
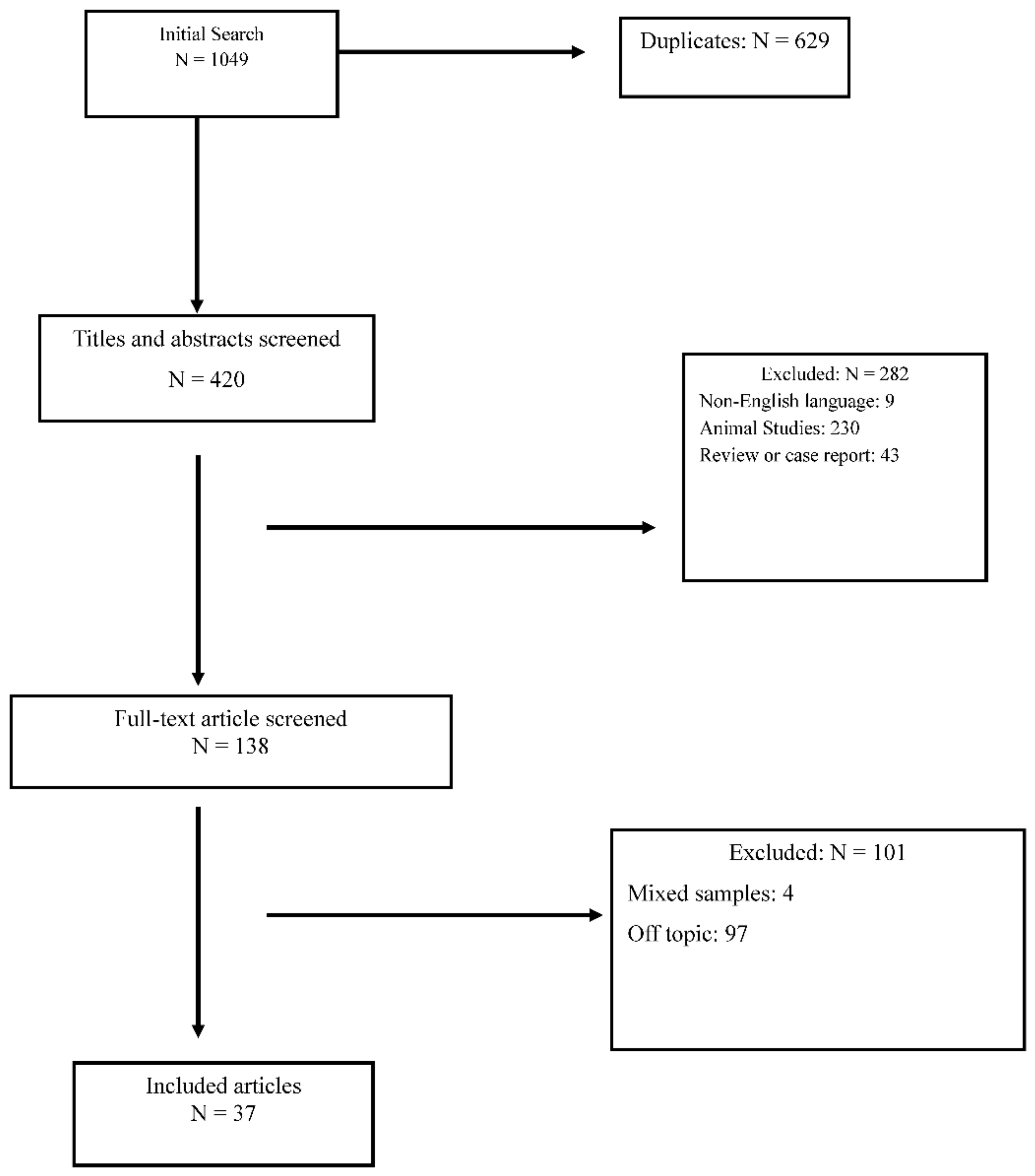
The Role of Cholesterol and Fatty Acids in the Etiology and Diagnosis of Autism Spectrum Disorders

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Methods
3. Results
3.1. ASDs and Cholesterol
3.2. ASDs and Fatty Acids
4. Discussion
- (1)
- a high heterogeneity of study designs, with a wide range of sample sizes, involving in some cases very small groups, so largely limiting the statistical power (rarely reported by authors);
- (2)
- differences in the type of cell or biological material (e.g., RBCs or plasma) used to measure the lipid/fatty acid profile;
- (3)
- the possible influence of concurrent pharmacological treatment on lipid levels and other potential confounding factors, including the concomitance of psychiatric or medical comorbidity;
- (4)
- the heterogeneity in the number of fatty acids tested and/or described in the different reports.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| APOB | apoprotein B |
| ASD | autism spectrum disorder |
| BMI | body mass index |
| CE | cholesterol ester |
| CRP | C-reactive protein |
| CS | cholesterol sulfate |
| DD | developmental delay |
| DHA | docosahexaenoic acid |
| EPA | eicosapentaenoic acid |
| FA | fatty acids |
| FAD | fatty acids deficiency |
| FS | free sterol |
| GF-CF | gluten-casein-free diet |
| GI | gastrointestinal |
| GM1 | ganglioside GM1 |
| HBL | hypobetalipoproteinemia |
| HC | healthy controls |
| HDL-C | high-density lipoprotein |
| ID | intellectual disabilities |
| LDL-C | low-density lipoprotein |
| LFD | low-fat diet |
| MDA-LDL | malondialdehyde-modified low-density lipoprotein |
| PC | phosphatidylcholine |
| PE | phosphatidylethanolamine |
| PS | phosphatidylserine |
| PUFA | polyunsaturated fatty acids |
| RBC | red blood cell |
| RD | regular diet |
| SLOS | Smith-Lemli-Opitz Syndrome |
| TC | total cholesterol |
| TG | triglycerides |
| VLDL | very-low-density lipoprotein |
| 7-DHC | 7-dehydrocholesterol |
| 8-DHC | 8-dehydrocholesterol |
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5; American Psychiatric Press: Washington, DC, USA, 2013. [Google Scholar]
- Carmo, J.C.; Rumiati, R.I.; Siugzdaite, R.; Brambilla, P. Preserved Imitation of Known Gestures in Children with High-Functioning Autism. ISRN Neurol. 2013, 2013, 751516. [Google Scholar] [CrossRef]
- Grecucci, A.; Brambilla, P.; Siugzdaite, R.; Londero, D.; Fabbro, F.; Rumiati, R.I. Emotional Resonance Deficits in Autistic Children. J. Autism Dev. Disord. 2012, 43, 616–628. [Google Scholar] [CrossRef]
- Frye, R.E. Social Skills Deficits in Autism Spectrum Disorder: Potential Biological Origins and Progress in Developing Therapeutic Agents. CNS Drugs 2018, 32, 713–734. [Google Scholar] [CrossRef]
- Calderoni, S.; Bellani, M.; Hardan, A.Y.; Muratori, F.; Brambilla, P. Basal ganglia and restricted and re-petitive behaviours in Autism Spectrum Disorders: Current status and future perspectives. Epidemiol. Psychiatr. Sci. 2014, 23, 235–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, A.; DeMayo, M.M.; Glozier, N.; Guastella, A.J. An Overview of Autism Spectrum Disorder, Heterogeneity and Treatment Options. Neurosci. Bull. 2017, 33, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Piccin, S.; Crippa, A.; Nobile, M.; Hardan, A.Y.; Brambilla, P. Video modeling for the development of personal hygiene skills in youth with autism spectrum disorder. Epidemiol. Psychiatr. Sci. 2017, 27, 127–132. [Google Scholar] [CrossRef]
- DeFilippis, M.; Wagner, K.D. Treatment of Autism Spectrum Disorder in Children and Adolescents. Psychopharmacol. Bull. 2016, 46, 18–41. [Google Scholar]
- Sanchack, K.E.; Thomas, C.A. Autism Spectrum Disorder: Primary Care Principles. Am. Fam. Physician 2016, 94, 972–979. [Google Scholar] [PubMed]
