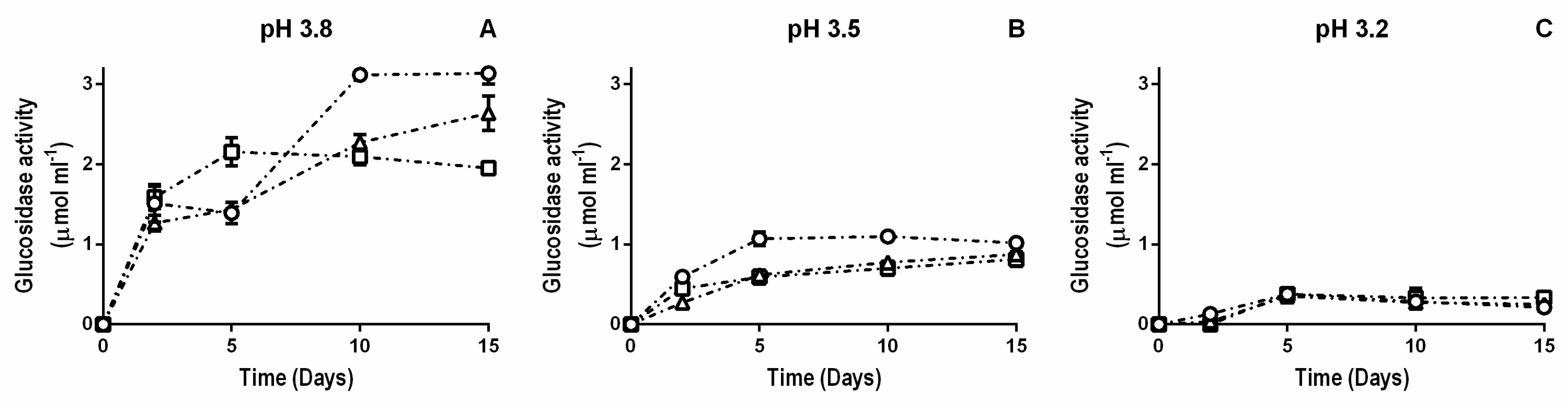
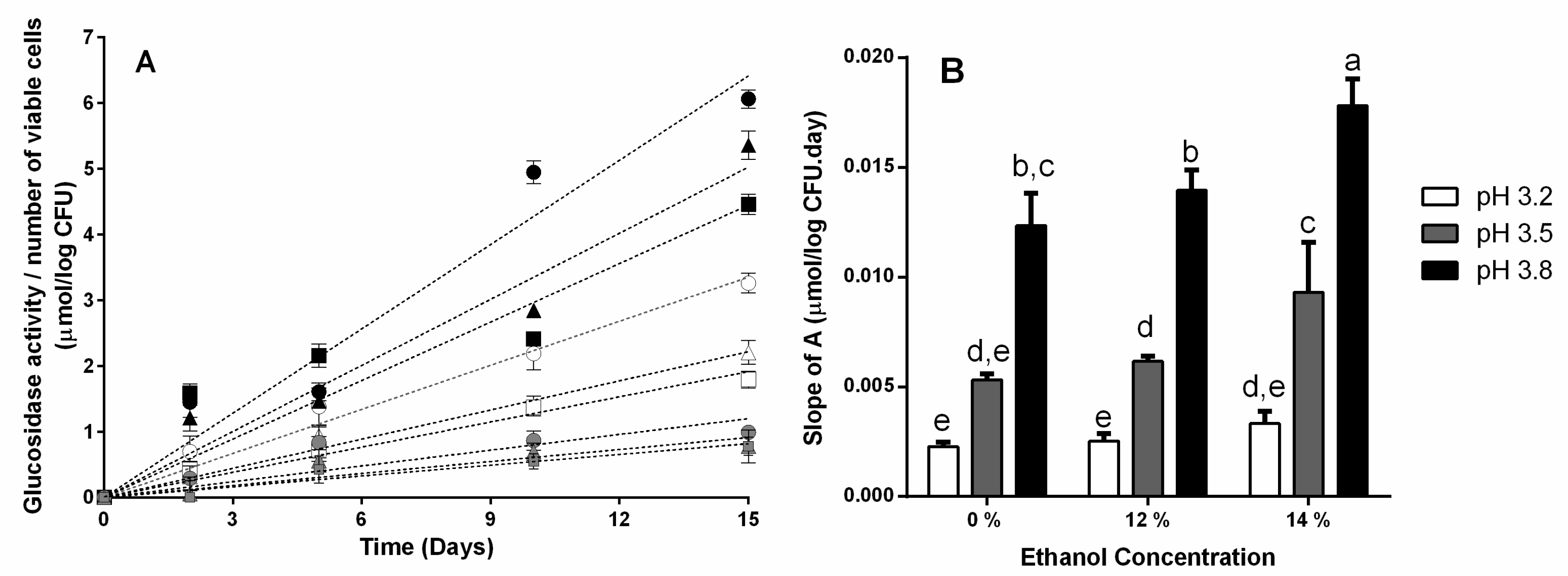
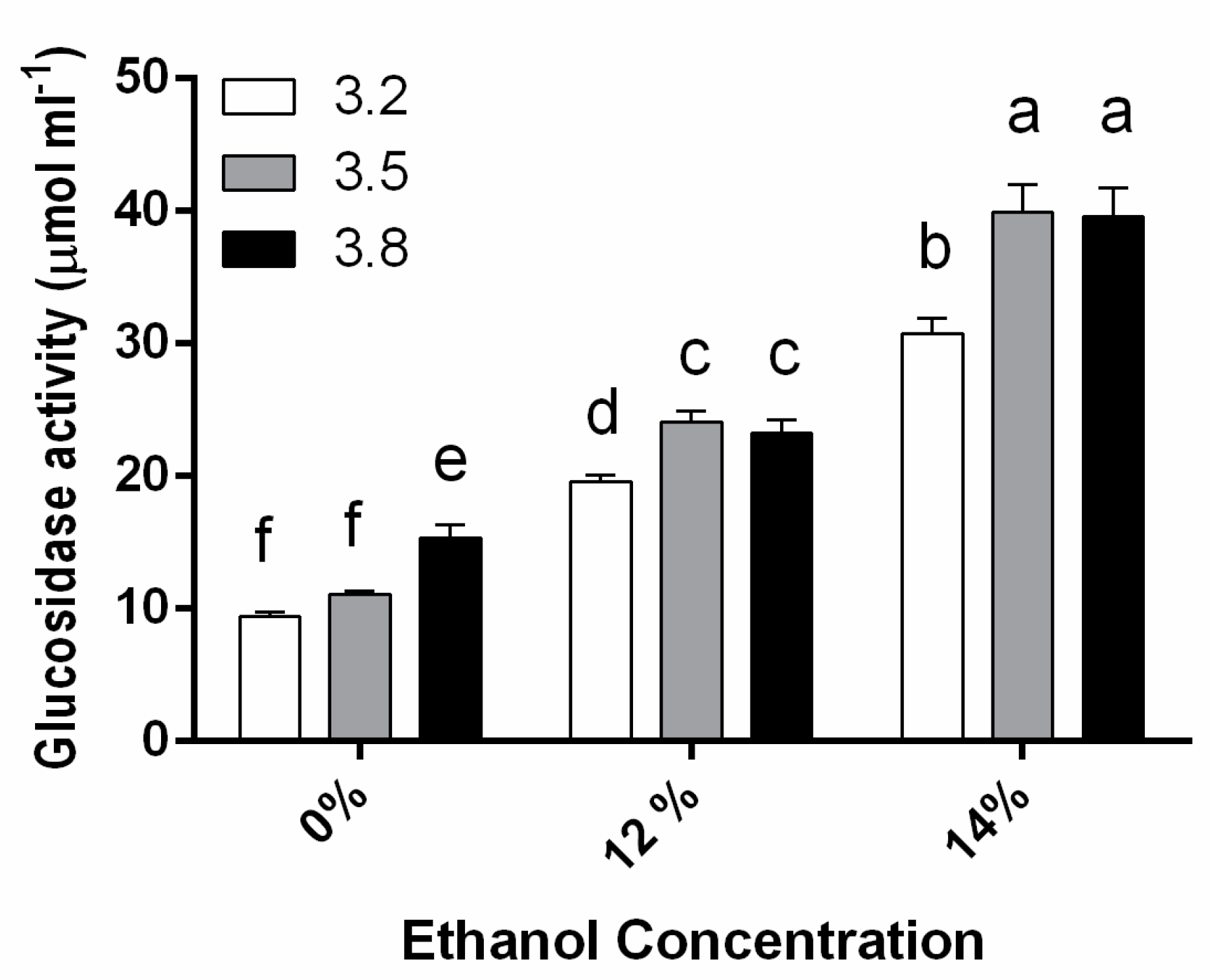
3.1. β-Glucosidase Activity during Fermentation in Synthetic Wine
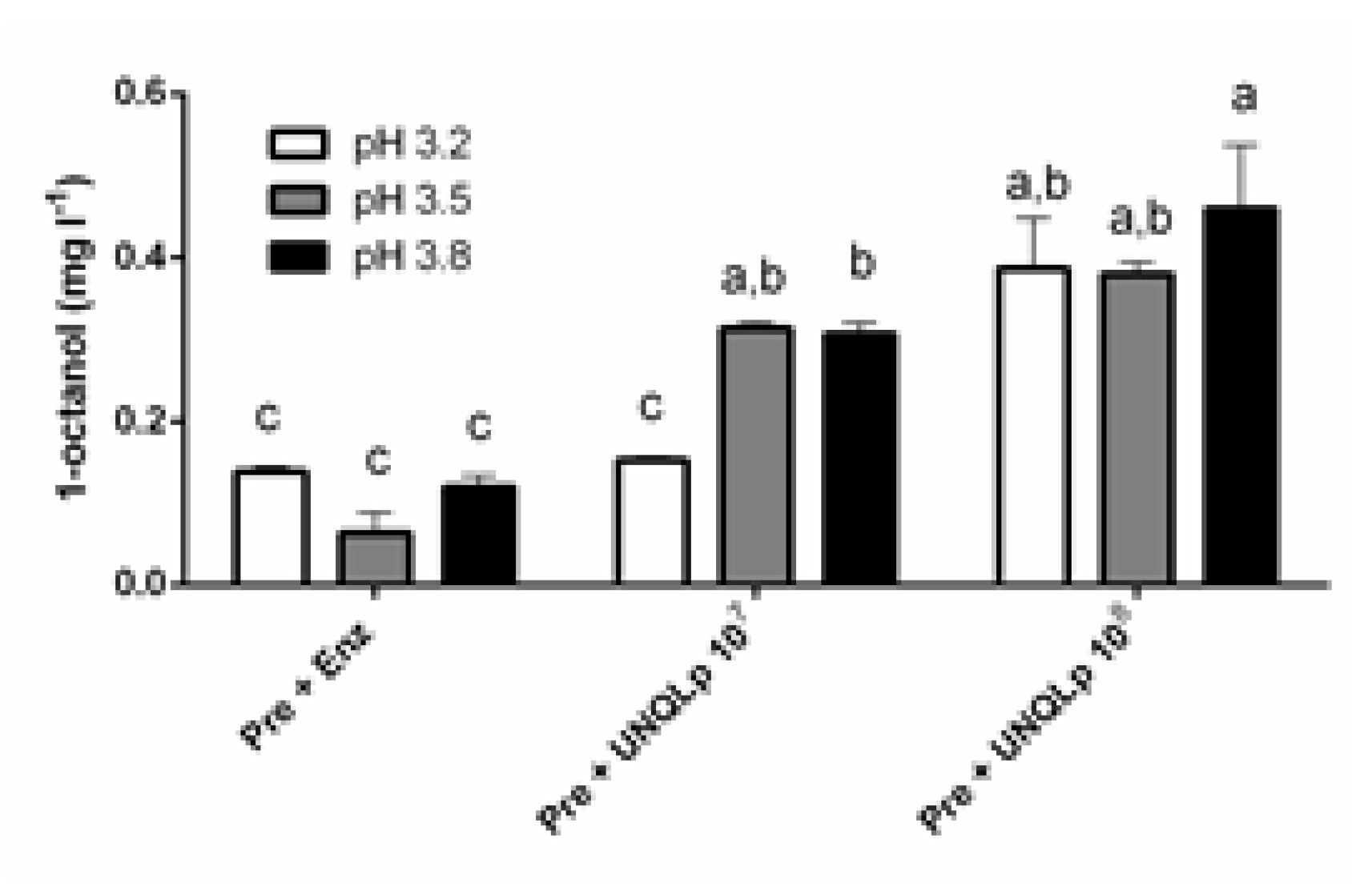
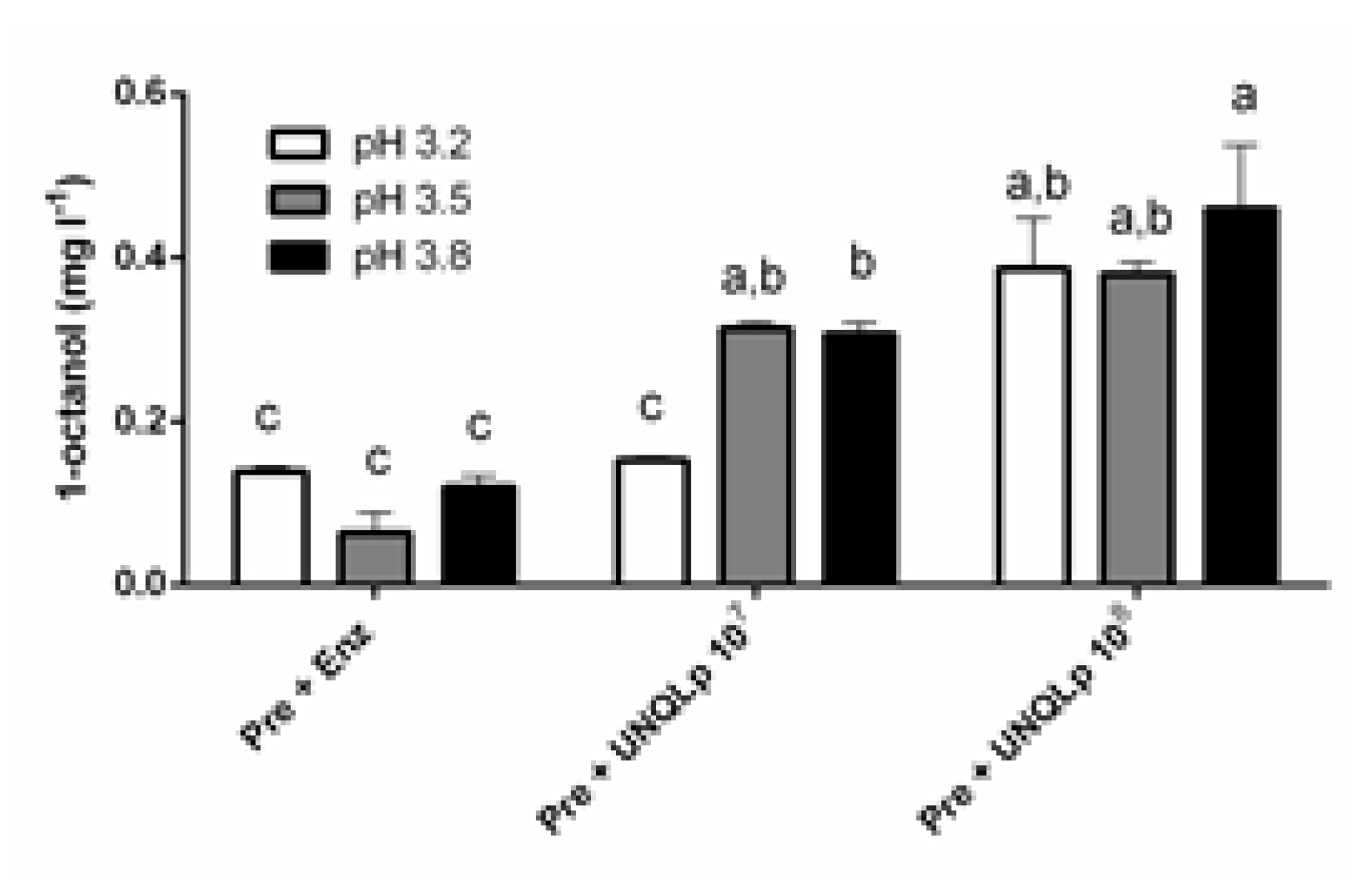
3.2. Hydrolysis of Octyl β-D-Glucopyranoside in sterile Pinot Noir Wine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Lerm, E.; Engelbrecht, L.; Du Toit, M. Malolactic fermentation: The ABC’s of MLF. S. Afr. J. Enol. Vitic. 2010, 31, 186–212. [Google Scholar] [CrossRef] [Green Version]
- du Toit, M.; Engelbrecht, L.; Lerm, E.; Krieger-Weber, S. Lactobacillus: The next generation of malolactic fermentation starter cultures—An overview. Food Bioprocess Technol. 2011, 4, 876–906. [Google Scholar] [CrossRef]
- Gunata, Y.Z.; Bayonove, C.L.; Baumes, R.L.; Cordonnier, R.E. The aroma of grapes. Localisation and evolution of free and bound fractions of some grape aroma components cv Muscat during first development and maturation. J. Sci. Food Agric. 1985, 36, 857–862. [Google Scholar] [CrossRef]
- Grimaldi, A.; Bartowsky, E.; Jiranek, V. A survey of glycosidase activities of commercial wine strains of Oenococcus oeni. Int. J. Food Microbiol. 2005, 105, 233–244. [Google Scholar] [CrossRef]
