
Pulmonary Vascular Complications in Hereditary Hemorrhagic Telangiectasia and the Underlying Pathophysiology

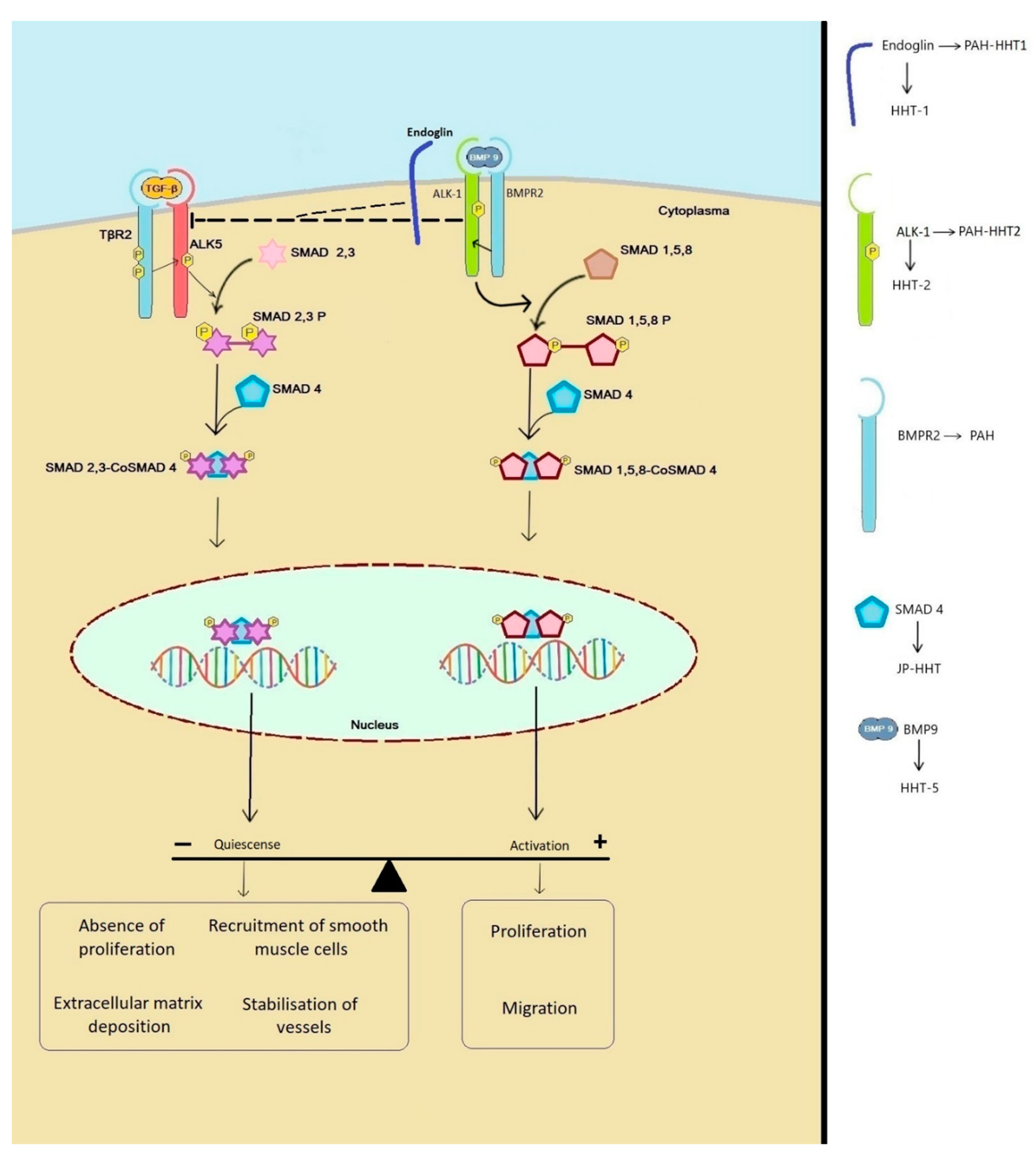
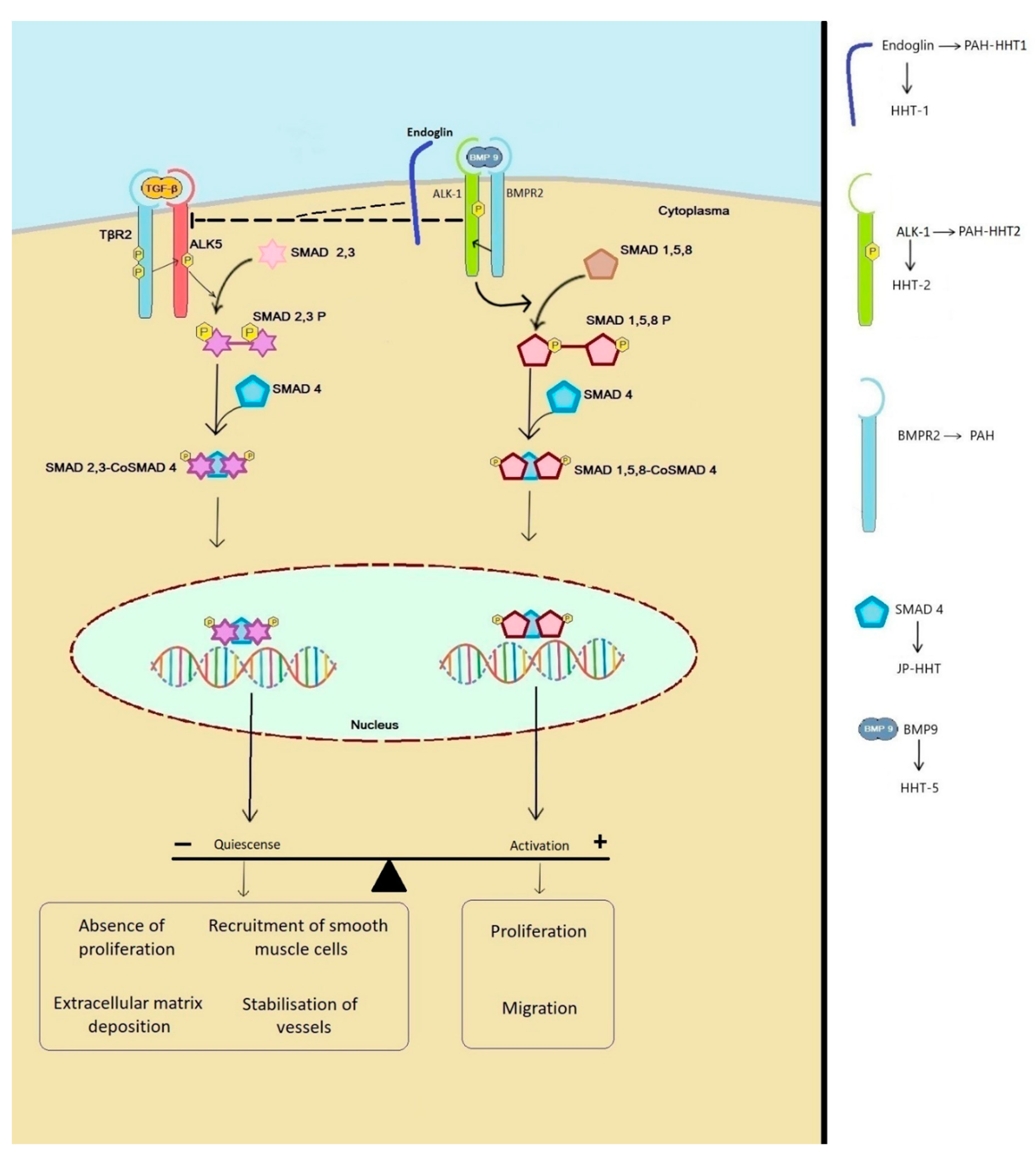{kind=link}
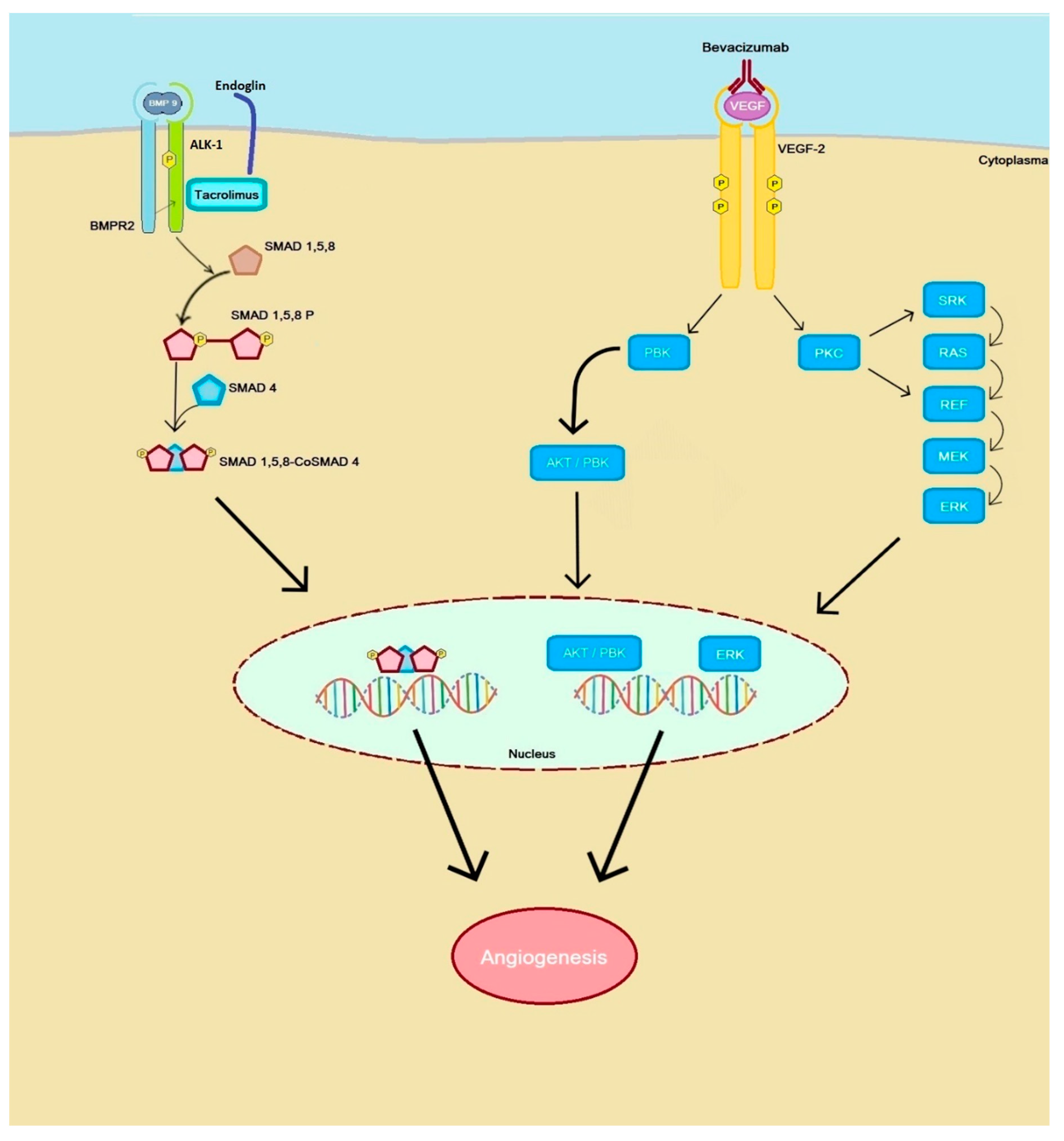
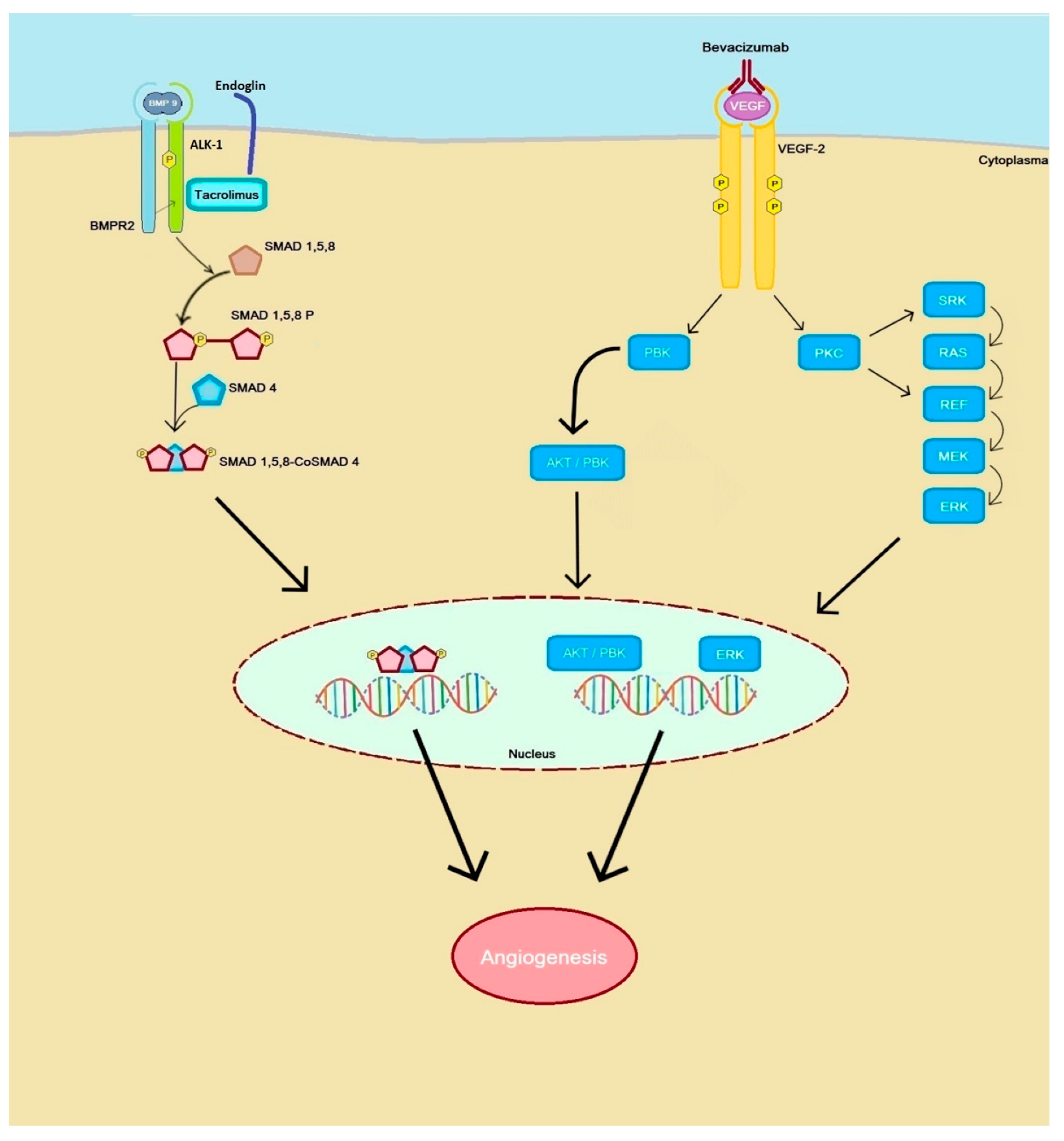{kind=link}
Abstract
:1. Hereditary Hemorrhagic Telangiectasia
- Frequent and recurrent epistaxis, which may be mild to severe
- Multiple telangiectases on characteristic sites: lips, oral cavity, fingers, and nose
- AVMs or telangiectases in one or more of the internal organs (lung, brain, liver, intestines, stomach, and spinal cord)
- A 1st-degree relative with HHT
2. Pulmonary Arteriovenous Malformations in HHT
3. Pulmonary Hypertension Caused by HHT
- Group 1—Pulmonary arterial hypertension (PAH)
- Group 2—PH caused by left heart disease
- Group 3—PH caused by lung diseases and hypoxia
- Group 4—Chronic thrombo-embolic PH and other pulmonary arterial obstructions
- Group 5—PH with unclear or multifactorial mechanisms
4. Treatment and Follow-Up of Pulmonary Vascular Disease in HHT
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dakeishi, M.; Shioya, T.; Wada, Y.; Shindo, T.; Otaka, K.; Manabe, M.; Nozaki, J.-I.; Inoue, S.; Koizumi, A. Genetic epidemiology of hereditary hemorrhagic telangiectasia in a local community in the northern part of Japan. Hum. Mutat. 2002, 19, 140–148. [Google Scholar] [CrossRef]
- Faughnan, M.E.; Mager, J.J.; Hetts, S.W.; Palda, V.A.; Lang-Robertson, K.; Buscarini, E.; Deslandres, E.; Kasthuri, R.S.; Lausman, A.; Poetker, D.; et al. Second International Guidelines for the Diagnosis and Management of Hereditary Hemorrhagic Telangiectasia. Ann. Intern. Med. 2020, 173, 989–1001. [Google Scholar] [CrossRef]
- McDonald, J.; Ewooderchak-Donahue, W.; Webb, C.; Van Sant, E.K.; Stevenson, D.A.; Ebayrak-Toydemir, P. Hereditary hemorrhagic telangiectasia: Genetics and molecular diagnostics in a new era. Front. Genet. 2015, 6, 1. [Google Scholar] [CrossRef]
- Johnson, D.W.; Berg, J.N.A.; Baldwin, M.; Gallione, C.J.; Marondel, I.; Yoon, S.-J.; Stenzel, T.T.; Speer, M.C.A.; Pericakvance, M.; Diamond, A.G.; et al. Mutations in the activin receptor-like kinase 1 gene in hereditary haemorrhagic telangiectasia type 2. Nat. Genet. 1996, 13, 189–195. [Google Scholar] [CrossRef]
- McAllister, K.A.; Grogg, K.; Johnson, D.; Gallione, C.J.; Baldwin, P.E.; Jackson, C.; Helmbold, E.; Markel, D.; McKinnon, W.; Murrell, J.R.; et al. Endoglin, a TGF-β binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. Nat. Genet. 1994, 8, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Gallione, C.J.; Repetto, G.M.; Legius, E.; Rustgi, A.K.; Schelley, S.L.; Tejpar, S.; Mitchell, G.; Drouin, E.; Westermann, C.J.A.; Marchuk, D. A combined syndrome of juvenile polyposis and hereditary haemorrhagic telangiectasia associated with mutations in MADH4 (SMAD4). Lancet 2004, 363, 852–859. [Google Scholar] [CrossRef]
- Goumans, M.-J.; Lebrin, F.; Valdimarsdottir, G. Controlling the Angiogenic SwitchA Balance between Two Distinct TGF-b Receptor Signaling Pathways. Trends Cardiovasc. Med. 2003, 13, 301–307. [Google Scholar] [CrossRef]
- Vorselaars, V.M.; Velthuis, S.; Snijder, R.J.; Vos, J.A.; Mager, J.J.; Post, M.C. Pulmonary hypertension in hereditary haemorrhagic telangiectasia. World J. Cardiol. 2015, 7, 230–237. [Google Scholar] [CrossRef]
- Vorselaars, V.M.M.; Hosman, A.E.; Westermann, C.J.J.; Snijder, R.J.; Mager, J.J.; Goumans, M.-J.; Post, M.C. Pulmonary Arterial Hypertension and Hereditary Haemorrhagic Telangiectasia. Int. J. Mol. Sci. 2018, 19, 3203. [Google Scholar] [CrossRef] [Green Version]