- Bellani, M.; Calderoni, S.; Muratori, F.; Brambilla, P. Brain anatomy of autism spectrum disorders I. Focus on corpus callosum. Epidemiol. Psychiatr. Sci. 2013, 22, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brambilla, P.; Hardan, A.Y.; Di Nemi, S.U.; Caverzasi, E.; Soares, J.C.; Perez, J.; Barale, F. The functional neuroanatomy of autism. Funct. Neurol. 2004, 19, 9–18. [Google Scholar] [PubMed]
- Mengotti, P.; D’Agostini, S.; Terlevic, R.; De Colle, C.; Biasizzo, E.; Londero, D.; Ferro, A.; Rambaldelli, G.; Balestrieri, M.; Zanini, S.; et al. Altered white matter integrity and development in children with autism: A combined voxel-based morphometry and diffusion imaging study. Brain Res. Bull. 2011, 84, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Elsabbagh, M.; Baird, G.; Veenstra-Vanderweele, J. Autism spectrum disorder. Lancet 2018, 392, 508–520. [Google Scholar] [CrossRef]
- Picardi, A.; Fagnani, C.; Medda, E.; Toccaceli, V.; Brambilla, P.; Stazi, M.A. Genetic and environmental influences underlying the relationship between autistic traits and temperament and character dimensions in adulthood. Compr. Psychiatry 2015, 58, 178–188. [Google Scholar] [CrossRef]
- Bellani, M.; Calderoni, S.; Muratori, F.; Brambilla, P. Brain anatomy of autism spectrum disorders II. Focus on amygdala. Epidemiol. Psychiatr. Sci. 2013, 22, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Frye, R.E.; Vassall, S.; Kaur, G.; Lewis, C.; Karim, M.; Rossignol, D. Emerging biomarkers in autism spectrum disorder: A systematic review. Ann. Transl. Med. 2019, 7, 792. [Google Scholar] [CrossRef]
- Zoccante, L.; Viviani, A.; Ferro, A.; Cerini, R.; Cerruti, S.; Rambaldelli, G.; Bellani, M.; Dusi, N.; Perlini, C.; Boscaini, F.; et al. Increased left parietal volumes relate to delayed language development in autism: A structural mri study. Funct. Neurol. 2011, 25, 217–221. [Google Scholar]
- Agostoni, C.; Nobile, M.; Ciappolino, V.; Delvecchio, G.; Tesei, A.; Turolo, S.; Crippa, A.; Mazzocchi, A.; Altamura, C.A.; Brambilla, P. The Role of Omega-3 Fatty Acids in Developmental Psychopathology: A Systematic Review on Early Psychosis, Autism, and ADHD. Int. J. Mol. Sci. 2017, 18, 2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botturi, A.; Ciappolino, V.; DelVecchio, G.; Boscutti, A.; Viscardi, B.; Brambilla, P. The Role and the Effect of Magnesium in Mental Disorders: A Systematic Review. Nutrients 2020, 12, 1661. [Google Scholar] [CrossRef]
- Shen, L.; Liu, X.; Zhang, H.; Lin, J.; Feng, C.; Iqbal, J. Biomarkers in autism spectrum disorders: Current progress. Clin. Chim. Acta 2020, 502, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Calderoni, S.; Billeci, L.; Narzisi, A.; Brambilla, P.; Retico, A.; Muratori, F. Rehabilitative Interventions and Brain Plasticity in Autism Spectrum Disorders: Focus on MRI-Based Studies. Front. Neurosci. 2016, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Penzes, P. Common mechanisms of excitatory and inhibitory imbalance in schizophrenia and autism spectrum disorders. Curr. Mol. Med. 2015, 15, 146–167. [Google Scholar] [CrossRef]
- Giuliano, A.; Saviozzi, I.; Brambilla, P.; Muratori, F.; Retico, A.; Calderoni, S. The effect of age, sex and clinical features on the volume of Corpus Callosum in pre-schoolers with Autism Spectrum Disorder: A case-control study. Eur. J. Neurosci. 2018, 47, 568–578. [Google Scholar] [CrossRef]
- Mohammad-Rezazadeh, I.; Frohlich, J.; Loo, S.K.; Jeste, S.S. Brain connectivity in autism spectrum disorder. Curr. Opin. Neurol. 2016, 29, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Serati, M.; DelVecchio, G.; Orsenigo, G.; Mandolini, G.M.; Lazzaretti, M.; Scola, E.; Triulzi, F.; Brambilla, P. The Role of the Subplate in Schizophrenia and Autism: A Systematic Review. Neuroscience 2019, 408, 58–67. [Google Scholar] [CrossRef]
- Petrov, A.M.; Kasimov, M.R.; Zefirov, A.L. Cholesterol in the Pathogenesis of Alzheimer’s, Parkinson’s Diseases and Autism: Link to Synaptic Dysfunction. Acta Naturae 2017, 9, 26–37. [Google Scholar] [CrossRef]