- Mateo, J.J.; Maicas, S. Application of non-Saccharomyces yeasts to wine-making process. Fermentation 2016, 2, 14. [Google Scholar] [CrossRef]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; García-Ruiz, A.; Muñoz-González, C.; Bartolomé, B.; Moreno-Arribas, M.V. Selection and technological potential of Lactobacillus plantarum bacteria suitable for wine malolactic fermentation and grape aroma release. LWT 2016, 73, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Le Traon-Masson, M.P.; Pellerin, P. Purification and characterization of two β-d-glucosidases from an Aspergillus niger enzyme preparation: Affinity and specificity toward glucosylated compounds characteristic of the processing of fruits. Enzym. Microb. Technol. 1998, 22, 374–382. [Google Scholar] [CrossRef]
- Codresi, C.; Râpeanu, G.; Alexe, P. Effect of β-glucosidases in the making of chardonnay wines. Ann. Univ. Dun Area De Jos Galati.Fascicle Vi. Food Technol. 2012, 36, 9. [Google Scholar]
- Spagna, G.; Barbagallo, R.N.; Greco, E.; Manenti, I.; Pifferi, P.G. A mixture of purified glycosidases from Aspergillus niger for oenological application immobilised by inclusion in chitosan gels. Enzym. Microb. Technol. 2002, 30, 80–89. [Google Scholar] [CrossRef]
- Grimaldi, A.; McLean, H.; Jiranek, V. Identification and partial characterization of glycosidic activities of commercial strains of the lactic acid bacterium, Oenococcus oeni. Am. J. Enol. Vitic. 2000, 51, 362–369. [Google Scholar]
- Grimaldi, A.; Bartowsky, E.; Jiranek, V. Screening of Lactobacillus spp. and Pediococcus spp. for glycosidase activities that are important in oenology. J. Appl. Microbiol. 2005, 99, 1061–1069. [Google Scholar] [CrossRef]