- Pierucci, P.; Lenato, G.M.; Suppressa, P.; Lastella, P.; Triggiani, V.; Valerio, R.; Comelli, M.; Salvante, D.; Stella, A.; Resta, N.; et al. A long diagnostic delay in patients with Hereditary Haemorrhagic Telangiectasia: A questionnaire-based retrospective study. Orphanet J. Rare Dis. 2012, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Shovlin, C.L.; Guttmacher, A.E.; Buscarini, E.; Faughnan, M.E.; Hyland, R.H.; Westermann, C.J.J.; Kjeldsen, A.D.; Plauchu, H. Diagnostic criteria for hereditary hemorrhagic telangiectasia (Rendu-Osler-Weber syndrome). Am. J. Med. Genet. 2000, 91, 66–67. [Google Scholar] [CrossRef]
- Galiè, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Hoeper, M. 2015 ESC/ERS Guidelines for the diagnosis and treatment of pulmonary hypertension-web addenda. Eur. Respir. J. 2015, 1183. [Google Scholar]
- Dingenouts, C.K.E.; Goumans, M.-J.; Bakker, W. Mononuclear cells and vascular repair in HHT. Front. Genet. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Llorente, L.; Gallardo-Vara, E.; Rossi, E.; Smadja, D.M.; Botella, L.M.; Bernabeu, C. Endoglin and alk1 as therapeutic targets for hereditary hemorrhagic telangiectasia. Expert Opin. Ther. Targets 2017, 21, 933–947. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.G.E.; Begbie, M.; Wallace, G.M.F.; Shovlin, C.L. A new locus for hereditary haemorrhagic telangiectasia (HHT3) maps to chromosome 5. J. Med. Genet. 2005, 42, 577–582. [Google Scholar] [CrossRef]
- Bayrak-Toydemir, P.; McDonald, J.; Akarsu, N.; Toydemir, R.M.; Calderon, F.; Tuncali, T.; Tang, W.; Miller, F.; Mao, R. A fourth locus for hereditary hemorrhagic telangiectasia maps to chromosome 7. Am. J. Med. Genet. Part A 2006, 140, 2155–2162. [Google Scholar] [CrossRef]
- Botella, L.-M.; Albiã, A.V.; Ojeda-Fernández, L.; Recio-Poveda, L.; Bernabã, U.C.; Albinana, V.; Bernabeu, C. Research on potential biomarkers in hereditary hemorrhagic telangiectasia. Front. Genet. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Wooderchak-Donahue, W.L.; McDonald, J.; O’Fallon, B.; Upton, P.D.; Li, W.; Roman, B.L.; Young, S.; Plant, P.; Fülöp, G.T.; Langa, C.; et al. BMP9 Mutations Cause a Vascular-Anomaly Syndrome with Phenotypic Overlap with Hereditary Hemorrhagic Telangiectasia. Am. J. Hum. Genet. 2013, 93, 530–537. [Google Scholar] [CrossRef] [Green Version]
- Wan, M.; Li, C.; Zhen, G.; Jiao, K.; He, W.; Jia, X.; Wang, W.; Shi, C.; Xing, Q.; Chen, Y.-F.; et al. Injury-Activated Transforming Growth Factor β Controls Mobilization of Mesenchymal Stem Cells for Tissue Remodeling. Stem Cells 2012, 30, 2498–2511. [Google Scholar] [CrossRef] [Green Version]
- Pepper, M.S.; Vassalli, J.-D.; Orci, L.; Montesano, R. Biphasic Effect of Transforming Growth Factor-β1 on in Vitro Angiogenesis. Exp. Cell Res. 1993, 204, 356–363. [Google Scholar] [CrossRef]
- Pepper, M.S. Transforming growth factor-beta: Vasculogenesis, angiogenesis, and vessel wall integrity. Cytokine Growth Factor Rev. 1997, 8, 21–43. [Google Scholar] [CrossRef]
- Rudini, N.; Felici, A.; Giampietro, C.; Lampugnani, M.G.; Corada, M.; Swirsding, K.; Garre, M.; Liebner, S.; Letarte, M.; Dijke, P.T.; et al. VE-cadherin is a critical endothelial regulator of TGF-β signalling. EMBO J. 2008, 27, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Goumans, M.; Valdimarsdottir, G.; Itoh, S.; Rosendahl, A.; Sideras, P.; Dijke, P.T. Balancing the activation state of the endothelium via two distinct TGF-beta type I receptors. EMBO J. 2002, 21, 1743–1753. [Google Scholar] [CrossRef]
- Oh, S.P.; Seki, T.; Goss, K.A.; Imamura, T.; Yi, Y.; Donahoe, P.K.; Li, L.; Miyazono, K.; Dijke, P.T.; Kim, S.; et al. Activin receptor-like kinase 1 modulates transforming growth factor-beta 1 signaling in the regulation of angiogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 2626–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlova, V.V.; Liu, Z.; Goumans, M.J.; Dijke, P.T. Controlling angiogenesis by two unique TGF-β type I receptor signaling pathways. Histol. Histopathol. 2011, 26. [Google Scholar] [CrossRef]
- Shao, E.S.; Lin, L.; Yao, Y.; Boström, K.I. Expression of vascular endothelial growth factor is coordinately regulated by the activin-like kinase receptors 1 and 5 in endothelial cells. Blood 2009, 114, 2197–2206. [Google Scholar] [CrossRef] [Green Version]