- Wang, H. Lipid rafts: A signaling platform linking cholesterol metabolism to synaptic deficits in autism spectrum disorders. Front. Behav. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segatto, M.; Tonini, C.; Pfrieger, F.W.; Trezza, V.; Pallottini, V. Loss of Mevalonate/Cholesterol Homeostasis in the Brain: A Focus on Autism Spectrum Disorder and Rett Syndrome. Int. J. Mol. Sci. 2019, 20, 3317. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.M.; Doherty, J.J.; Robichaud, A.J.; Belfort, G.M.; Chow, B.Y.; Hammond, R.S.; Crawford, D.C.; Linsenbardt, A.J.; Shu, H.-J.; Izumi, Y.; et al. The Major Brain Cholesterol Metabolite 24(S)-Hydroxycholesterol Is a Potent Allosteric Modulator of N-Methyl-D-Aspartate Receptors. J. Neurosci. 2013, 33, 17290–17300. [Google Scholar] [CrossRef] [Green Version]
- Nasrallah, H.A. The puzzling relationship between cholesterol and psychopathology. Curr. Psychiatry 2018, 17, 4–5. [Google Scholar]
- Kawicka, A.; Regulska-Ilow, B. How nutritional status, diet and dietary supplements can affect autism. A review. Roczniki Państwowego Zakładu Higieny 2013, 64, 1–12. [Google Scholar] [PubMed]
- Gillberg, C.; Fernell, E.; Kočovská, E.; Minnis, H.; Bourgeron, T.; Thompson, L.; Allely, C.S. The role of cholesterol metabolism and various steroid abnormalities in autism spectrum disorders: A hypothesis paper. Autism Res. 2017, 10, 1022–1044. [Google Scholar] [CrossRef]
- Błażewicz, A.; Szymańska, I.; Astel, A.; Stenzel-Bembenek, A.; Dolliver, W.R.; Makarewicz, A. Assessment of Changes over Time of Lipid Profile, C-Reactive Protein Level and Body Mass Index in Teenagers and Young Adults on Different Diets Belonging to Autism Spectrum Disorder. Nutrients 2020, 12, 2594. [Google Scholar] [CrossRef]
- Lee, R.W.Y.; Corley, M.J.; Pang, A.; Arakaki, G.; Abbott, L.; Nishimoto, M.; Miyamoto, R.; Lee, E.; Yamamoto, S.; Maunakea, A.K.; et al. A modified ketogenic gluten-free diet with MCT improves behavior in children with autism spectrum disorder. Physiol. Behav. 2018, 188, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Peretti, S.; Mariano, M.; Mazzocchetti, C.; Mazza, M.; Pino, M.C.; Di Pianella, A.V.; Valenti, M. Diet: The keystone of autism spectrum disorder? Nutr. Neurosci. 2019, 22, 825–839. [Google Scholar] [CrossRef]
- Bastiaanssen, T.F.S.; Cowan, C.S.M.; Claesson, M.J.; Dinan, T.G.; Cryan, J.F. Making Sense of … the Microbiome in Psychiatry. Int. J. Neuropsychopharmacol. 2019, 22, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Sivamaruthi, B.S.; Suganthy, N.; Kesika, P.; Chaiyasut, C. The Role of Microbiome, Dietary Supplements, and Probiotics in Autism Spectrum Disorder. Int. J. Environ. Res. Public Health 2020, 17, 2647. [Google Scholar] [CrossRef] [Green Version]
- Tierney, E.; Nwokoro, N.A.; Porter, F.D.; Freund, L.S.; Ghuman, J.K.; Kelley, R.I. Behavior phenotype in the RSH/Smith-Lemli-Opitz syndrome. Am. J. Med. Genet. 2001, 98, 191–200. [Google Scholar] [CrossRef]
- Vancassel, S.; Durand, G.; Barthélémy, C.; Lejeune, B.; Martineau, J.; Guilloteau, D.; Andrès, C.; Chalon, S. Plasma fatty acid levels in autistic children. Prostaglandins Leukot. Essent. Fat. Acids 2001, 65, 1–7. [Google Scholar] [CrossRef]
- Goldenberg, A.; Chevy, F.; Bernard, C.; Wolf, C.; Cormier-Daire, V. Circonstances cliniques du diagnostic du syndrome de Smith-Lemli-Opitz et tentatives de corrélation phénotype-génotype: À propos de 45 cas [Clinical characteristics and diagnosis of Smith-Lemli-Opitz syndrome and tentative phenotype-genotype correlation: Report of 45 cases]. Arch. Pediatr. 2003, 10, 4–10. [Google Scholar]
- Bell, J.; MacKinlay, E.; Dick, J.; MacDonald, D.; Boyle, R.; Glen, A. Essential fatty acids and phospholipase A2 in autistic spectrum disorders. Prostaglandins Leukot. Essent. Fat. Acids 2004, 71, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Dziobek, I.; Gold, S.M.; Wolf, O.T.; Convit, A. Hypercholesterolemia in Asperger syndrome: Independence from lifestyle, obsessive-compulsive behavior, and social anxiety. Psychiatry Res. 2007, 149, 321–324. [Google Scholar] [CrossRef]