- Bravo-Ferrada, B.M.; Hollmann, A.; Delfederico, L.; La Hens, D.V.; Caballero, A.; Semorile, L. Patagonian red wines: Selection of Lactobacillus plantarum isolates as potential starter cultures for malolactic fermentation. World J. Microbiol. Biotechnol. 2013, 29, 1537–1549. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; La Hens, D.V.; Hollmann, A.; Delfederico, L.; Caballero, A.; Tymczyszyn, E.E.; Semorile, L. Comparative vinification assays with selected Patagonian strains of Oenococcus oeni and Lactobacillus plantarum. LWT 2017, 77, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Barbagallo, R.N.; Spagna, G.; Palmeri, R.; Restuccia, C.; Giudici, P. Selection, characterization and comparison of β-glucosidase from mould and yeasts employable for enological applications. Enzym. Microb. Technol. 2004, 35, 58–66. [Google Scholar] [CrossRef]
- Olguín, N.; Alegret, J.O.; Bordons, A.; Reguant, C. β-Glucosidase activity and bgl gene expression of Oenococcus oeni strains in model media and Cabernet Sauvignon wine. Am. J. Enol. Vitic. 2011, 62, 99–105. [Google Scholar] [CrossRef]
- Pérez-Martín, F.; Seseña, S.; Izquierdo, P.M.; Palop, M.L. Esterase activity of lactic acid bacteria isolated from malolactic fermentation of red wines. Int. J. Food Microbiol. 2013, 163, 153–158. [Google Scholar] [CrossRef] [PubMed]
- De Orduna, R.M. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Kontoudakis, N.; Esteruelas, M.; Fort, F.; Canals, J.M.; Zamora, F. Use of unripe grapes harvested during cluster thinning as a method for reducing alcohol content and pH of wine. Aust. J. Grape Wine Res. 2011, 17, 230–238. [Google Scholar] [CrossRef]
