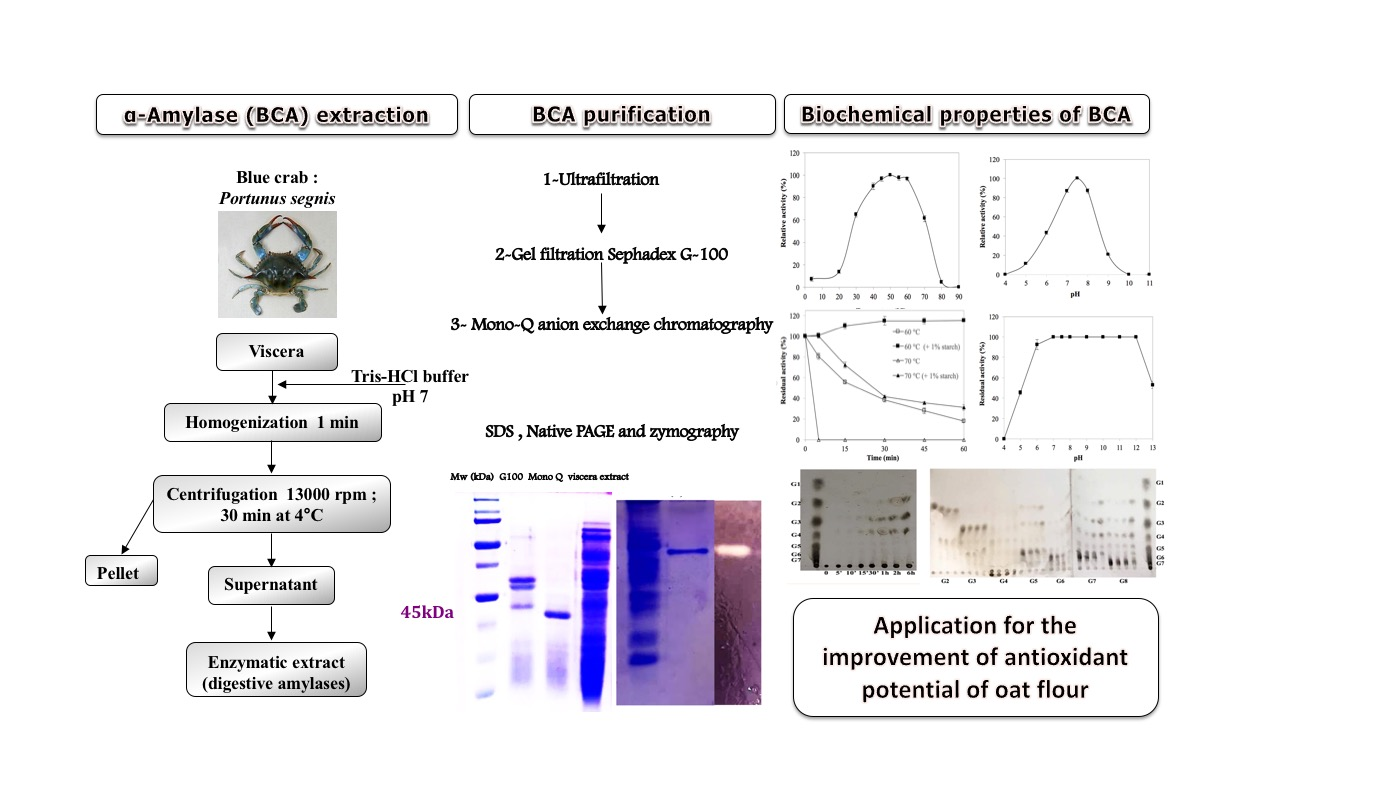
A Novel Digestive α-Amylase from Blue Crab (Portunus segnis) Viscera: Purification, Biochemical Characterization and Application for the Improvement of Antioxidant Potential of Oat Flour



Abstract
:
1. Introduction
2. Results
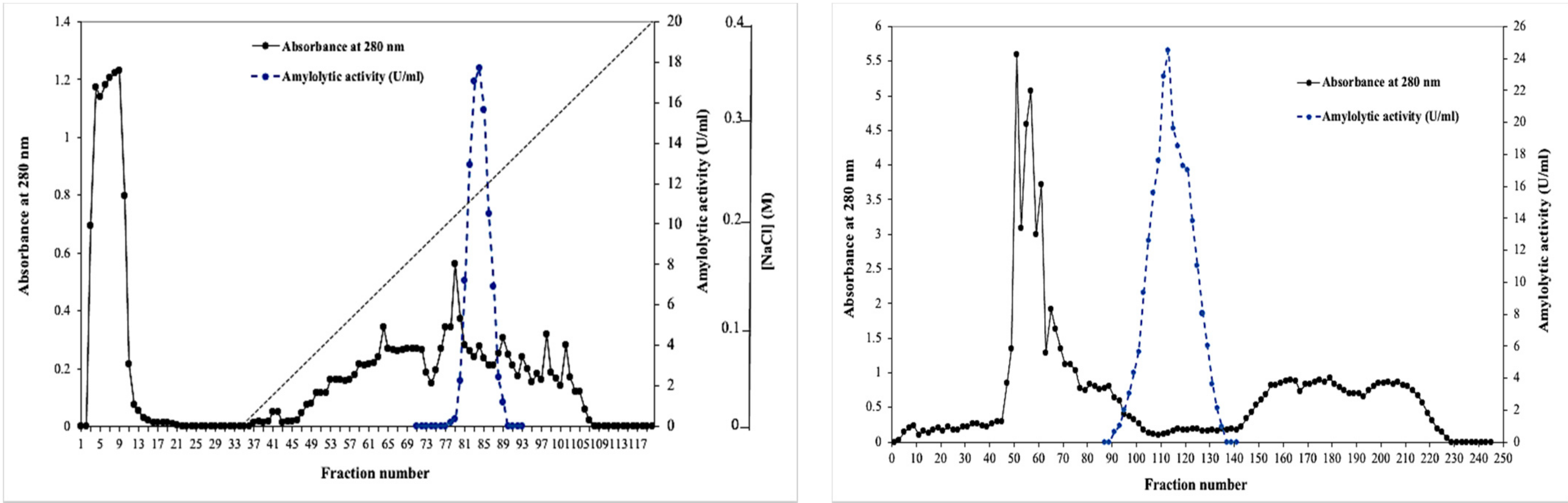
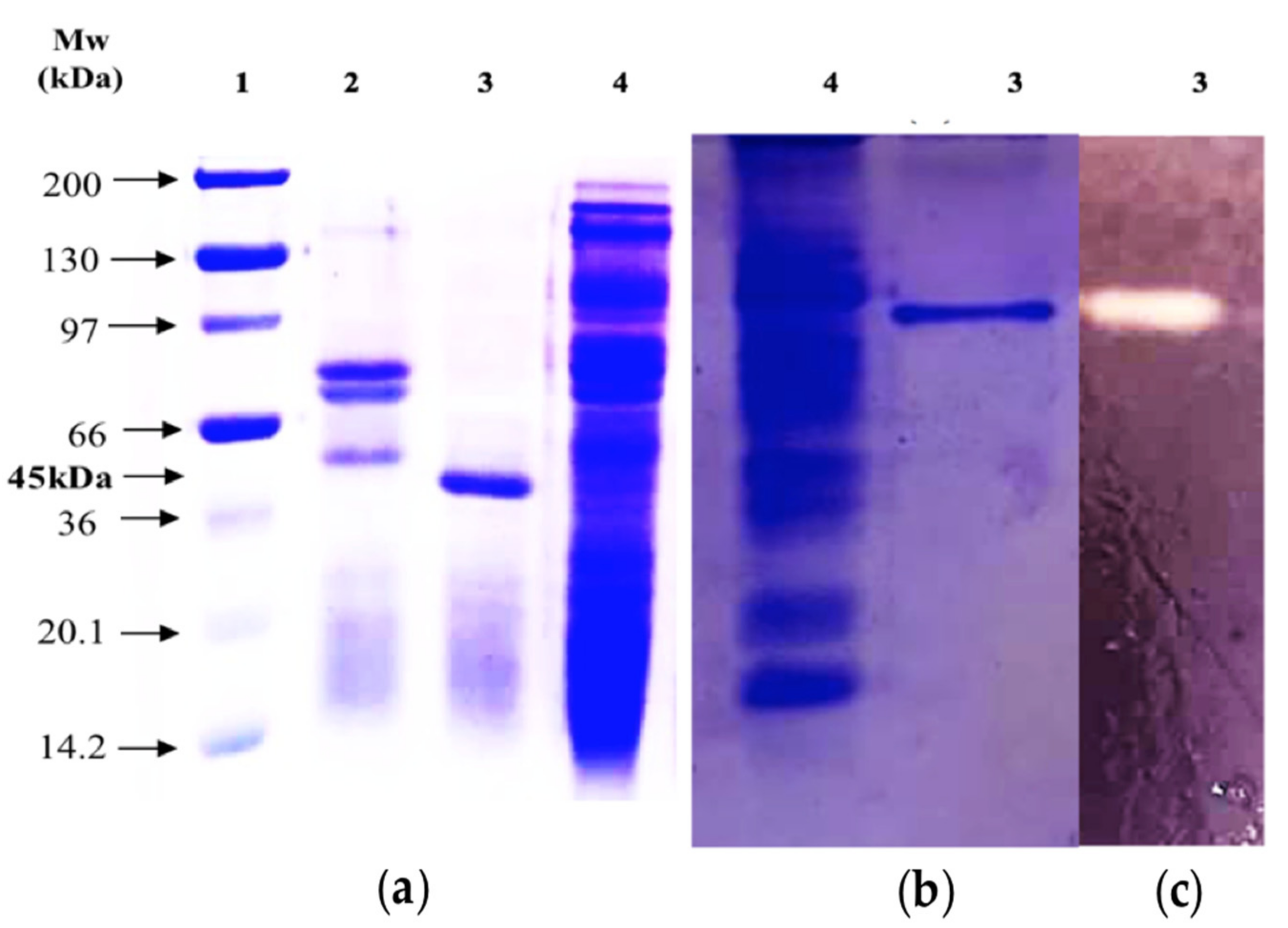
2.1. Purification of the BCA α-Amylase
2.2. Biochemical Characterization of the Purified BCA
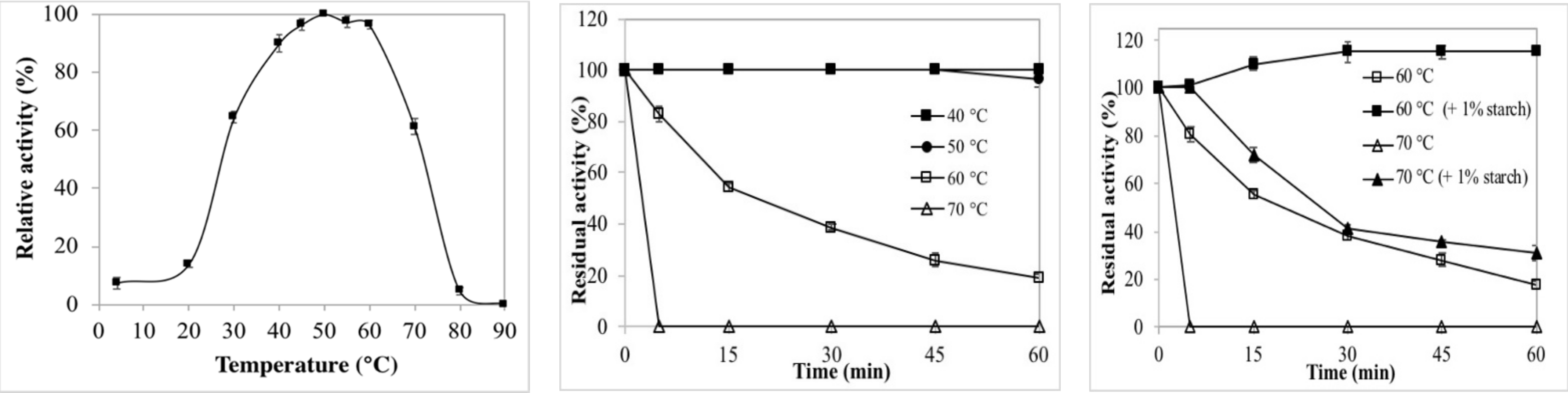
2.2.1. Effects of Temperature on Enzyme Activity and Stability
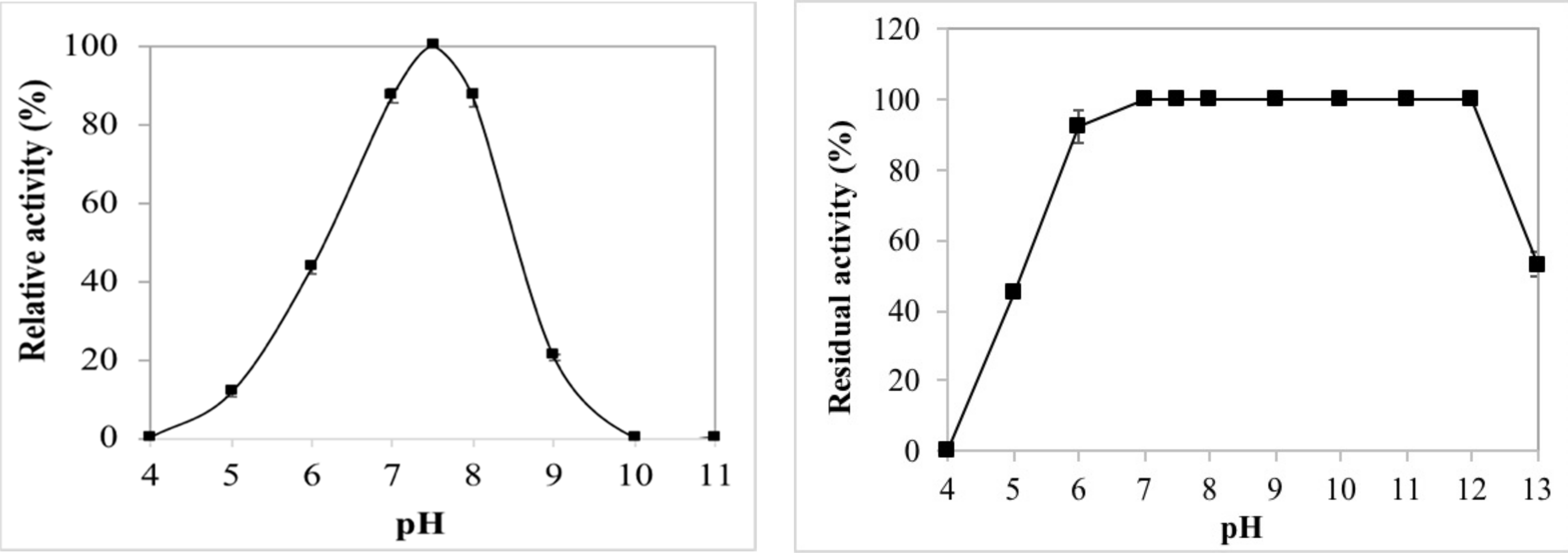
2.2.2. Effects of pH on Enzyme Activity and Stability
2.2.3. Effects of Metal Ions on BCA Activity
2.2.4. Effects of Surfactants and Enzyme Inhibitors on BCA Stability
2.3. Kinetic Study and Thin-Layer Chromatographic (TLC) Analysis
2.3.1. Substrate Specificity Profile
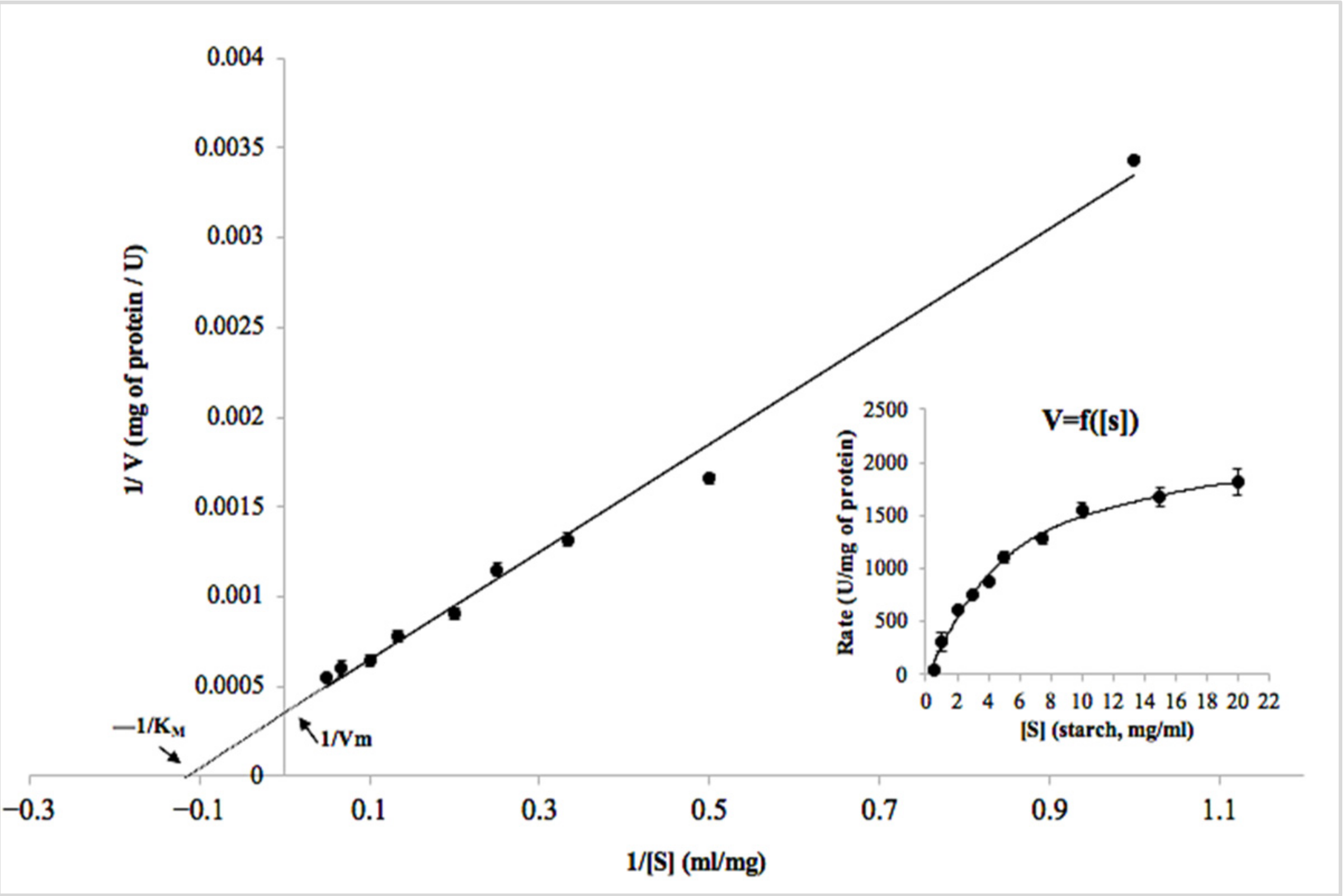
2.3.2. Determination of Kinetic Parameters
2.3.3. TLC Analysis of Starch and Maltooligosaccharides Hydrolysis Products
2.4. Storage Stability
2.5. Application of Purified BCA for the Improvement of Antioxidant Potential of Oat
3. Materials and Methods
3.1. Obtaining the Raw Materials
3.2. Preparation of Crude Digestive Enzyme Extract
3.3. Protein Concentration and α-Amylase Activity Assays
3.4. Enzyme Purification Procedure
3.4.1. Ultrafiltration
3.4.2. Sephadex G-100 Chromatography
3.4.3. Anion Exchange Chromatography
3.4.4. Sodium Dodecyl Sulphate–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
3.4.5. Native PAGE and Zymography
3.5. Biochemical Characterization of the Purified BCA
3.5.1. Effects of Temperature on Enzyme Activity and Stability
3.5.2. Effects of pH on Enzyme Activity and Stability
3.5.3. Effects of Metallic Ions on Enzyme Activity
3.5.4. Effects of Inhibitors and Surfactants on Enzyme Stability
3.6. Kinetic Study and Thin-Layer Chromatographic Analysis
3.6.1. Substrate Specificity Profile
3.6.2. Kinetic Parameters Determination
3.6.3. TLC Analysis of Starch and Maltooligosaccharides Hydrolysis Products
3.7. Storage Stability
3.8. Application of Purified BCA for the Improvement of Antioxidant Potential of Oat