- Goumans, M.-J.; Valdimarsdottir, G.; Itoh, S.; Lebrin, F.; Larsson, J.; Mummery, C.; Karlsson, S.; Dijke, P.T. Activin Receptor-like Kinase (ALK)1 Is an Antagonistic Mediator of Lateral TGFβ/ALK5 Signaling. Mol. Cell 2003, 12, 817–828. [Google Scholar] [CrossRef]
- Lamouille, S.; Mallet, C.; Feige, J.-J.; Bailly, S. Activin receptor-like kinase 1 is implicated in the maturation phase of angiogenesis. Blood 2002, 100, 4495–4501. [Google Scholar] [CrossRef]
- Lebrin, F.; Mummery, C.L. Endoglin-Mediated Vascular Remodeling: Mechanisms Underlying Hereditary Hemorrhagic Telangiectasia. Trends Cardiovasc. Med. 2008, 18, 25–32. [Google Scholar] [CrossRef]
- Lebrin, F.; Goumans, M.-J.; Jonker, L.; Carvalho, R.L.C.; Valdimarsdottir, G.; Thorikay, M.; Mummery, C.; Arthur, H.M.; Dijke, P.T. Endoglin promotes endothelial cell proliferation and TGF-β/ALK1 signal transduction. EMBO J. 2004, 23, 4018–4028. [Google Scholar] [CrossRef] [Green Version]
- Scharpfenecker, M.; Van Dinther, M.; Liu, Z.; Van Bezooijen, R.L.; Zhao, Q.; Pukac, L.; Löwik, C.W.G.M.; Dijke, P.T. BMP-9 signals via ALK1 and inhibits bFGF-induced endothelial cell proliferation and VEGF-stimulated angiogenesis. J. Cell Sci. 2007, 120, 964–972. [Google Scholar] [CrossRef] [Green Version]
- Scherner, O.; Meurer, S.K.; Tihaa, L.; Gressner, A.M.; Weiskirchen, R. Endoglin Differentially Modulates Antagonistic Transforming Growth Factor-β1 and BMP-7 Signaling. J. Biol. Chem. 2007, 282, 13934–13943. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Elsner, T.; Botella, L.M.; Velasco, B.; Langa, C.; Bernabéu, C. Endoglin Expression is Regulated by Transcriptional Cooperation between the Hypoxia and Transforming Growth Factor-β Pathways. J. Biol. Chem. 2002, 277, 43799–43808. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Ohga, N.; Morishita, Y.; Hida, K.; Miyazono, K.; Watabe, T. BMP-9 induces proliferation of multiple types of endothelial cells in vitro and in vivo. J. Cell Sci. 2010, 123, 1684–1692. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Salmon, R.M.; Jiang, H.; Morrell, N.W. Regulation of the ALK1 ligands, BMP9 and BMP10. Biochem. Soc. Trans. 2016, 44, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Mostafa, S.; Pakvasa, M.; Coalson, E.; Zhu, A.; Alverdy, A.; Castillo, H.; Fan, J.; Li, A.; Feng, Y.; Wu, D.; et al. The wonders of BMP9: From mesenchymal stem cell differentiation, angiogenesis, neurogenesis, tumorigenesis and metabolism to regenerative medicine. Genes Dis. 2019, 6, 201–223. [Google Scholar] [CrossRef]
- Jerkic, M.; Rodríguez-Barbero, A.; Prieto, M.; Toporsian, M.; Pericacho, M.; Rivas-Elena, J.V.; Obreo, J.; Wang, A.; Pérez-Barriocanal, F.; Arévalo, M.; et al. Reduced angiogenic responses in adult endoglin heterozygous mice. Cardiovasc. Res. 2006, 69, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Düwel, A.; Eleno, N.; Jerkic, M.; Arévalo, M.; Bolaños, J.P.; Bernabeu, C.; López-Novoa, J.M. Reduced Tumor Growth and Angiogenesis in Endoglin-Haploinsufficient Mice. Tumor Biol. 2007, 28, 1–8. [Google Scholar] [CrossRef]
- Liu, Z.; Lebrin, F.; Maring, J.A.; Driesche, S.V.D.; Van Der Brink, S.; Van Dinther, M.; Thorikay, M.; Martin, S.; Kobayashi, K.; Hawinkels, L.J.A.C.; et al. Endoglin is Dispensable for Vasculogenesis, but Required for Vascular Endothelial Growth Factor-Induced Angiogenesis. PLoS ONE 2014, 9, e86273. [Google Scholar] [CrossRef] [Green Version]
- Roelen, B.A.J.; Van Rooijen, M.A.; Mummery, C.L. Expression of ALK-1, a type 1 serine/threonine kinase receptor, coincides with sites of vasculogenesis and anglogenesis in early mouse development. Dev. Dyn. 1997, 209. [Google Scholar] [CrossRef]
- Seki, T.; Hong, K.-H.; Oh, S.P. Nonoverlapping expression patterns of ALK1 and ALK5 reveal distinct roles of each receptor in vascular development. Lab. Investig. 2005, 86, 116–129. [Google Scholar] [CrossRef]