- Gockowiak, H.; Henschke, P.A. Interaction of pH, ethanol concentration and wine matrix on induction of malolactic fermentation with commercial ‘direct inoculation’starter cultures. Aust. J. Grape Wine Res. 2003, 9, 200–209. [Google Scholar] [CrossRef]
- Cinquanta, L.; De Stefano, G.; Formato, D.; Niro, S.; Panfili, G. Effect of pH on malolactic fermentation in southern Italian wines. Eur. Food Res. Technol. 2018, 244, 1261–1268. [Google Scholar] [CrossRef]
- Davis, C.R.; Wibowo, D.J.; Lee, T.H.; Fleet, G.H. Growth and metabolism of lactic acid bacteria during and after malolactic fermentation of wines at different pH. Appl. Environ. Microbiol. 1986, 51, 539–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brizuela, N.; Tymczyszyn, E.E.; Semorile, L.C.; La Hens, D.V.; Delfederico, L.; Hollmann, A.; Bravo-Ferrada, B. Lactobacillus plantarum as a malolactic starter culture in winemaking: A new (old) player? Electron. J. Biotechnol. 2019, 38, 10–18. [Google Scholar] [CrossRef]
- Lerm, E.; Engelbrecht, L.; Du Toit, M. Selection and characterisation of Oenococcus oeni and Lactobacillus plantarum South African wine isolates for use as malolactic fermentation starter cultures. South Afr. J. Enol. Vitic. 2011, 32, 280–295. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Russo, P.; Beneduce, L.; Weidmann, S.; Grieco, F.; Guzzo, J.; Spano, G. Technological properties of Oenococcus oeni strains isolated from typical southern Italian wines. Lett. Appl. Microbiol. 2010, 50, 327–334. [Google Scholar] [CrossRef]