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coutinho, P.M.; Henrissat, B. Carbohydrate-activeenzymes: An integrated database approach. In Recent Advances in Carbohydrate Bioengineering; Gilbert, H.J., Davies, G., Henrissat, H., Svensson, B., Eds.; The Royal Society of Chemistry: Cambridge, UK, 1999; pp. 3–12. [Google Scholar]
- Zhang, Q.; Han, Y.; Xiao, H. Microbial α-amylase: A biomolecular overview. Process Biochem. 2017, 53, 88–101. [Google Scholar]
- Homaei, A.; Ghanbarzadeh, M.; Monsef, F. Biochemical features and kinetic properties of α-amylases from marine organisms. Int. J. Biol. Macromol. 2016, 83, 306–314. [Google Scholar]
- Elleuche, S.; Schröder, C.; Sahm, K.; Antranikian, G. Extremozymes—Biocatalysts with unique properties from extremophilic microorganisms. Curr. Opin. Biotechnol. 2014, 29, 116–123. [Google Scholar]
- Villamil, O.; Váquiro, H.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar]
- Siddik, M.A.B.; Howieson, J.; Fotedar, R.; Partridge, G.J. Enzymatic fish protein hydrolysates in finfish aquaculture: A review. Rev. Aquac. 2020, 13, 406–430. [Google Scholar]
- Al Khawli, F.; Pateiro, M.; Domínguez, R.; Lorenzo, J.M.; Gullón, P.; Kousoulaki, K.; Barba, F.J. Innovative green technologies of intensification for valorization of seafood and their by-products. Mar. Drugs 2019, 17, 689. [Google Scholar]
- Wang, Y.; Song, Q.; Zhang, X.H. Marine microbiological enzymes: Studies with multiple strategies and prospects. Mar. Drugs 2016, 14, 171. [Google Scholar]
- Morrissey, M.T.; Okada, T. Marine enzymes from seafood by-products. In Maximizing the Value of Marine by-Products; Woodhead Publishing: Sawston, UK, 2007; pp. 374–396. [Google Scholar]
- De Melo Oliveira, V.; da Cunha, M.N.C.; Assis, C.R.D.; de Batista, J.M.; Nascimento, T.P.; Santos, J.F.; Porto, A.L.F. Separation and partial purification of collagenolytic protease from peacock bass (Cichla ocellaris) using different protocol: Precipitation and partitioning approaches. Biocatal. Agric. Biotechnol. 2020, 24, 101509. [Google Scholar]
