Effects of Naphthazarin (DHNQ) Combined with Lawsone (NQ-2-OH) or 1,4-Naphthoquinone (NQ) on the Auxin-Induced Growth of Zea mays L. Coleoptile Segments


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
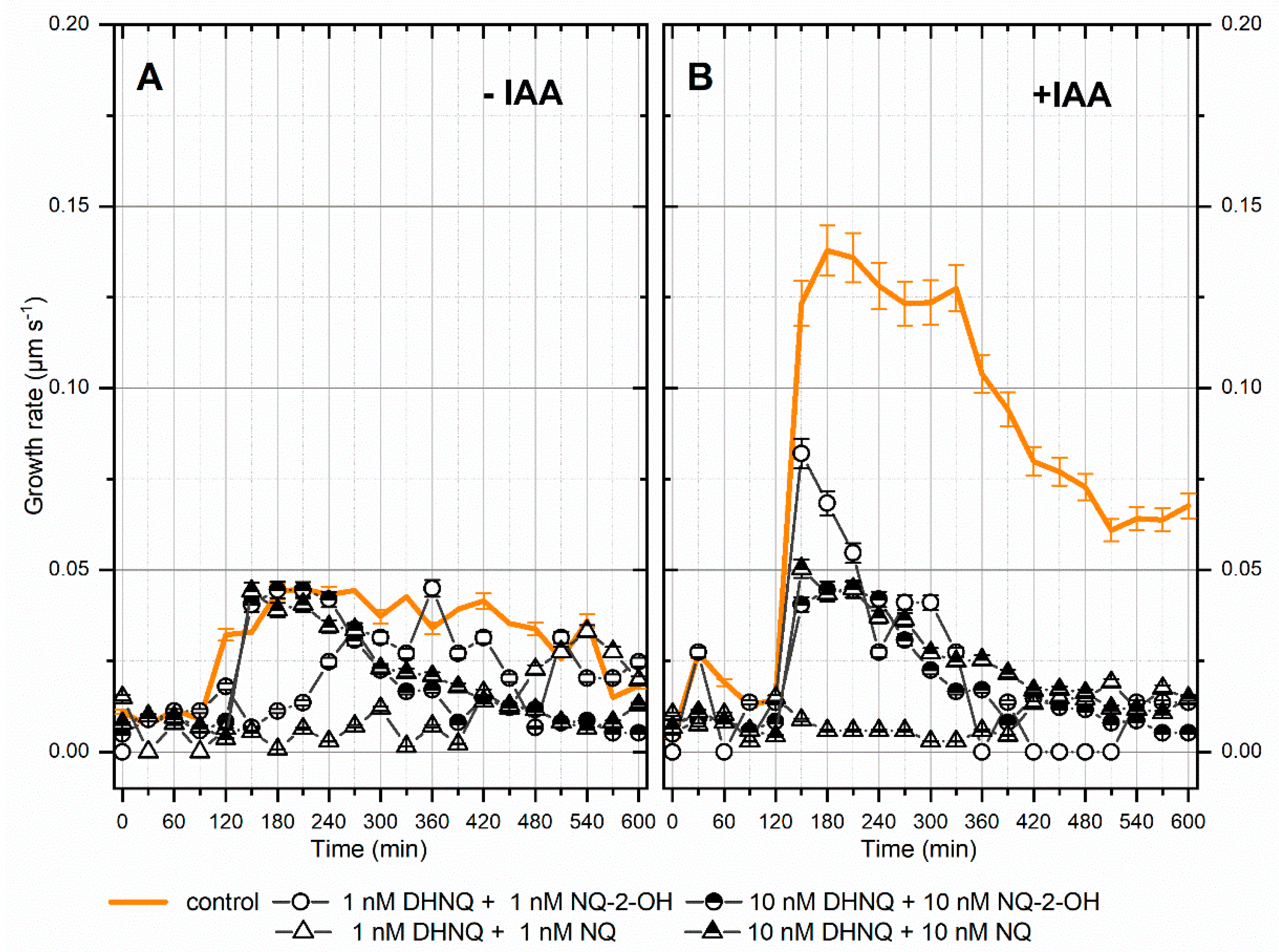
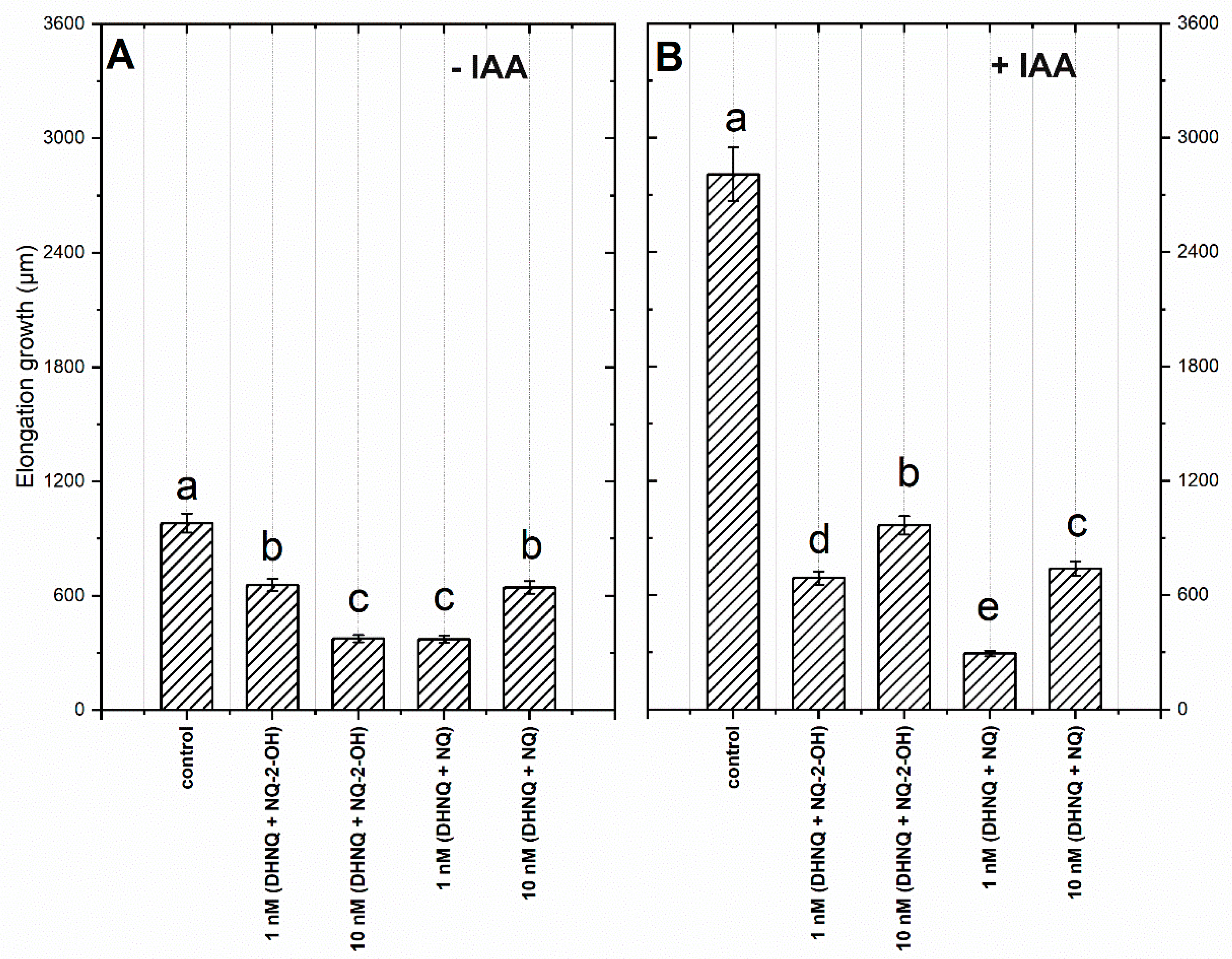
2.1. Effects of Naphthazarin (DHNQ) Combined with Lawsone (NQ-2-OH) or 1,4-Naphthoquinone (NQ) on the Endogenous and Auxin-Induced Growth of Maize Coleoptile Segments
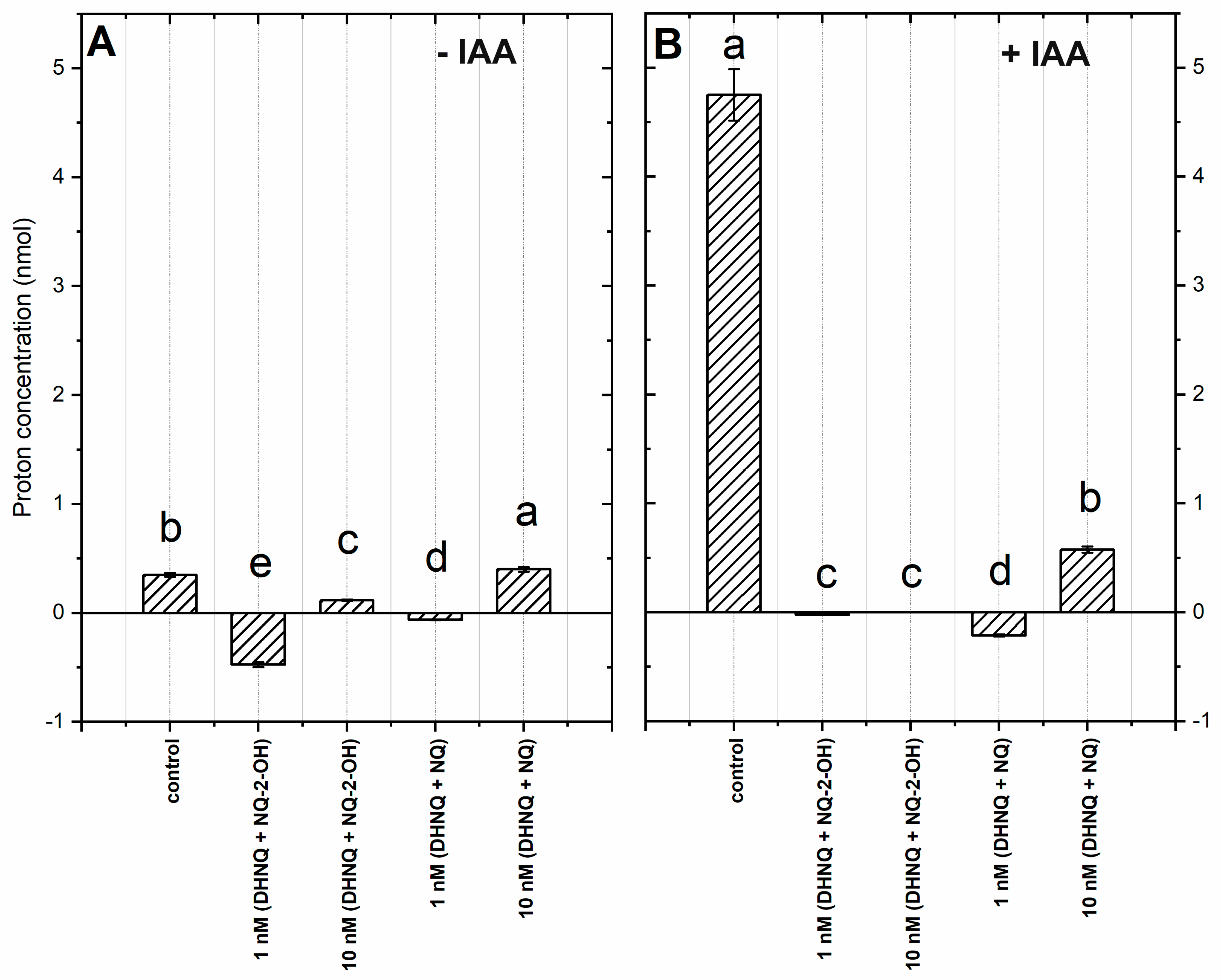
2.2. Proton Concentration in the Incubation Medium of the Coleoptile Segments
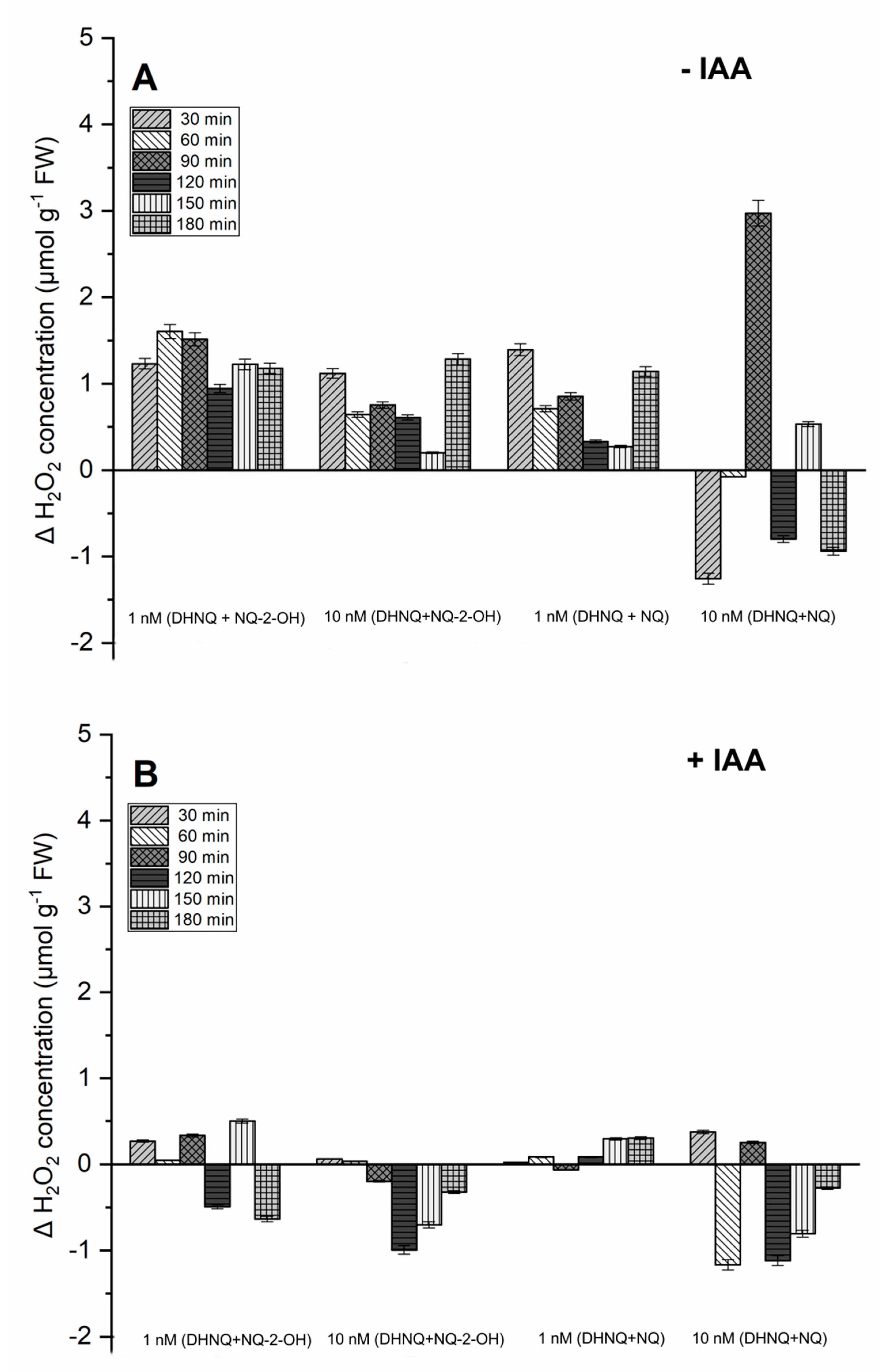
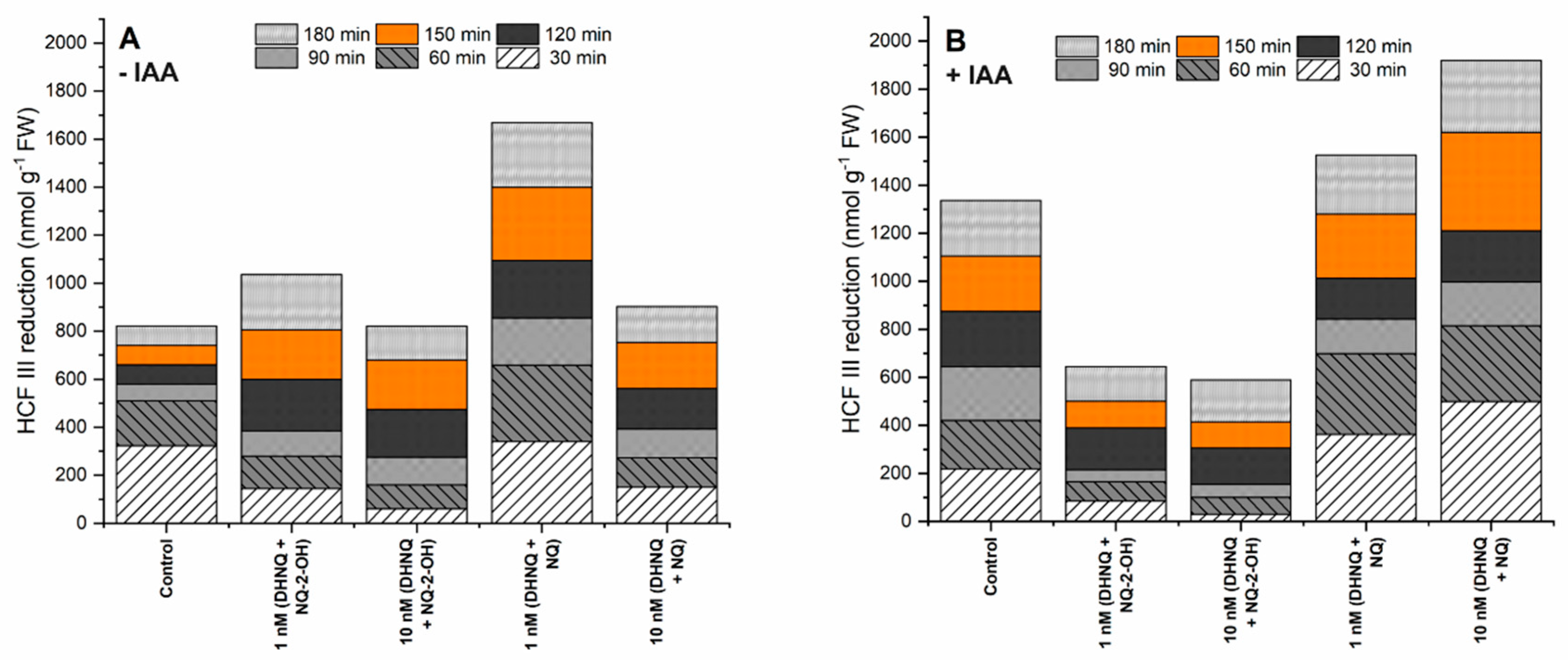
2.3. Effects of Naphthazarin (DHNQ) Combined with Lawsone (NQ-2-OH) or 1,4-Naphthoquinone (NQ) on Oxidative Stress, Measured as the Production of H2O2 and Redox Activity
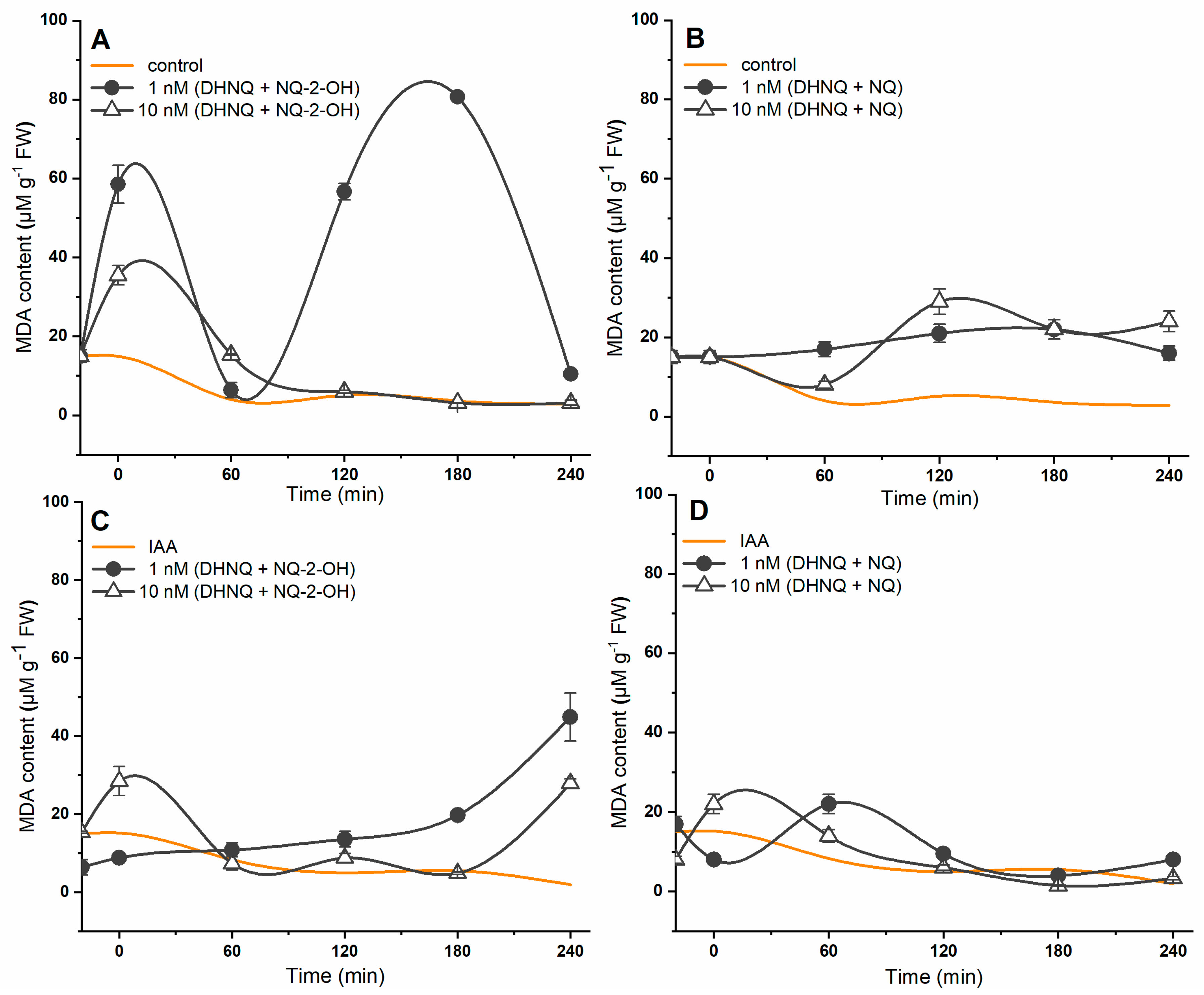
2.4. Effects of Naphthazarin (DHNQ) Combined with Lawsone (NQ-2-OH) or 1,4-Naphthoquinone (NQ) on Lipid Peroxidation and Cell Viability
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Growth and pH Measurements
4.3. Hydrogen Peroxide Detection
4.4. Redox Activity
4.5. Lipid Peroxidation
4.6. Viability
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DHNQ | 5,8-dihydroxy-1,4-naphthoquinone |
| NQ-2-OH | 2-hydroxy-1,4-naphthoquinone |
| NQ | 1,4-naphthoquinone |
| IAA | Indole acetic acid |
References
- Paque, S.; Weijers, D. Q&A: Auxin: The plant molecule that influences almost anything. BMC Boil. 2016, 14, 67. [Google Scholar]
- Gray, W.M. Hormonal Regulation of Plant Growth and Development. PLoS Boil. 2004, 2, e311. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Bai, Y.; Shen, C.; Wu, Y.; Zhang, S.; Jiang, D.; Guilfoyle, T.J.; Chen, M.; Qi, Y. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genom. 2010, 10, 533–546. [Google Scholar] [CrossRef]
- Wang, S.; Fu, J. Insights into Auxin Signaling in Plant–Pathogen Interactions. Front. Plant Sci. 2011, 2, 74. [Google Scholar]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef]
- Zandomeni, K.; Schopfer, P. Reorientation of microtubules at the outer epidermal wall of maize coleoptiles by phytochrome, blue-light photoreceptor, and auxin. Protoplasma 1993, 173, 103–112. [Google Scholar] [CrossRef]