- Ruiz, P.; Izquierdo, P.M.; Seseña, S.; Palop, M.L. Selection of autochthonous Oenococcus oeni strains according to their oenological properties and vinification results. Int. J. Food Microbiol. 2010, 137, 230–235. [Google Scholar] [CrossRef]
- Valdes la Hens, D.; Bravo-Ferrada, B.M.; Delfederico, L.; Caballero, A.C.; Semorile, L.C. Prevalence of Lactobacillus plantarum and Oenococcus oeni during spontaneous malolactic fermentation in Patagonian red wines revealed by polymerase chain reaction-denaturing gradient gel electrophoresis with two targeted genes. Aust. J. Grape Wine Res. 2015, 21, 49–56. [Google Scholar] [CrossRef]
- Cerdeira, V.; Bravo-Ferrada, B.M.; Semorile, L.; Tymczyszyn, E. Design of a low-cost culture medium based in whey permeate for biomass production of enological Lactobacillus plantarum strains. Biotechnol. Prog. 2019, 35, e2791. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; Pozo-Bayón, M.Á.; Semorile, L.; Tymczyszyn, E.E. Changes in the volatile profile of Pinot noir wines caused by Patagonian Lactobacillus plantarum and Oenococcus oeni strains. Food Res. Int. 2018, 106, 22–28. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; Curilén, Y.; Delfederico, L.; Caballero, A.; Semorile, L.; Pozo-Bayón, M.; Tymczyszyn, E.E. Advantages of using blend cultures of native Lb. plantarum and O. oeni strains to induce malolactic fermentation of Patagonian Malbec wine. Front. Microbiol. 2018, 9, 2109. [Google Scholar] [CrossRef] [Green Version]