- Assis, C.R.D.; de Melo Oliveira, V.; Silva, J.C.; Silva, Q.J.; Bezerra, R.d.; Porto, A.L.F. Recovery of fibrinolytic and collagenolytic enzymes from fish and shrimp byproducts: Potential source for biomedical applications. Boletim Inst. Pesca 2019, 45, 1. [Google Scholar]
- Lassoued, I.; Hajji, S.; Mhamdi, S.; Jridi, M.; Bayoudh, A.; Barkia, A.; Nasri, M. Digestive alkaline proteases from thornback ray (Raja clavata): Characteristics and applications. Int. J. Biol. Macromol. 2015, 80, 668–675. [Google Scholar]
- Tiwari, S.P.; Srivastava, R.; Singh, C.S.; Shukla, K.; Singh, R.K.; Singh, P.; Sharma, R. Amylases: An overview with special reference to alpha amylase. J. Glob. Biosci. 2015, 4, 1886–1901. [Google Scholar]
- Aiyer, P.V. Amylases and their applications. Afr. J. Biotechnol. 2015, 4, 1525–1529. [Google Scholar]
- Mezeal, I.A.; Mezil, S.A.; Shnewer, G.J. Presentation Uses of Amylase in Manufacturing. Int. J. Pharm. Qual. Assur. 2019, 10, 85–89. [Google Scholar]
- Gupta, R.; Gigras, P.; Mohapatra, H.; Goswami, V.K.; Chauhan, B. Microbial α-amylases: A biotechnological perspective. Process. Biochem. 2019, 38, 1599–1616. [Google Scholar]
- Kim, S.K. Marine microbial amylases: Properties and applications. In Marine Enzymes Biotechnology: Production and Industrial Applications, Part II—Marine Organisms Producing Enzymes; Elsevier Science: Amsterdam, The Netherlands, 2016; p. 161. [Google Scholar]
- Sindhu, R.; Binod, P.; Madhavan, A.; Beevi, U.S.; Mathew, A.K.; Abraham, A.; Kumar, V. Molecular improvements in microbial α-amylases for enhanced stability and catalytic efficiency. Bioresour. Technol. 2017, 245, 1740–1748. [Google Scholar]
- Sanchez, A.C.; Ravanal, M.C.; Andrews, B.A.; Asenjo, J.A. Heterologous expression and biochemical characterization of a novel cold-active α-amylase from the Antarctic bacteria Pseudoalteromonas sp. 2-3. Protein Expr. Purif. 2019, 155, 78–85. [Google Scholar]
- Ottoni, J.R.; e Silva, T.R.; de Oliveira, V.M.; Passarini, M.R.Z. Characterization of amylase produced by cold-adapted bacteria from Antarctic samples. Biocatal. Agric. Biotechnol. 2020, 23, 101452. [Google Scholar]