- Philippar, K.; Fuchs, I.; Lüthen, H.; Hoth, S.; Bauer, C.S.; Haga, K.; Thiel, G.; Ljung, K.; Sandberg, G.; Böttger, M. Auxin-induced K+ channel expression represents an essential step in coleoptile growth and gravitropism. Proc. Natl. Acad. Sci. USA 1999, 96, 12186–12191. [Google Scholar] [CrossRef]
- Becker, D.; Hedrich, R. Channelling auxin action: Modulation of ion transport by indole-3-acetic acid. Plant Mol. Boil. 2002, 49, 349–356. [Google Scholar] [CrossRef]
- Chan, J. Microtubule and cellulose microfibril orientation during plant cell and organ growth. J. Microsc. 2012, 247, 23–32. [Google Scholar] [CrossRef]
- Burdach, Z.; Kurtyka, R.; Siemieniuk, A.; Karcz, W. Role of chloride ions in the promotion of auxin-induced growth of maize coleoptile segments. Ann. Bot. 2014, 114, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Dharmasiri, N.; Estelle, M. Auxin signaling and regulated protein degradation. Trends Plant Sci. 2004, 9, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 441. [Google Scholar] [CrossRef]
- dos Santos Maraschin, F.; Memelink, J.; Offringa, R. Auxin-induced, SCFTIR1-mediated poly-ubiquitination marks AUX/IAA proteins for degradation. Plant J. 2009, 59, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Weijers, D.; Wagner, D. Transcriptional responses to the auxin hormone. Annu. Rev. Plant Boil. 2016, 67, 539–574. [Google Scholar] [CrossRef]
- Papageorgiou, V.P.; Assimopoulou, A.N.; Couladouros, E.A.; Hepworth, D.; Nicolaou, K. The chemistry and biology of alkannin, shikonin, and related naphthazarin natural products. Angew. Chem. Int. Ed. 1999, 38, 270–301. [Google Scholar] [CrossRef]
- Babula, P.; Adam, V.; Havel, L.; Kizek, R. Noteworthy secondary metabolites naphthoquinones-their occurrence, pharmacological properties and analysis. Curr. Pharm. Anal. 2009, 5, 47–68. [Google Scholar] [CrossRef]
- Devi, S.P.; Kumaria, S.; Rao, S.R.; Tandon, P. Carnivorous Plants as a Source of Potent Bioactive Compound: Naphthoquinones. Trop. Plant Boil. 2016, 9, 267–279. [Google Scholar] [CrossRef]
- Widhalm, J.R.; Rhodes, D. Biosynthesis and molecular actions of specialized 1,4-naphthoquinone natural products produced by horticultural plants. Hortic. Res. 2016, 3, 16046. [Google Scholar] [CrossRef] [Green Version]