- Sikora, D.M.; Pettit-Kekel, K.; Penfield, J.; Merkens, L.S.; Steiner, R.D. The near universal presence of autism spectrum disorders in children with Smith–Lemli–Opitz syndrome. Am. J. Med Genet. Part A 2006, 140, 1511–1518. [Google Scholar] [CrossRef]
- Sliwinski, S.; Croonenberghs, J.; Christophe, A.; Deboutte, D.; Maes, M. Polyunsaturated fatty acids: Do they have a role in the pathophysiology of autism? Neuro Endocrinol. Lett. 2006, 27, 465–471. [Google Scholar] [PubMed]
- Tierney, E.; Bukelis, I.; Thompson, R.E.; Ahmed, K.; Aneja, A.; Kratz, L.; Kelley, R.I. Abnormalities of cholesterol metabolism in autism spectrum disorders. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2006, 141B, 666–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meguid, N.A.; Atta, H.M.; Gouda, A.S.; Khalil, R.O. Role of polyunsaturated fatty acids in the man-agement of Egyptian children with autism. Clin. Biochem. 2008, 41, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.G.; Miller, D.; MacDonald, D.J.; MacKinlay, E.E.; Dick, J.R.; Cheseldine, S.; Boyle, R.M.; Graham, C.; O’Hare, A.E. The fatty acid compositions of erythrocyte and plasma polar lipids in children with autism, developmental delay or typically developing controls and the effect of fish oil intake. Br. J. Nutr. 2010, 103, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Wiest, M.; German, J.; Harvey, D.; Watkins, S.; Hertz-Picciotto, I. Plasma fatty acid profiles in autism: A case-control study. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 221–227. [Google Scholar] [CrossRef]
- Kim, E.K.; Neggers, Y.H.; Shin, C.S.; Kim, E.; Kim, E.M. Alterations in lipid profile of autistic boys: A case control study. Nutr. Res. 2010, 30, 255–260. [Google Scholar] [CrossRef]
- El-Ansary, A.K.; Ben Bacha, A.G.; Ayahdi, L.Y.A. Impaired plasma phospholipids and relative amounts of essential polyunsaturated fatty acids in autistic patients from Saudi Arabia. Lipids Health Dis. 2011, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Ansary, A.K.; Bacha, A.G.; Al-Ayahdi, L.Y. Plasma fatty acids as diagnostic markers in autistic patients from Saudi Arabia. Lipids Health Dis. 2011, 10, 62. [Google Scholar] [CrossRef] [Green Version]
- Schengrund, C.L.; Ali-Rahmani, F.; Ramer, J.C. Cholesterol, GM1, and autism. Neurochem. Res. 2012, 37, 1201–1207. [Google Scholar] [CrossRef]
- Fong, B.M.-W.; Tam, S.; Leung, K.S.-Y. Determination of plasma cholesterol sulfate by LC–APCI-MS/MS in the context of pediatric autism. Talanta 2013, 116, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Ghezzo, A.; Visconti, P.; Abruzzo, P.M.; Bolotta, A.; Ferreri, C.; Gobbi, G.; Malisardi, G.; Manfredini, S.; Marini, M.; Nanetti, L.; et al. Oxidative Stress and Erythrocyte Membrane Alterations in Children with Autism: Correlation with Clinical Features. PLoS ONE 2013, 8, e66418. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.M.; Austin, D.W.; Busija, L. Observable essential fatty acid deficiency markers and autism spectrum disorder. Breastfeed. Rev. Prof. Publ. Nurs. Mothers Assoc. Aust. 2014, 22, 21–26. [Google Scholar]
- Moses, L.; Katz, N.; Weizman, A. Metabolic profiles in adults with autism spectrum disorder and intellectual disabilities. Eur. Psychiatry 2014, 29, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Brigandi, S.A.; Shao, H.; Qian, S.Y.; Shen, Y.; Wu, B.-L.; Kang, J.X. Autistic Children Exhibit Decreased Levels of Essential Fatty Acids in Red Blood Cells. Int. J. Mol. Sci. 2015, 16, 10061–10076. [Google Scholar] [CrossRef] [Green Version]
- Esparham, A.E.; Smith, T.; Belmont, J.M.; Haden, M.; Wagner, L.E.; Evans, R.G.; Drisko, J.A. Nutritional and Metabolic Biomarkers in Autism Spectrum Disorders: An Exploratory Study. Integr. Med. (Encinitas) 2015, 14, 40–53. [Google Scholar] [PubMed]
- Jory, J. Abnormal fatty acids in Canadian children with autism. Nutrition 2016, 32, 474–477. [Google Scholar] [CrossRef]