- Peng, H.; Chen, M.; Yi, L.; Zhang, X.; Wang, M.; Xiao, Y.; Zhang, N. Identification and characterization of a novel raw-starch-degrading α-amylase (AmyASS) from the marine fish pathogen Aeromonas salmonicida ssp. Salmonicida. J. Mol. Catal. B Enzym. 2015, 119, 71–77. [Google Scholar]
- Wu, Y.R.; Mao, A.; Sun, C.; Shanmugam, S.; Li, J.; Zhong, M.; Hu, Z. Catalytic hydrolysis of starch for biohydrogen production by using a newly identified amylase from a marine bacterium Catenovulum sp. X3. Int. J. Biol. Macromol. 2017, 104, 716–723. [Google Scholar]
- Wu, G.; Qin, Y.; Cheng, Q.; Liu, Z. Characterization of a novel alkali-stable and salt-tolerant α-amylase from marine bacterium Zunongwangia profundal. J. Mol. Catal. B Enzym. 2014, 110, 8–15. [Google Scholar]
- Khamassi, F.; Ghanem, R.; Khamassi, S.; Dhifallah, F.; Souissi, J.B. Socio-economic impacts of the alien invasive crab Portunus segnis (Forskal, 1775) in the gulf of Gabès. Tunisia. Rapp. Comm. Int. Pour L’exploration Sci. Méditerranée (CIESM) 2019, 42, 253. [Google Scholar]
- Bejaoui, S.; Ghribi, F.; Hatira, S.; Chetoui, I.; Rebah, I. First investigation in the biochemical analysis of the invasive crab Portunus segnis from Tunisian waters. J. Am. Oil Chem. Soc. 2017, 94, 673–682. [Google Scholar]
- Tsao, C.Y.; Hsu, Y.H.; Chao, L.M.; Jiang, S.T. Purification and characterization of three amylases from viscera of hard clam Meretrix lusoria. Fish. Sci. 2004, 70, 174–182. [Google Scholar]
- Hsieh, M.S.; Yin, L.J.; Jiang, S.T. Purification and characterization of the amylase from a small abalone Haliotis sieboldii. Fish. Sci. 2008, 74, 425–432. [Google Scholar]
- Hara, K.; Ishihara, T.; Yasuda, M. Studies of amylase from crystalline style of short-necked clam-I: Purification and properties of amylase. Nippon Suisan Gakkaishi 1979, 45, 1005–1012. [Google Scholar]
- Feller, G.; Lonhienne, T.; Deroanne, C.; Libioulle, C.; van Beeumen, J.; Gerday, C. Purification, characterization, and nucleotide sequence of the thermolabile alpha-amylase from the antarctic psychrotroph Alteromonas haloplanctis A23. J. Biol. Chem. 1992, 267, 5217–5221. [Google Scholar]
- Mayzaud, O. Purification and kinetic properties of the α-amylase from the copepod Acartia clausi (Giesbrecht, 1889). Comp. Biochem. Physiol. Part B Comp. Biochem. 1985, 82, 725–730. [Google Scholar]