- Duffey Sean, S.; Stout Michael, J. Antinutritive and toxic components of plant defense against insects. Arch. Insect Biochem. Physiol. 1996, 32, 3–37. [Google Scholar] [CrossRef]
- Eilenberg, H.; Zilberstein, A. Carnivorous pitcher plants–towards understanding the molecular basis of prey digestion. Floric. Ornam. Plant Biotechnol. Adv. Top. Issues 2008, 5, 287–294. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Kot, M.; Karcz, W.; Zaborska, W. 5-Hydroxy-1,4-naphthoquinone (juglone) and 2-hydroxy-1,4-naphthoquinone (lawsone) influence on jack bean urease activity: Elucidation of the difference in inhibition activity. Bioorg. Chem. 2010, 38, 132–137. [Google Scholar] [CrossRef]
- Klotz, L.-O.; Hou, X.; Jacob, C. 1,4-naphthoquinones: From oxidative damage to cellular and inter-cellular signaling. Molecules 2014, 19, 14902–14918. [Google Scholar] [CrossRef]
- Oda, Y.; Nakashima, S.; Kondo, E.; Nakamura, S.; Yano, M.; Kubota, C.; Masumoto, Y.; Hirao, M.; Ogawa, Y.; Matsuda, H. Comparison of lawsone contents among Lawsonia inermis plant parts and neurite outgrowth accelerators from branches. J. Nat. Med. 2018, 72, 890–896. [Google Scholar] [CrossRef]
- Sherif, E.; Park, S.-M. Effects of 1,4-naphthoquinone on aluminum corrosion in 0.50 M sodium chloride solutions. Electrochim. Acta 2006, 51, 1313–1321. [Google Scholar] [CrossRef]
- Ostovari, A.; Hoseinieh, S.; Peikari, M.; Shadizadeh, S.; Hashemi, S. Corrosion inhibition of mild steel in 1 M HCl solution by henna extract: A comparative study of the inhibition by henna and its constituents (Lawsone, Gallic acid, α-d-Glucose and Tannic acid). Corros. Sci. 2009, 51, 1935–1949. [Google Scholar] [CrossRef]
- Camara, C.A.; Silva, T.; Da-Silva, T.G.; Martins, R.M.; Barbosa, T.P.; Pinto, A.C.; Vargas, M.D. Molluscicidal activity of 2-hydroxy-[1,4] naphthoquinone and derivatives. An. Acad. Bras. Ciênc. 2008, 80, 329–334. [Google Scholar] [CrossRef]
- Sundriyal, S.; Viswanad, B.; Bharathy, E.; Ramarao, P.; Chakraborti, A.K.; Bharatam, P.V. New PPARγ ligands based on 2-hydroxy-1,4-naphthoquinone: Computer-aided design, synthesis, and receptor-binding studies. Bioorg. Med. Chem. Lett. 2008, 18, 3192–3195. [Google Scholar] [CrossRef]