- Torsney, E.; Charlton, R.; Parums, D.; Collis, M.; Arthur, H.M. Inducible expression of human endoglin during inflammation and wound healing in vivo. Inflamm. Res. 2002, 51, 464–470. [Google Scholar] [CrossRef]
- Seki, T.; Yun, J.; Oh, S.P. Arterial Endothelium-Specific Activin Receptor-Like Kinase 1 Expression Suggests its Role in Arterialization and Vascular Remodeling. Circ. Res. 2003, 93, 682–689. [Google Scholar] [CrossRef] [Green Version]
- Tellapuri, S.; Park, H.S.; Kalva, S.P. Pulmonary arteriovenous malformations. Int. J. Cardiovasc. Imaging 2018, 35, 1421–1428. [Google Scholar] [CrossRef]
- Sadick, H.; Riedel, F.; Naim, R.; Goessler, U.; Hormann, K.; Hafner, M.; Lux, A. Patients with hereditary hemorrhagic telangiectasia have increased plasma levels of vascular endothelial growth factor and transforming growth factor-β1 as well as high ALK1 tissue expression. Haematologica 2005, 90, 818–828. [Google Scholar]
- Sadick, H.; Hage, J.; Bran, G.; Hoermann, K.; Goessler, U.; Riedel, F.; Bugert, P. Does the genotype of HHT patients with mutations of the ENG and ACVRL1 gene correlate to different expression levels of the angiogenic factor VEGF? Int. J. Mol. Med. 1998, 22, 575–580. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Choe, S.-W.; Kim, Y.H.; Acharya, A.P.; Keselowsky, B.G.; Sorg, B.S.; Lee, Y.-J.; Oh, S.P. VEGF neutralization can prevent and normalize arteriovenous malformations in an animal model for hereditary hemorrhagic telangiectasia 2. Angiogenesis 2014, 17, 823–830. [Google Scholar] [CrossRef]
- Abdalla, S.A. Hereditary haemorrhagic telangiectasia: Current views on genetics and mechanisms of disease. J. Med. Genet. 2005, 43, 97–110. [Google Scholar] [CrossRef]
- Tual-Chalot, S.; Oh, S.P.; Arthur, H.M. Mouse models of hereditary hemorrhagic telangiectasia: Recent advances and future challenges. Front. Genet. 2015, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, C.; Bayrak-Toydemir, P.; McDonald, J.; Letarte, M. Potential Second-Hits in Hereditary Hemorrhagic Telangiectasia. J. Clin. Med. 2020, 9, 3571. [Google Scholar] [CrossRef]
- Dijke, P.T.; Goumans, M.-J.; Pardali, E. Endoglin in angiogenesis and vascular diseases. Angiogenesis 2008, 11, 79–89. [Google Scholar] [CrossRef]
- De Vinuesa, A.G.; Bocci, M.; Pietras, K.; Dijke, P.T. Targeting tumour vasculature by inhibiting activin receptor-like kinase (ALK)1 function. Biochem. Soc. Trans. 2016, 44, 1142–1149. [Google Scholar] [CrossRef] [Green Version]
- López-Novoa, J.M.; Bernabeu, C. The physiological role of endoglin in the cardiovascular system. Am. J. Physiol. Circ. Physiol. 2010, 299, H959–H974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, E.J.; Su, H.; Shen, F.; Degos, V.; Amend, G.; Jun, K.; Young, W.L. Bevacizumab attenuates VEGF-induced angiogenesis and vascular malformations in the adult mouse brain. Stroke 2012, 43, 1925–1930. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, M.; Allinson, K.R.; Zhai, Z.; Oakenfull, R.; Ghandi, P.; Adams, R.H.; Fruttiger, M.; Arthur, H.M. Pathogenesis of Arteriovenous Malformations in the Absence of Endoglin. Circ. Res. 2010, 106, 1425–1433. [Google Scholar] [CrossRef] [Green Version]
- Park, S.O.; Wankhede, M.; Lee, Y.J.; Choi, E.-J.; Fliess, N.; Choe, S.-W.; Oh, S.-H.; Walter, G.; Raizada, M.K.; Sorg, B.S. Real-time imaging of de novo arteriovenous malformation in a mouse model of hereditary hemorrhagic telangiectasia. J. Clin. Investig. 2009, 119, 3487–3496. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-J.; Walker, E.J.; Shen, F.; Oh, S.P.; Arthur, H.M.; Young, W.L.; Su, H. Minimal Homozygous Endothelial Deletion of Eng with VEGF Stimulation is Sufficient to Cause Cerebrovascular Dysplasia in the Adult Mouse. Cerebrovasc. Dis. 2012, 33, 540–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottin, V.; Plauchu, H.; Bayle, J.-Y.; Barthelet, M.; Revel, D.; Cordier, J.-F. Pulmonary Arteriovenous Malformations in Patients with Hereditary Hemorrhagic Telangiectasia. Am. J. Respir. Crit. Care Med. 2004, 169, 994–1000. [Google Scholar] [CrossRef] [Green Version]