- Tsao, C.Y.; Pan, Y.Z.; Jiang, S.T. Purification and characterization of amylases from small abalone (Sulculus diversicolor aquatilis). J. Agric. Food Chem. 2003, 51, 1064–1070. [Google Scholar]
- Yamada, A.; Takano, K.; Kamoi, I. Purification and properties of amylases from tilapia intestine. Nippon Suisan Gakkaishi 1991, 57, 1903–1909. [Google Scholar]
- Banerjee, G.; Mukherjee, S.; Bhattacharya, S.; Ray, A.K. Purification and characterization of extracellular protease and amylase produced by the bacterial strain, Corynebacterium alkanolyticum ATH3 isolated from fish gut. Arab. J. Sci. Eng. 2016, 41, 9–16. [Google Scholar]
- Maugle, P.D.; Simpson, K.L. Digestive enzymes of the shrimp Penaeus japonicus-I. Characteristics of amylase and protease of the shrimp Penaeus japonicus. Comp. Biochem. Physiol. 1982, 48, 1753–1757. [Google Scholar]
- Yamada, A.; Takano, K.; Kamoi, I. Purification and properties of amylase from tilapia stomach. Nippon Suisan Gakkaishi 1996, 62, 269–274. [Google Scholar]
- Sabapathy, U.; Teo, L.H. A kinetic study of the a-amylase from the digestive gland of Perna viridis L. Comp. Biochem. Physiol. 1992, 101B, 73–77. [Google Scholar]
- Hmidet, N.; Bougatef, A.; Nasri, M. Fish Amylolytic Enzymes: Biochemical characterization and application for oligosaccharides production. Nat. Prod. J. 2013, 3, 125–130. [Google Scholar]
- Fang, W.; Xue, S.; Deng, P.; Zhang, X.; Wang, X.; Xiao, Y.; Fang, Z. AmyZ1: A novel α-amylase from marine bacterium Pontibacillus sp. ZY with high activity toward raw starches. Biotechnol. Biofuels 2019, 12, 95. [Google Scholar]
- Lim, S.J.; Oslan, S.N.H.; Oslan, S.N. Purification and characterization of thermostable α-amylases from microbial sources. BioResources 2020, 15, 2005–2029. [Google Scholar]
- Zhang, J.W.; Zeng, R.Y. Purification and characterization of a cold-adapted α-amylase produced by Nocardiopsis sp. 7326 isolated from Prydz Bay. Antarctic. Mar. Biotechnol. 2008, 10, 75–82. [Google Scholar]
- Lu, M.; Wang, S.; Fang, Y.; Li, H.; Liu, S.; Liu, H. Cloning, expression, purification, and characterization of cold-adapted α-amylase from Pseudoalteromonas arctica GS230. Protein J. 2010, 29, 591–597. [Google Scholar]