- Yang, W.; Du, D.-M. Chiral Squaramide-Catalyzed Highly Enantioselective Michael Addition of 2-Hydroxy-1,4-naphthoquinones to Nitroalkenes. Adv. Synth. Catal. 2011, 353, 1241–1246. [Google Scholar] [CrossRef]
- Thorson, M.K.; Puerta, D.T.; Cohen, S.M.; Barrios, A.M. Inhibition of the lymphoid tyrosine phosphatase: The effect of zinc (II) ions and chelating ligand fragments on enzymatic activity. Bioorg. Med. Chem. Lett. 2014, 24, 4019–4022. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Shi, D.; Hu, G.; Zhang, B.; Li, X.; Liu, R.; Han, X.; Yao, X.; Fang, J. Synthesis of naphthazarin derivatives and identification of novel thioredoxin reductase inhibitor as potential anticancer agent. Eur. J. Med. Chem. 2017, 140, 435–447. [Google Scholar] [CrossRef]
- Hijji, Y.M.; Barare, B.; Zhang, Y. Lawsone (2-hydroxy-1,4-naphthoquinone) as a sensitive cyanide and acetate sensor. Sens. Actuators B Chem. 2012, 169, 106–112. [Google Scholar] [CrossRef]
- Yao, M.; Umetani, S.; Ando, H.; Kiyobayashi, T.; Takeichi, N.; Kondo, R.; Takeshita, H.T. Rechargeable organic batteries using chloro-substituted naphthazarin derivatives as positive electrode materials. J. Mater. Sci. 2017, 52, 12401–12408. [Google Scholar] [CrossRef]
- Miroshnikov, M.; Kato, K.; Babu, G.; Divya, K.P.; Arava, L.M.R.; Ajayan, P.M.; John, G. A common tattoo chemical for energy storage: Henna plant-derived naphthoquinone dimer as a green and sustainable cathode material for Li-ion batteries. RSC Adv. 2018, 8, 1576–1582. [Google Scholar] [CrossRef]
- Macias-Rubalcava, M.L.; Ruiz-Velasco Sobrino, M.E.; Melendez-Gonzalez, C.; Hernandez-Ortega, S. Naphthoquinone spiroketals and organic extracts from the endophytic fungus Edenia gomezpompae as potential herbicides. J. Agric. Food Chem. 2014, 62, 3553–3562. [Google Scholar] [CrossRef]
- Rudnicka, M.; Polak, M.; Karcz, W. Cellular responses to naphthoquinones: Juglone as a case study. Plant Growth Regul. 2014, 72, 239–248. [Google Scholar] [CrossRef]
- Rudnicka, M.; Ludynia, M.; Karcz, W. A comparison of the effects of 1,4-naphthoquinone and 2-hydroxy-1,4-naphthoquinone (lawsone) on indole-3-acetic acid (IAA)-induced growth of maize coleoptile cells. Plant Growth Regul. 2018, 84, 107–122. [Google Scholar] [CrossRef]