- Mostafa, G.A.; Al-Ayadhi, L.Y. Reduced levels of plasma polyunsaturated fatty acids and serum carnitine in autistic children: Relation to gastrointestinal manifestations. Behav. Brain Funct. 2015, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Yui, K.; Imataka, G.; Kawasak, Y.; Yamada, H. Increased ω-3 polyunsaturated fatty acid/arachidonic acid ratios and upregulation of signaling mediator in individuals with autism spectrum disorders. Life Sci. 2016, 145, 205–212. [Google Scholar] [CrossRef]
- Yui, K.; Imataka, G.; Kawasaki, Y.; Yamada, H. Down-regulation of a signaling mediator in association with lowered plasma arachidonic acid levels in individuals with autism spectrum disorders. Neurosci. Lett. 2016, 610, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Parletta, N.; Niyonsenga, T.; Duff, J. Omega-3 and Omega-6 Polyunsaturated Fatty Acid Levels and Correlations with Symptoms in Children with Attention Deficit Hyperactivity Disorder, Autistic Spectrum Disorder and Typically Developing Controls. PLoS ONE 2016, 11, e0156432. [Google Scholar] [CrossRef]
- Puig-Alcaraz, C.; Fuentes-Albero, M.; Cauli, O. Relationship between adipic acid concentration and the core symptoms of autism spectrum disorders. Psychiatry Res. 2016, 242, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, S.; Wang, M.; Gao, J.; Sun, C.; Wang, J.; Xia, W.; Wu, S.; Sumner, S.J.; Zhang, F.; et al. Potential serum biomarkers from a metabolomics study of autism. J. Psychiatry Neurosci. 2016, 41, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yui, K.; Imataka, G.; Kawasaki, Y. Competitive Interaction between Plasma Omega-3 Fatty Acids and Arachidonic Acid is Related to Down-Regulation of A Signaling Mediator. Med. Chem. 2016, 12, 318–327. [Google Scholar] [CrossRef]
- Cariou, B.; Challet-Bouju, G.; Bernard, C.; Marrec, M.; Hardouin, J.-B.; Authier, C.; Bach-Ngohou, K.; Leux, C.; Pichelin, M.; Grall-Bronnec, M. Prevalence of hypobetalipoproteinemia and related psychiatric characteristics in a psychiatric population: Results from the retrospective HYPOPSY Study. Lipids Health Dis. 2018, 17, 249. [Google Scholar] [CrossRef] [Green Version]
- Howsmon, D.P.; Adams, J.B.; Krüger, U.; Geis, E.; Gehn, E.; Hahn, J. Erythrocyte fatty acid profiles in children are not predictive of autism spectrum disorder status: A case control study. Biomark. Res. 2018, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Toscano, C.V.A.; Carvalho, H.M.; Ferreira, J.P. Exercise Effects for Children With Autism Spectrum Disorder: Metabolic Health, Autistic Traits, and Quality of Life. Percept Mot Skills. 2018, 125(1), 126–146. [Google Scholar] [CrossRef] [Green Version]
- Benachenhou, S.; Etcheverry, A.; Galarneau, L.; Dubé, J.; Çaku, A. Implication of hypocholesterolemia in autism spectrum disorder and its associated comorbidities: A retrospective case–control study. Autism Res. 2019, 12, 1860–1869. [Google Scholar] [CrossRef]
- Hassan, M.H.; Desoky, T.; Sakhr, H.M.; Gabra, R.H.; Bakri, A.H. Possible Metabolic Alterations among Autistic Male Children: Clinical and Biochemical Approaches. J. Mol. Neurosci. 2019, 67, 204–216. [Google Scholar] [CrossRef]
- Usui, N.; Iwata, K.; Miyachi, T.; Takagai, S.; Wakusawa, K.; Nara, T.; Tsuchiya, K.J.; Matsumoto, K.; Kurita, D.; Kameno, Y.; et al. VLDL-specific increases of fatty acids in autism spectrum disorder correlate with social interaction. EBioMedicine 2020, 58, 102917. [Google Scholar] [CrossRef]
- Yui, K.; Imataka, G.; Sasaki, H.; Shiroki, R. The role of lipid peroxidation in individuals with autism spectrum disorders. Metab. Brain Dis. 2020, 35, 1101–1108. [Google Scholar] [CrossRef]
- Crippa, A.; Del Vecchio, G.; Ceccarelli, S.B.; Nobile, M.; Arrigoni, F.; Brambilla, P. Cortico-Cerebellar Connectivity in Autism Spectrum Disorder: What Do We Know So Far? Front. Psychiatry 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- El-Ansary, A.; Chirumbolo, S.; Bhat, R.S.; Dadar, M.; Ibrahim, E.M.; Bjørklund, G. The Role of Lipidomics in Autism Spectrum Disorder. Mol. Diagn. Ther. 2020, 24, 31–48. [Google Scholar] [CrossRef]