- Shovlin, C.; Jackson, J.; Wilmshurst, P. Pulmonary arteriovenous malformations and other pulmonary aspects of HHT. In Orphan Lung Diseases; European Respiratory Society (ERS): Lausanne, Switzerland, 2011; pp. 218–245. [Google Scholar]
- Shovlin, C.L.; Condliffe, R.; Donaldson, J.W.; Kiely, D.G.; Wort, S.J. British Thoracic Society Clinical Statement on Pulmonary Arteriovenous Malformations. Thorax 2017, 72, 1154–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, W.E.; Jackson, J.; Layton, D.M.; Shovlin, C.L. Hypoxaemia, sport and polycythaemia: A case from Imperial College London. Thorax 2015, 70, 601–603. [Google Scholar] [CrossRef] [Green Version]
- Santhirapala, V.; Williams, L.C.; Tighe, H.C.; Jackson, J.E.; Shovlin, C.L. Arterial Oxygen Content is Precisely Maintained by Graded Erythrocytotic Responses in Settings of High/Normal Serum Iron Levels and Predicts Exercise Capacity: An Observational Study of Hypoxaemic Patients with Pulmonary Arteriovenous Malformations. PLoS ONE 2014, 9, e90777. [Google Scholar] [CrossRef] [Green Version]
- Van Gent, M.W.F.; Post, M.C.; Snijder, R.J.; Westermann, C.J.J.; Plokker, H.W.M.; Mager, J.J. Real Prevalence of Pulmonary Right-to-Left Shunt According to Genotype in Patients with Hereditary Hemorrhagic Telangiectasia. Chest 2010, 138, 833–839. [Google Scholar] [CrossRef]
- Cottin, V.; Chinet, T.; Lavolé, A.; Corre, R.; Marchand, E.; Reynaud-Gaubert, M.; Plauchu, H.; Cordier, J.-F. Pulmonary Arteriovenous Malformations in Hereditary Hemorrhagic Telangiectasia. Medicine 2007, 86, 1–17. [Google Scholar] [CrossRef]
- Shovlin, C.L.; Sodhi, V.; McCarthy, A.; Lasjaunias, P.E.; Jackson, J.; Sheppard, M.N. Estimates of maternal risks of pregnancy for women with hereditary haemorrhagic telangiectasia (Osler-Weber-Rendu syndrome): Suggested approach for obstetric services. BJOG Int. J. Obstet. Gynaecol. 2008, 115, 1108–1115. [Google Scholar] [CrossRef] [Green Version]
- Gossage, J.R.; Kanj, G. Pulmonary Arteriovenous Malformations. Am. J. Respir. Crit. Care Med. 1998, 158, 643–661. [Google Scholar] [CrossRef]
- Hosman, A.E.; De Gussem, E.M.; Balemans, W.A.F.; Gauthier, A.; Westermann, C.J.J.; Snijder, R.J.; Post, M.C.; Mager, J.J. Screening children for pulmonary arteriovenous malformations: Evaluation of 18 years of experience. Pediatr. Pulmonol. 2017, 52, 1206–1211. [Google Scholar] [CrossRef]
- White, R.I.E.; Mitchell, S.; Barth, K.H.; Kaufman, S.L.; Kadir, S.; Chang, R.; Terry, P.B. Angioarchitecture of pulmonary arteriovenous malformations: An important consideration before embolotherapy. Am. J. Roentgenol. 1983, 140, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Pierucci, P.; Murphy, J.; Henderson, K.J.; Chyun, D.A.; White, R.I. New Definition and Natural History of Patients with Diffuse Pulmonary Arteriovenous Malformations. Chest 2008, 133, 653–661. [Google Scholar] [CrossRef]
- Kjeldsen, A.D.; Oxhøj, H.; Andersen, P.E.; Elle, B.; Jacobsen, J.P.; Vase, P. Pulmonary Arteriovenous Malformations. Chest 1999, 116, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Circo, S.; Gossage, J.R. Pulmonary vascular complications of hereditary haemorrhagic telangiectasia. Curr. Opin. Pulm. Med. 2014, 20, 421–428. [Google Scholar] [CrossRef]
- Velthuis, S.; Vorselaars, V.M.M.; Van Gent, M.W.F.; Westermann, C.J.J.; Snijder, R.J.; Mager, J.J.; Post, M.C. Role of Transthoracic Contrast Echocardiography in the Clinical Diagnosis of Hereditary Hemorrhagic Telangiectasia. Chest 2013, 144, 1876–1882. [Google Scholar] [CrossRef] [PubMed]