- Brizuela, N.S.; Franco-Luesma, E.; Bravo-Ferrada, B.M.; Pérez-Jiménez, M.; Semorile, L.; Tymczyszyn, E.; Pozo-Bayon, M.A. Influence of Patagonian Lactiplantibacillus plantarum and Oenococcus oeni strains on sensory perception after malolactic fermentation of Pinot Noir wine. Aust. J. Grape Wine Res. 2021, 27, 118–127. [Google Scholar] [CrossRef]
- Iglesias, N.G.; Brizuela, N.S.; Tymczyszyn, E.E.; Hollmann, A.; Valdés La Hens, D.; Semorile, L.; Bravo-Ferrada, B.M. Complete Genome Sequencing of Lactobacillus plantarum UNQLp 11 Isolated from a Patagonian Pinot Noir Wine. S. Afr. J. Enol. Vitic. 2020, 41, 197–209. [Google Scholar] [CrossRef]
- Bravo-Ferrada, B.M.; Tymczyszyn, E.E.; Gómez-Zavaglia, A.; Semorile, L. Effect of acclimation medium on cell viability, membrane integrity and ability to consume malic acid in synthetic wine by oenological Lactobacillus plantarum strains. J. Appl. Microbiol. 2014, 116, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Muñoz-González, C.; García-Ruiz, A.; Pozo-Bayón, M.; Lombardi, S.J.; Iorizzo, M.; Moreno-Arribas, M. Producción de compuestos aromáticos varietales por cepas de bacterias lácticas autóctonas a partir de un extracto de precursores glicosilados de uva blanca. Available online: https://digital.csic.es/handle/10261/127119 (accessed on 1 December 2020).