- Rayle, D.L.; Cleland, R.E. The Acid Growth Theory of auxin-induced cell elongation is alive and well. Plant Physiol. 1992, 99, 1271–1274. [Google Scholar] [CrossRef]
- Hager, A. Role of the plasma membrane H+-ATPase in auxin-induced elongation growth: Historical and new aspects. J. Plant Res. 2003, 116, 483–505. [Google Scholar] [CrossRef]
- Kurtyka, R.; Pokora, W.; Tukaj, Z.; Karcz, W. Effects of juglone and lawsone on oxidative stress in maize coleoptile cells treated with IAA. AoB Plants 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Zaborska, W.; Kot, M.; Bala, A. Kinetics of jack bean urease inhibition by 2, 3-dichloro-1,4-naphthoquinone. Elucidation of the mechanism: Redox cycling and sulfhydryl arylation. J. Enzym. Inhib. Med. Chem. 2009, 24, 1082–1087. [Google Scholar] [CrossRef]
- Vaverkova, V.; Vrana, O.; Adam, V.; Pekarek, T.; Jampilek, J.; Babula, P. The study of naphthoquinones and their complexes with DNA by using Raman spectroscopy and surface enhanced Raman spectroscopy: New insight into interactions of DNA with plant secondary metabolites. BioMed Res. Int. 2014, 2014, 461393. [Google Scholar] [CrossRef]
- Rudnicka, M.; Ludynia, M.; Karcz, W. The effect of naphthazarin on the growth, electrogenicity, oxidative stress, and microtubule array in Z. mays Coleoptile Cells Treated With IAA. Front. Plant Sci. 2019, 9, 1940. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. Plant Physiol. 2014, 166, 1090. [Google Scholar] [CrossRef]
- Petrov, V.V.; Pardo, J.P.; Slayman, C.W. Reactive cysteines of the Yeast plasma-membrane H+-ATPase (PMA1) mapping the sites of inactivation by N-ethylmaleimide. J. Boil. Chem. 1997, 272, 1688–1693. [Google Scholar] [CrossRef]
- Yang, Y.L.; Zhang, F.; He, W.L.; Wang, X.M.; Zhang, L.X. Iron-mediated inhibition of H+-ATPase in plasma membrane vesicles isolated from wheat roots. Cell. Mol. Life Sci. 2003, 60, 1249–1257. [Google Scholar] [CrossRef]
- Janicka-Russak, M.; Kabała, K.; Burzyński, M. Different effect of cadmium and copper on H+-ATPase activity in plasma membrane vesicles from Cucumis sativus roots. J. Exp. Bot. 2012, 63, 4133–4142. [Google Scholar] [CrossRef]