- Maalej, H.; Hmidet, N.; Ghorbel-Bellaaj, O.; Nasri, M. Purification and biochemical characterization of a detergent stable α-amylase from Pseudomonas stutzeri AS22. Biotechnol. Bioprocess Eng. 2013, 18, 878–887. [Google Scholar]
- Koyama, K.; Shono, J.; Taguchi, H.; Toriba, A.; Hayakawa, K. Effect of Starch on the Inactivation of Amylase in Starch-Containing Foods. Food Sci. Technol. Res. 2013, 19, 989–993. [Google Scholar]
- Wu, X.; Wang, Y.; Tong, B.; Chen, X.; Chen, J. Purification and biochemical characterization of a thermostable and acid-stable alpha-amylase from Bacillus licheniformis B4-423. Int. J. Biol. Macromol. 2018, 109, 329–337. [Google Scholar]
- Fernandez, I.; Moyano, F.J.; Dıaz, M.; Martınez, T. Characterization of α-amylase activity in five species of Mediterranean sparid fishes (Sparidae, Teleostei). J. Exp. Mar. Biol. Ecol. 2001, 262, 1–12. [Google Scholar]
- Vallee, B.L.; Stein, E.A.; Sumerwell, W.N.; Fischer, E.H. Metal content of a-amylase of various organs. J. Biol. Chem. 1959, 234, 2901–2950. [Google Scholar]
- Aygan, A.; Arikan, B.; Korkmaz, H.; Dinçer, S.; Çolak, Ö. Highly thermostable and alkaline α-amylase from a halotolerant-alkaliphilic Bacillus sp. AB68. Braz. J. Microbiol. 2008, 39, 547–553. [Google Scholar]
- Chai, Y.Y.; Rahman, R.N.Z.R.A.; Illias, R.M.; Goh, K.M. Cloning and characterization of two new thermostable and alkalitolerant α-amylases from the Anoxybacillus species that produce high levels of maltose. J. Ind. Microbiol. Biotechnol. 2012, 39, 731–741. [Google Scholar]
- Fincan, S.A.; Enez, B.; Özdemir, S.; Bekler, F.M. Purification and characterization of thermostable α-amylase from thermophilic Anoxybacillus flavithermus. Carbohydr. Polym. 2014, 102, 144–150. [Google Scholar]
- Abd-Elaziz, A.M.; Karam, E.A.; Ghanem, M.M.; Moharam, M.E.; Kansoh, A.L. Production of a novel α-amylase by Bacillus atrophaeus NRC1 isolated from honey: Purification and characterization. Int. J. Biol. Macromol. 2020, 148, 292–301. [Google Scholar]