- Capuzzi, E.; Caldiroli, A.; Capellazzi, M.; Tagliabue, I.; Auxilia, A.; Ghilardi, G.; Buoli, M.; Clerici, M. Exploring the role of serum lipid profile and neutrophil-to-lymphocyte ratio in violent suicide attempters: A cross sectional study. CNS Spectr. 2020, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Buoli, M.; Caldiroli, L.; Guenzani, D.; Carnevali, G.S.; Cesari, M.; Turolo, S.; Barkin, J.L.; Messa, P.; Agostoni, C.; Vettoretti, S. Associations Between Cholesterol and Fatty Acid Profile on the Severity of Depression in Older Persons with Nondialysis Chronic Kidney Disease. J. Ren. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.D.; Deckler, E.; Jones, M.T.; Jarskog, L.F.; Penn, D.L.; Pinkham, A.E. Autism symptoms, depression, and active social avoidance in schizophrenia: Association with self-reports and informant assessments of everyday functioning. J. Psychiatr. Res. 2019, 115, 36–42. [Google Scholar] [CrossRef]
- Macchi, C.; Favero, C.; Ceresa, A.; Vigna, L.; Conti, D.M.; Pesatori, A.C.; Racagni, G.; Corsini, A.; Ferri, N.; Sirtori, C.R.; et al. Depression and cardiovascular risk—Association among Beck Depression Inventory, PCSK9 levels and insulin resistance. Cardiovasc. Diabetol. 2020, 19, 1–11. [Google Scholar] [CrossRef]
- Brambilla, P.; Bellani, M.; Isola, M.; Bergami, A.; Marinelli, V.; Dusi, N.; Rambaldelli, G.; Tansella, M.; Finardi, A.M.; Martino, G.; et al. Increased M1/decreased M2 signature and signs of Th1/Th2 shift in chronic patients with bipolar disorder, but not in those with schizophrenia. Transl. Psychiatry 2014, 4, e406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellani, M.; Zanette, G.; Zovetti, N.; Barillari, M.; Del Piccolo, L.; Brambilla, P. Adult Mild Encephalitis With Reversible Splenial Lesion Associated With Delirious Mania: A Case Report. Front. Psychiatry 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Guenzani, D.; Buoli, M.; Caldiroli, L.; Carnevali, G.S.; Serati, M.; Vezza, C.; Armelloni, S.; Messa, P.; Vettoretti, S. DREAM Project Group Malnutrition and inflammation are associated with severity of depressive and cognitive symptoms of old patients affected by chronic kidney disease. J. Psychosom. Res. 2019, 124, 109783. [Google Scholar] [CrossRef] [PubMed]
- Corradini, I.; Focchi, E.; Rasile, M.; Morini, R.; Desiato, G.; Tomasoni, R.; Lizier, M.; Ghirardini, E.; Fesce, R.; Morone, D.; et al. Maternal Immune Activation Delays Excitatory-to-Inhibitory Gamma-Aminobutyric Acid Switch in Offspring. Biol. Psychiatry 2018, 83, 680–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qasem, H.; Al-Ayadhi, L.; Bjørklund, G.; Chirumbolo, S.; El-Ansary, A. Impaired lipid metabolism markers to assess the risk of neuroinflammation in autism spectrum disorder. Metab. Brain Dis. 2018, 33, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Loomes, R.; Hull, L.; Mandy, W.P.L. What Is the Male-to-Female Ratio in Autism Spectrum Disorder? A Systematic Review and Meta-Analysis. J. Am. Acad. Child Adolesc. Psychiatry 2017, 56, 466–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Study | Design | Sample | Intervention | Biomarkers | Findings in ASDs |
|---|---|---|---|---|---|
| Tierney et al., 2001 [38] | Cross-sectional study | 28 SLOS children | / | Plasma TC | SLOS, which presents with hypocholesterolemia, is associated with the presence of autism. |
| Vancassel et al., 2001 [39] | Cross-sectional study | 15 ASD subjects: Mean age 8.4 years 73.33% males 18 ID subjects: Mean age 8.8 years 72.22% males | / | Plasma FA | A marked reduction in the levels of DHA (23%) was shown in ASD, resulting in significantly lower levels of n − 3 PUFA (20%; p = 0.032) and a significant increase in the ω6/ω3 ratio (25%; p = 0.039). |