- Velthuis, S.; Buscarini, E.; Mager, J.J.; Vorselaars, V.M.; Van Gent, M.W.; Gazzaniga, P.; Manfredi, G.; Danesino, C.; Diederik, A.L.; Vos, J.A.; et al. Predicting the size of pulmonary arteriovenous malformations on chest computed tomography: A role for transthoracic contrast echocardiography. Eur. Respir. J. 2014, 44, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Post, M.C.; Letteboer, T.G.W.; Mager, J.J.; Plokker, T.H.; Kelder, J.C.; Westermann, C.J.J. A Pulmonary Right-to-Left Shunt in Patients with Hereditary Hemorrhagic Telangiectasia is Associated with an Increased Prevalence of Migraine. Chest 2005, 128, 2485–2489. [Google Scholar] [CrossRef] [Green Version]
- Lesca, G.; Olivieri, C.; Burnichon, N.; Pagella, F.; Carette, M.-F.; Gilbert-Dussardier, B.; Goizet, C.; Roume, J.; Rabilloud, M.; Saurin, J.-C.; et al. Genotype-phenotype correlations in hereditary hemorrhagic telangiectasia: Data from the French-Italian HHT network. Genet. Med. 2007, 9, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Eyries, M.; Montani, D.; Nadaud, S.; Girerd, B.; Levy, M.; Bourdin, A.; Trésorier, R.; Chaouat, A.; Cottin, V.; Sanfiorenzo, C.; et al. Widening the landscape of heritable pulmonary hypertension mutations in paediatric and adult cases. Eur. Respir. J. 2019, 53, 1801371. [Google Scholar] [CrossRef] [Green Version]
- Cogan, J.D.; Vnencak-Jones, C.L.A.; Phillips, J.; Lane, K.B.A.; Wheeler, L.; Robbins, I.M.; Garrison, G.; Hedges, L.K.E.; Loyd, J. Gross BMPR2 gene rearrangements constitute a new cause for primary pulmonary hypertension. Genet. Med. 2005, 7, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Cogan, J.D.; Pauciulo, M.W.; Batchman, A.P.; Prince, M.A.; Robbins, I.M.; Hedges, L.K.; Stanton, K.C.; Wheeler, L.A.; Phillips, J.A.; Loyd, J.E.; et al. High Frequency ofBMPR2Exonic Deletions/Duplications in Familial Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2006, 174, 590–598. [Google Scholar] [CrossRef]
- Aldred, M.A.; Vijayakrishnan, J.; James, V.; Soubrier, F.; Gómez-Sánchez, M.A.; Mårtensson, G.; Galié, N.; Manes, A.; Corris, P.; Simonneau, G.; et al. BMPR2 gene rearrangements account for a significant proportion of mutations in familial and idiopathic pulmonary arterial hypertension. Hum. Mutat. 2006, 27, 212–213. [Google Scholar] [CrossRef]
- Thomson, J.R.; Machado, R.D.; Pauciulo, M.W.; Morgan, N.V.; Humbert, M.; Elliott, G.C.; Ward, K.; Yacoub, M.; Mikhail, G.; Rogers, P.; et al. Sporadic primary pulmonary hypertension is associated with germline mutations of the gene encoding BMPR-II, a receptor member of the TGF-beta family. J. Med. Genet. 2000, 37, 741–745. [Google Scholar] [CrossRef]
- Atkinson, C.; Stewart, S.; Upton, P.D.; Machado, R.D.; Thomson, J.R.; Trembath, R.C.; Morrell, N.W. Primary Pulmonary Hypertension is Associated with Reduced Pulmonary Vascular Expression of Type II Bone Morphogenetic Protein Receptor. Circulation 2002, 105, 1672–1678. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Mallet, C.; Mazerbourg, S.; Feige, J.-J.; Bailly, S. Identification of BMP9 and BMP10 as functional activators of the orphan activin receptor-like kinase 1 (ALK1) in endothelial cells. Blood 2006, 109, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Ormiston, M.L.; Yang, X.; Southwood, M.; Gräf, S.; Machado, R.D.; Mueller, M.; Kinzel, B.; Yung, L.M.; Wilkinson, J.M.; et al. Selective enhancement of endothelial BMPR-II with BMP9 reverses pulmonary arterial hypertension. Nat. Med. 2015, 21, 777–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girerd, B.; Montani, D.; Coulet, F.; Sztrymf, B.; Yaici, A.; Jaïs, X.; Tregouet, D.; Reis, A.; Drouin-Garraud, V.; Fraisse, A.; et al. Clinical Outcomes of Pulmonary Arterial Hypertension in Patients Carrying anACVRL1 (ALK1) Mutation. Am. J. Respir. Crit. Care Med. 2010, 181, 851–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenzweig, E.B.; Morse, J.H.; Knowles, J.A.; Chada, K.K.; Khan, A.M.; Roberts, K.E.; McElroy, J.J.; Juskiw, N.K.; Mallory, N.C.; Rich, S.; et al. Clinical Implications of Determining BMPR2 Mutation Status in a Large Cohort of Children and Adults with Pulmonary Arterial Hypertension. J. Heart Lung Transplant. 2008, 27, 668–674. [Google Scholar] [CrossRef]