- Lei, Y.; Peng, H.; Wang, Y.; Liu, Y.; Han, F.; Xiao, Y.; Gao, Y. Preferential and rapid degradation of raw rice starch by an α-amylase of glycoside hydrolase subfamily GH13_37. Appl. Microbiol. Biotechnol. 2012, 94, 1577–1584. [Google Scholar]
- Gangadharan, D.; Nampoothiri, K.M.; Sivaramakrishnan, S.; Pandey, A. Biochemical characterization of raw-starch-digesting alpha amylase purified from Bacillus amyloliquefaciens. Appl. Biochem. Biotechnol. 2009, 158, 653–662. [Google Scholar]
- Liu, X.D.; Xu, Y. A novel raw starch digesting α-amylase from a newly isolated Bacillus sp. YX-1: Purification and characterization. Bioresour. Technol. 2008, 99, 4315–4320. [Google Scholar]
- Bergthaller, W.; Hollmann, J. Starch. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier Science: Amsterdam, The Netherlands, 2014; pp. 579–612. [Google Scholar]
- Fardet, A.; Rock, E.; Rémésy, C. Is the in vitro antioxidant potential of whole-grain cereals and cereal products well reflected in vivo? J. Cereal Sci. 2008, 48, 258–276. [Google Scholar]
- Alrahmany, R.; Avis, T.J.; Tsopmo, A. Treatment of oat bran with carbohydrases increases soluble phenolic acid content and influences antioxidant and antimicrobial activities. Food Res. Int. 2013, 52, 568–574. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar]
- Laemmli, U. Most commonly used discontinuous buffer system for SDS electrophoresis. Nature 1970, 227, 680–686. [Google Scholar]
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Total Activity (U) | Total Protein (mg) | Specific Activity (U mg−1) | Recovery (%) | Purification Fold |
|---|---|---|---|---|---|
| Crude visceral extract | 3000.00 | 913.70 | 3.28 | 100.00 | 1.00 |
| Ultrafiltration (10 kDa) | 2550.00 | 165.00 | 15.45 | 85.00 | 4.71 |
| Sephadex G-100 | 1470.00 | 14.40 | 102.08 | 49.00 | 31.12 |
| Mono Q Sepharose | 834.50 | 0.60 | 1390.80 | 27.80 | 424.02 |
| Metal Ions (5 mM) | Relative Activity (%) |
|---|---|
| None | 100.00 |
| Ca2+ | 97.00 ± 1.40 |
| Fe2+ | 17.40 ± 0.84 |
| Mg2+ | 109.70 ± 0.67 |
| Ba2+ | 118.50 ± 2.12 |
| Hg2+ | 0 |
| Zn2+ | 0 |
| Mn2+ | 6.20 ± 0.42 |
| Co2+ | 18.75 ± 0.35 |
| Cu2+ | 8.40 ± 0.56 |
| Al2+ | 91.30 ± 0.98 |
| K+ | 95.50 ± 0.61 |
| Na+ | 100.00 |