| Goldenberg et al., 2002 [40] | Cross-sectional study | 45 SLOS subjects | / | Plasma TC, 7-DHC, 8-DHC | Plasma hypocholesterolemia correlates significantly with clinical severity (1–2.5% increased risk). |
| Bell et al., 2004 [41] | Cross-sectional study | ASD children HC | / | Plasma and RBC membranes FA composition | Higher frequency of FAD was found in patients with autism and Asperger syndrome compared to HC. Patients with regressive autism had higher percentages of stearic acid, linoleic acid and total saturates in their RBC membranes than HC, as well as higher lignoceric acid, docosapentaenoic acid, nervonic acid and AA/EPA ratio; these latter characteristics are shared by regressive autism and Asperger syndrome. Patients with regressive autism showed lower oleic acid and AA values and patients both with regressive autism and with Asperger syndrome presented lower docosapentaenoic acid and total n − 3 PUFAs compared to HC. |
| Dziobek et al., 2006 [42] | Cross-sectional study | 22 Asperger children: Mean age 40.8 ± 10.8 years 77.3% male 22 HC: Mean age 44.6 ± 14.8 years 77.3% male | / | Plasma TC, LDL-C, TG | Asperger subjects showed statistically significant elevated levels of TC, LDL-C and TG and significant lower levels of HDL compared with HC. After controlling for physical activity, group differences remained significant for TC (p = 0.016) and LDL-C (p = 0.017), but not for HDL-C (p = 0.323) and TG (p = 0.217). |
| Sikora et al., 2006 [43] | Clinical trial | 14 SLOS children: Mean age 7.1 ± 3.5 years | 2-years cholesterol supplementation | Plasma TC, 7-DHC, 8-DHC | Plasma TC, 7-DHC and 8-DHC at baseline and after supplementation did not correlate with the presence or severity of autistic symptoms. |
| Sliwinski et al., 2006 [44] | Cross-sectional study | 16 high-functioning ASD subjects aged 12–18 years: 100% males 22 HC | / | Plasma FA | In ASD there was a significant increase in the fraction of DHA and an increase in the total ω3/ω6 ratio. |
| Tierney et al., 2006 [45] | Cross-sectional study | 100 ASD children | / | Plasma TC, 7-DHC | Of 19% of subjects with low TC (<100 mg/dL), 31.5% met criteria for ASD. |
| Meguild et al., 2008 [46] | Clinical trial | 30 ASD children aged 3–11 years: 60% males 30 age- and sex-matched HC | 3-months fish oil supplementation | Plasma FA | First assessment: In ASD group, linolenic acid showed a significant reduction (71%), followed by DHA (65%) and AA (45%), while linoleic acid was the least affected PUFA (32%), compared to HC. Second assessment: 66% of autistic children showed clinical and biochemical improvement (decrease in Childhood Autism Rating Scale scores: p < 0.0001). Plasma PUFA showed increase in plasma levels after supplementation (all p < 0.0001). |
| Bell et al., 2010 [47] | Cross-sectional study | 49 ASD children: Mean age 7.5 ± 3.5 years 89.80% males 39 DD children: Mean age 6.0 ± 3.3 years 89.74% males 52 HC: Mean age 7.5 ± 3.6 years 94.23% males | / | Plasma and RBC membranes FA composition | RBC and plasma FA in ASD had an increased AA/EPA ratio. Decreased levels of lignoceric acid and nervonic acid were found in children with DD with respect to ASD. ASD subjects consuming fish oil showed reduced erythrocyte AA, adrenic acid, docosapentanaeoic acid and total n − 6 FA and increased EPA, DHA and total n − 3 FA along with reduced n − 6/n − 3 and AA/EPA ratios. |
| Wiest et al., 2009 [48] | Cross-sectional study | 153 ASD children: Mean age 3.75 years 88.89% males 97 HC: Mean age 3.42 years 78.36% males | / | Plasma lipidomics | Levels of CE, FS, PC and FA did not differ between ASD and HC. Total TG, lysophosphatidylcholine, PE and diglyceride levels were higher in the ASD than the HC group (respectively, p = 0.006; p = 0.001; p = 0.007; p = 0.005). DHA was significantly decreased in ASD with respect to HC (p < 0.05). |