- Pérez-Jiménez, M.; Pozo-Bayón, M.Á. Development of an in-mouth headspace sorptive extraction method (HSSE) for oral aroma monitoring and application to wines of different chemical composition. Food Res. Int. 2019, 121, 97–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fia, G.; Millarini, V.; Granchi, L.; Bucalossi, G.; Guerrini, S.; Zanoni, B.; Rosi, I. Beta-glucosidase and esterase activity from Oenococcus oeni: Screening and evaluation during malolactic fermentation in harsh conditions. LWT 2018, 89, 262–268. [Google Scholar] [CrossRef]
- Narasimha, G.; Sridevi, A.; Ramanjaneyulu, G.; Rajasekhar Reddy, B. Purification and Characterization of β-Glucosidase from Aspergillus niger. Int. J. Food Prop. 2016, 19, 652–661. [Google Scholar] [CrossRef]
- Leite, R.S.R.; Alves-Prado, H.F.; Cabral, H.; Pagnocca, F.C.; Gomes, E.; Da-Silva, R. Production and characteristics comparison of crude β-glucosidases produced by microorganisms Thermoascus aurantiacus e Aureobasidium pullulans in agricultural wastes. Enzym. Microb. Technol. 2008, 43, 391–395. [Google Scholar] [CrossRef]
- Mateo, J.J.; Di Stefano, R. Enological properties of β-glucosidase in wine yeasts. Food Microbiol. 1998, 14, 583–591. [Google Scholar] [CrossRef]
- Maicas, S.; Mateo, J.J. Hydrolysis of terpenyl glycosides in grape juice and other fruit juices: A review. Appl. Microbiol. Biotechnol. 2005, 67, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Spano, G.; Rinaldi, A.; Ugliano, M.; Moio, L.; Beneduce, L.; Massa, S. A β-glucosidase gene isolated from wine Lactobacillus plantarum is regulated by abiotic stresses. J. Appl. Microbiol. 2005, 98, 855–861. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brizuela, N.S.; Arnez-Arancibia, M.; Semorile, L.; Pozo-Bayón, M.Á.; Bravo-Ferrada, B.M.; Tymczyszyn, E.E. β-Glucosidase Activity of Lactiplantibacillus plantarum UNQLp 11 in Different Malolactic Fermentations Conditions: Effect of pH and Ethanol Content. Fermentation 2021, 7, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010022
Brizuela NS, Arnez-Arancibia M, Semorile L, Pozo-Bayón MÁ, Bravo-Ferrada BM, Tymczyszyn EE. β-Glucosidase Activity of Lactiplantibacillus plantarum UNQLp 11 in Different Malolactic Fermentations Conditions: Effect of pH and Ethanol Content. Fermentation. 2021; 7(1):22. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010022
Chicago/Turabian StyleBrizuela, Natalia S., Marina Arnez-Arancibia, Liliana Semorile, María Ángeles Pozo-Bayón, Bárbara M. Bravo-Ferrada, and E. Elizabeth Tymczyszyn. 2021. "β-Glucosidase Activity of Lactiplantibacillus plantarum UNQLp 11 in Different Malolactic Fermentations Conditions: Effect of pH and Ethanol Content" Fermentation 7, no. 1: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010022