- Lüthen, H.; Bigdon, M.; Böttger, M. Reexamination of the acid growth theory of auxin action. Plant Physiol. 1990, 93, 931–939. [Google Scholar] [CrossRef]
- Kutschera, U. The current status of the acid-growth hypothesis. New Phytol. 1994, 126, 549–569. [Google Scholar] [CrossRef] [Green Version]
- Kutschera, U. Acid growth and plant development. Science 2006, 311, 952. [Google Scholar] [CrossRef]
- Karcz, W.; Burdach, Z. A comparison of the effects of IAA and 4-Cl-IAA on growth, proton secretion and membrane potential in maize coleoptile segments. J. Exp. Bot. 2002, 53, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Shabala, S. Salinity and programmed cell death: Unravelling mechanisms for ion specific signalling. J. Exp. Bot. 2009, 60, 709–712. [Google Scholar] [CrossRef]
- Rodríguez-Fernández, T.; Ugalde-Saldívar, V.M.; González, I.; Escobar, L.I.; García-Valdés, J. Electrochemical strategy to scout 1,4-naphthoquinones effect on voltage gated potassium channels. Bioelectrochemistry 2012, 86, 1–8. [Google Scholar] [CrossRef]
- Ollinger, K.; Brunmark, A. Effect of hydroxy substituent position on 1,4-naphthoquinone toxicity to rat hepatocytes. J. Boil. Chem. 1991, 266, 21496–21503. [Google Scholar]
- Brunmark, A.; Cadenas, E. Redox and addition chemistry of quinoid compounds and its biological implications. Free Radic. Boil. Med. 1989, 7, 435–477. [Google Scholar] [CrossRef]
- Elzenga, J.T.M.; Staal, M.; Prins, H.B. ATPase activity of isolated plasma membrane vesicles of leaves of Elodea as affected by thiol reagents and NADH/NAD+ ratio. Physiol. Plant. 1989, 76, 379–385. [Google Scholar] [CrossRef]
- Hager, A.; Lanz, C. Essential sulfhydryl groups in the catalytic center of the tonoplast H+-ATPase from coleoptiles of Zea mays L. as demonstrated by the biotin-streptavidin-peroxidase system. Planta 1989, 180, 116–122. [Google Scholar] [CrossRef]
- Kinoshita, T.; Nishimura, M.; Shimazaki, K. Cytosolic concentration of Ca2+ regulates the plasma membrane H+-ATPase in guard cells of fava bean. Plant Cell 1995, 7, 1333–1342. [Google Scholar] [CrossRef]
- Polevoi, V.; Sinyutina, N.; Salamatova, T.; Inge-Vechtomova, N.; Tankelyun, O.; Sharova, E.; Shishova, M. Mechanism of auxin action: Second messengers. In Plant Hormone Signal Perception and Transduction; Springer: Dordrecht, The Netherlands, 1996; pp. 223–231. [Google Scholar]