- Elliott, C.G.; Glissmeyer, E.W.; Havlena, G.T.; Carlquist, J.; McKinney, J.T.; Rich, S.; McGoon, M.D.; Scholand, M.B.; Kim, M.; Jensen, R.L.; et al. Relationship of BMPR2 Mutations to Vasoreactivity in Pulmonary Arterial Hypertension. Circulation 2006, 113, 2509–2515. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xiong, C.-M.; Gu, Q.; Wang, X.-T.; Cheng, X.-L.; Huang, L.; Yang, T.; Luo, Q.; Zhao, Z.-H.; Ni, X.-H.; et al. The clinical characteristics and long-term prognosis of pulmonary arterial hypertension associated with hereditary hemorrhagic telangiectasia. Pulm. Circ. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Buscarini, E.; Plauchu, H.; Tsao, G.G.; White, R.I.; Sabbà, C.; Miller, F.; Saurin, J.C.; Pelage, J.P.; Lesca, G.; Marion, M.J.; et al. Liver involvement in hereditary hemorrhagic telangiectasia: Consensus recommendations. Liver Int. 2006, 26, 1040–1046. [Google Scholar] [CrossRef]
- Remy-Jardin, M.; Wattinne, L.; Remy, J. Transcatheter occlusion of pulmonary arterial circulation and collateral supply: Failures, incidents and complications. Radiology 1991, 180, 699–705. [Google Scholar] [CrossRef]
- Shovlin, C.L.; Buscarini, E.; Hughes, J.M.B.; Allison, D.J.E.; Jackson, J. Long-term outcomes of patients with pulmonary arteriovenous malformations considered for lung transplantation, compared with similarly hypoxaemic cohorts. BMJ Open Respir. Res. 2017, 4, e000198. [Google Scholar] [CrossRef]
- Cartin-Ceba, R.; Swanson, K.L.; Krowka, M.J. Pulmonary Arteriovenous Malformations. Chest 2013, 144, 1033–1044. [Google Scholar] [CrossRef]
- Remy-Jardin, M.; Dumont, P.; Brillet, P.-Y.; Dupuis, P.; Duhamel, A.; Remy, J. Pulmonary Arteriovenous Malformations Treated with Embolotherapy: Helical CT Evaluation of Long-term Effectiveness after 2-21-Year Follow-up 1. Radiology 2006, 239, 576–585. [Google Scholar] [CrossRef]
- Robert, F.; Desroches-Castan, A.; Bailly, S.; Dupuis-Girod, S.; Feige, J.-J. Future treatments for hereditary hemorrhagic telangiectasia. Orphanet J. Rare Dis. 2020, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ospina, E.F.; Echeverri, A.; Osorio, I.P.; Jaimes, L.; Gutierrez, J.; Tobón, G.J. Bevacizumab as a treatment for hereditary hemorrhagic telangiectasia in children: A case report. Colomb. Med. 2017, 48, 88–93. [Google Scholar] [CrossRef]
- Roy, H.; Bhardwaj, S.; Ylä-Herttuala, S. Biology of vascular endothelial growth factors. FEBS Lett. 2006, 580, 2879–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirulli, A.; Liso, A.; D’Ovidio, F.; Mestice, A.; Pasculli, G.; Gallitelli, M.; Rizzi, R.; Specchia, G.; Sabbà, C. Vascular Endothelial Growth Factor Serum Levels are Elevated in Patients with Hereditary Hemorrhagic Telangiectasia. Acta Haematol. 2003, 110, 29–32. [Google Scholar] [CrossRef]
- Albiñana, V.; Cuesta, A.M.; De Rojas, P.I.; Gallardo-Vara, E.; Recio-Poveda, L.; Bernabéu, C.; Botella, L.M. Review of Pharmacological Strategies with Repurposed Drugs for Hereditary Hemorrhagic Telangiectasia Related Bleeding. J. Clin. Med. 2020, 9, 1766. [Google Scholar] [CrossRef]
- Spiekerkoetter, A.; Sung, Y.K.; Sudheendra, D.; Bill, M.; Aldred, M.A.; Van De Veerdonk, M.C.; Noordegraaf, A.V.; Long-Boyle, J.; Dash, R.; Yang, P.C.; et al. Low-Dose FK506 (Tacrolimus) in End-Stage Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2015, 192, 254–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiekerkoetter, E.; Tian, X.; Cai, J.; Hopper, R.K.; Sudheendra, D.; Li, C.G.; El-Bizri, N.; Sawada, H.; Haghighat, R.; Chan, R.; et al. FK506 activates BMPR2, rescues endothelial dysfunction and reverses pulmonary hypertension. J. Clin. Investig. 2013, 123, 3600–3613. [Google Scholar] [CrossRef] [Green Version]