| Chemical Reagents | Concentration | Remaining Activity (%) |
|---|---|---|
| None | --- | 100.00 |
| Surfactants | ||
| Tween 20 | 5% (v/v) | 71.13 ± 1.49 |
| Tween 80 | 5% (v/v) | 71.00 ± 0.42 |
| Triton X-100 | 5% (v/v) | 80.78 ± 2.52 |
| SDS | 1% (w/v) | 26.20 ± 1.54 |
| 2% (w/v) | 17.55 ± 1.48 | |
| Inhibitors | ||
| β-mercaptoethanol | 5 mM | 93.90 ± 2.68 |
| EDTA | 5 mM | 92.89 ± 0.84 |
| Substrates | Relative Activity (%) |
|---|---|
| Potato starch | 100.00 |
| Maize starch | 83.40 ± 1.55 |
| Wheat starch | 96.20 ± 1.05 |
| Amylose | 17.80 ± 2.25 |
| Amylopectin | 71.13 ± 0.74 |
| CMC | 0 |
| α-Cyclodextrin | 0 |
| Dextrin | 0 |
| Chitosan | 0 |
| Sample | α-Amylase Activity (Units) or Gallic Acid Concentration (µg mL−1) | DPPH Scavenging Activity (% of Inhibition) | Total Soluble Phenolic Compounds (TSPC) (Gallic Acid Equivalent in µg g−1 Oat) |
|---|---|---|---|
| Native oat (4%) | 0 | 38.40 ± 1.15 | 23.00 ± 2.50 |
| BCA-treated oat | 10.00 | 67.53 ± 2.05 | 75.00 ± 4.25 |
| Gallic acid * | — | 50.00 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maalej, H.; Maalej, A.; Affes, S.; Hmidet, N.; Nasri, M. A Novel Digestive α-Amylase from Blue Crab (Portunus segnis) Viscera: Purification, Biochemical Characterization and Application for the Improvement of Antioxidant Potential of Oat Flour. Int. J. Mol. Sci. 2021, 22, 1070. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031070
Maalej H, Maalej A, Affes S, Hmidet N, Nasri M. A Novel Digestive α-Amylase from Blue Crab (Portunus segnis) Viscera: Purification, Biochemical Characterization and Application for the Improvement of Antioxidant Potential of Oat Flour. International Journal of Molecular Sciences. 2021; 22(3):1070. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031070
Chicago/Turabian StyleMaalej, Hana, Amina Maalej, Sawsan Affes, Noomen Hmidet, and Moncef Nasri. 2021. "A Novel Digestive α-Amylase from Blue Crab (Portunus segnis) Viscera: Purification, Biochemical Characterization and Application for the Improvement of Antioxidant Potential of Oat Flour" International Journal of Molecular Sciences 22, no. 3: 1070. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031070