- Dat, J.; Vandenabeele, S.; Vranová, E.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef]
- Brault, M.; Amiar, Z.; Pennarun, A.-M.; Monestiez, M.; Zhang, Z.; Cornel, D.; Dellis, O.; Knight, H.; Bouteau, F.; Rona, J.-P. Plasma membrane depolarization induced by abscisic acid in Arabidopsis suspension cells involves reduction of proton pumping in addition to anion channel activation, which are both Ca2+ dependent. Plant Physiol. 2004, 135, 231–243. [Google Scholar] [CrossRef]
- Mori, I.C.; Schroeder, J.I. Reactive oxygen species activation of plant Ca2+ channels. A signaling mechanism in polar growth, hormone transduction, stress signaling, and hypothetically mechanotransduction. Plant Physiol. 2004, 135, 702–708. [Google Scholar] [CrossRef]
- Tian, Q.; Nagpal, P.; Reed, J.W. Regulation of Arabidopsis SHY2/IAA3 protein turnover. Plant J. 2003, 36, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Hennig, L.; Christner, C.; Kipping, M.; Schelbert, B.; Rücknagel, K.P.; Grabley, S.; Küllertz, G.; Fischer, G. Selective inactivation of parvulin-like peptidyl-prolyl cis/trans isomerases by juglone. Biochemistry 1998, 37, 5953–5960. [Google Scholar] [CrossRef]
- Dharmasiri, N.; Dharmasiri, S.; Jones, A.M.; Estelle, M. Auxin action in a cell-free system. Curr. Boil. 2003, 13, 1418–1422. [Google Scholar] [CrossRef]
- Kepinski, S.; Leyser, O. Auxin-induced SCFTIR1–Aux/IAA interaction involves stable modification of the SCFTIR1 complex. Proc. Natl. Acad. Sci. USA 2004, 101, 12381–12386. [Google Scholar] [CrossRef]
- Hejl, A.M.; Koster, K.L. Juglone disrupts root plasma membrane H+-ATPase activity and impairs water uptake, root respiration, and growth in soybean (Glycine max) and corn (Zea mays). J. Chem. Ecol. 2004, 30, 453–471. [Google Scholar] [CrossRef]
- Chaimovitsh, D.; Abu-Abied, M.; Belausov, E.; Rubin, B.; Dudai, N.; Sadot, E. Microtubules are an intracellular target of the plant terpene citral. Plant J. 2010, 61, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Acharya, B.R.; Bhattacharyya, S.; Choudhury, D.; Chakrabarti, G. The microtubule depolymerizing agent naphthazarin induces both apoptosis and autophagy in A549 lung cancer cells. Apoptosis 2011, 16, 924–939. [Google Scholar] [CrossRef]