- Albiñana, V.; Sanz-Rodríguez, F.; Recio-Poveda, L.; Bernabeu, C.; Botella, L.M. Immunosuppressor FK506 Increases Endoglin and Activin Receptor-Like Kinase 1 Expression and Modulates Transforming Growth Factor-β1 Signaling in Endothelial Cells. Mol. Pharmacol. 2011, 79, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Albiñana, V.; Velasco, L.; Zarrabeitia, R.; Botella, M. Tacrolimus as a therapeutic drug in hereditary hemorrhagic telangiectasia (HHT). In Tacrolimus: Effectiveness, Safety and Drug Interactions; Nova Biomedical: Milan, Italy, 2013. [Google Scholar]
- Ruiz, S.; Chandakkar, P.; Zhao, H.; Papoin, J.; Chatterjee, P.K.; Christen, E.; Metz, C.N.; Blanc, L.; Campagne, F.; Marambaud, P. Tacrolimus rescues the signaling and gene expression signature of endothelial ALK1 loss-of-function and improves HHT vascular pathology. Hum. Mol. Genet. 2017, 26, 4786–4798. [Google Scholar] [CrossRef]
- Spiekerkoetter, E.; Sung, Y.K.; Sudheendra, D.; Scott, V.; Del Rosario, P.; Bill, M.; Haddad, F.; Long-Boyle, J.; Hedlin, H.; Zamanian, R.T. Randomised placebo-controlled safety and tolerability trial of FK506 (tacrolimus) for pulmonary arterial hypertension. Eur. Respir. J. 2017, 50, 1602449. [Google Scholar] [CrossRef] [Green Version]
- Sommer, N.; Droege, F.; Gamen, K.; Geisthoff, U.; Gall, H.; Tello, K.; Richter, M.J.; Deubner, L.; Schmiedel, R.; Hecker, M.; et al. Treatment with low-dose tacrolimus inhibits bleeding complications in a patient with hereditary hemorrhagic telangiectasia and pulmonary arterial hypertension. Pulm. Circ. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Vicari-Christensen, M.; Repper, S.; Basile, S.; Young, D. Tacrolimus: Review of pharmacokinetics, pharmacodynamics, and pharmacogenetics to facilitate practitioner’s understanding and offer strategies for educating patients and promoting adherence. Prog. Transplant. 2009, 19, 277–284. [Google Scholar] [CrossRef]
- Wang, Z.; Cuddigan, J.L.; Gupta, S.K.; Meenach, S.A. Nanocomposite microparticles (nCmP) for the delivery of tacrolimus in the treatment of pulmonary arterial hypertension. Int. J. Pharm. 2016, 512, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flieger, D.; Hainke, S.; Fischbach, W. Dramatic improvement in hereditary hemorrhagic telangiectasia after treatment with the vascular endothelial growth factor (VEGF) antagonist bevacizumab. Ann. Hematol. 2006, 85, 631–632. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.; Adams, L.A.; MacQuillan, G.; Tibballs, J.; Driesen, R.V.; Delriviere, L. Bevacizumab reverses need for liver transplantation in hereditary hemorrhagic telangiectasia. Liver Transplant. 2008, 14, 210–213. [Google Scholar] [CrossRef]
- Dupuis-Girod, S.; Ginon, I.; Saurin, J.-C.; Marion, D.; Guillot, E.; Decullier, E.; Roux, A.; Carette, M.-F.; Gilbert-Dussardier, B.; Hatron, P.-Y.; et al. Bevacizumab in Patients with Hereditary Hemorrhagic Telangiectasia and Severe Hepatic Vascular Malformations and High Cardiac Output. JAMA 2012, 307, 948–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bofarid, S.; Hosman, A.E.; Mager, J.J.; Snijder, R.J.; Post, M.C. Pulmonary Vascular Complications in Hereditary Hemorrhagic Telangiectasia and the Underlying Pathophysiology. Int. J. Mol. Sci. 2021, 22, 3471. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073471
Bofarid S, Hosman AE, Mager JJ, Snijder RJ, Post MC. Pulmonary Vascular Complications in Hereditary Hemorrhagic Telangiectasia and the Underlying Pathophysiology. International Journal of Molecular Sciences. 2021; 22(7):3471. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073471
Chicago/Turabian StyleBofarid, Sala, Anna E. Hosman, Johannes J. Mager, Repke J. Snijder, and Marco C. Post. 2021. "Pulmonary Vascular Complications in Hereditary Hemorrhagic Telangiectasia and the Underlying Pathophysiology" International Journal of Molecular Sciences 22, no. 7: 3471. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073471