- Polak, M.; Zaborska, W.; Tukaj, Z.; Karcz, W. Effect of thiosulphinates contained in garlic extract on growth, proton fluxes and membrane potential in maize (Zea mays L.) coleoptile segments. Acta Physiol. Plant. 2012, 34, 41–52. [Google Scholar] [CrossRef]
- Karcz, W.; Stolarek, J.; Lekacz, H.; Kurtyka, R.; Burdach, Z. Comparative investigation of auxin and fusicoccin-induced growth and H-extrusion in coleoptile segments of Zea mays L. Acta Physiol. Plant. 1995, 17. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized assay for hydrogen peroxide determination in plant tissue using potassium iodide. Am. J. Anal. Chem. 2014, 5, 730. [Google Scholar] [CrossRef]
- Federico, R.; Giartosio, C.E. A transplasmamembrane electron transport system in maize roots. Plant Physiol. 1983, 73, 182–184. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Hunter, C.T.; Kirienko, D.H.; Sylvester, A.W.; Peter, G.F.; McCarty, D.R.; Koch, K.E. Cellulose Synthase-Like D1 is integral to normal cell division, expansion, and leaf development in maize. Plant Physiol. 2011, 158, 708–724. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudnicka, M.; Ludynia, M.; Karcz, W. Effects of Naphthazarin (DHNQ) Combined with Lawsone (NQ-2-OH) or 1,4-Naphthoquinone (NQ) on the Auxin-Induced Growth of Zea mays L. Coleoptile Segments. Int. J. Mol. Sci. 2019, 20, 1788. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071788
Rudnicka M, Ludynia M, Karcz W. Effects of Naphthazarin (DHNQ) Combined with Lawsone (NQ-2-OH) or 1,4-Naphthoquinone (NQ) on the Auxin-Induced Growth of Zea mays L. Coleoptile Segments. International Journal of Molecular Sciences. 2019; 20(7):1788. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071788
Chicago/Turabian StyleRudnicka, Małgorzata, Michał Ludynia, and Waldemar Karcz. 2019. "Effects of Naphthazarin (DHNQ) Combined with Lawsone (NQ-2-OH) or 1,4-Naphthoquinone (NQ) on the Auxin-Induced Growth of Zea mays L. Coleoptile Segments" International Journal of Molecular Sciences 20, no. 7: 1788. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071788