| Kim et al., 2010 [49] | Cross-sectional study | 29 ASD children: Mean age, 10.1 ± 1.3 years 100% male 29 age- and sex-matched HC | / | Plasma TC, HDL-C, LDL-C, LDL/HDL ratio, TG | The mean TG level was significantly higher, whereas the mean HDL-C level was significantly lower in cases as compared to controls (respectively, p = 0.01; p = 0.003). There was no significant difference in TC and LDL-C levels between ASD and HC. The LDL/HDL ratio was significantly higher in ASD compared to HC (p = 0.03). Autism was associated with plasma TG, HDL and the LDL/HDL ratio (respectively, p = 0.01; p = 0.0003; p = 0.04). |
| El-Ansary et al., 2011 [50] | Cross-sectional study | 25 ASD children aged 4–12 years 16 HC aged 4–11 years | / | Plasma FA, PE, PS and PC | ASD patients showed higher LA/AA, ALA/DHA, AA/DHA, EPA/AA ratios compared to HC (respectively, p = 0.034; p = 0.004; p = 0.000; p = 0.000). ASD patients showed higher plasma PE, PS and PC compared to HC (respectively, p = 0.002; p = 0.000; p = 0.000). |
| El-Ansary et al., 2011 [51] | Cross-sectional study | 26 ASD children aged 4–12 years 26 age- and sex-matched HC | / | Plasma FA | ASD patients showed an increase in acetic, valeric, hexanoic and stearidonic acid compared to HC (respectively p = 0.000; p = 0.000; p = 0.000; p = 0.009). ASD subjects show different percentage decrease of saturated acids (propionic, butyric, caprylic, decanoic, lauric, palmitic and stearic) together with mono (oleic) and polyunsaturated fatty acids (arachidic, α-linolenic, DHA, linoleic, γ-linolenic, AA, elaidic) compared to HC (respectively, p = 0.000; p = 0.028; p = 0.000; p = 0.000; p = 0.000; p = 0.0037; p = 0.000; p = 0.000; p = 0.000; p = 0.0045; p = 0.000; p = 0.023; p = 0.000; p = 0.000; p = 0.000). |
| Schengrund et al., 2012 [52] | Cross-sectional study | 16 ASD children: Mean age 5.13 years 93.75% male 20 HC: Mean age 6.0 years 40% male | / | RBC membranes composition: GM1 and cholesterol | ASDs children have less cholesterol and more GM1 in their RBC membranes than HC (respectively, p = 0.012; p = 0.019). |
| Fong et al., 2013 [53] | Cross-sectional study | 102 ASD children: Mean age 3.5 years 52.0% male 102 HC: Mean age 3.9 years 52.0% male | / | Plasma CS | Comparison of normal and autistic children showed no statistically significant difference in plasma CS level. |
| Ghezzo et al., 2013 [54] | Cross-sectional study | 25 ASD children: Mean age 7.8 ± 2.23 years 80.95% males 23 HC: Mean age 7.6 ±1.96 years 70% males | / | RBC membranes FA composition | Alteration in RBC FA membrane profile (increase in monounsaturated fatty acids, decrease in EPA and DHA with a consequent increase in ω6/ω3 ratio) were found in ASD compared to HC (respectively, p < 0.01; p < 0.05; p < 0.01). |
| Brown et al., 2014 [55] | Cross-sectional study | 19 ASD children 23 HC (siblings) | / | Plasma FA | Those infants not breastfed (with colostrum) within the first hour of life and who had a history of FAD symptoms were more likely to have an ASD diagnosis. |
| Moses et al., 2014 [56] | Cross-sectional study | 80 adults with ASD 77 adults with ID 828 HC | / | Plasma TC | TC levels of people with ASD and ID were significantly lower than those of HC (p < 0.001) but after adjusting for gender, age and BMI and using Bonferroni correction, the significance was lost. |
| Brigandi et al., 2015 [57] | Cross-sectional study | 121 ASD subjects aged 3–17 years 110 HC aged 3–17 years | / | Plasma and RBC membranes FA composition | The percentage of total PUFA was lower in ASD than in HC; levels of AA and DHA were particularly decreased (p < 0.001). |
| Esparham et al., 2015 [58] | Cross-sectional study | 7 ASD children aged 7–18 years: 71.43% males | / | Plasma FA | An abnormal level of α-linolenic, linoleic acid and high levels of DHA were found, as well as an elevated ω6/ω3 ratio. |
| Jory, 2015 [59] | Cross-sectional study | 11 ASD children: Mean age 3.05 ± 0.79 years 72.73% males 15 HC: Mean age 3.87 ± 1.06 years 40% males | / | Plasma and RBC membranes FA composition | Children with ASD demonstrated lower RBC DHA, EPA, AA and ω3/ω6 ratios (respectively, p < 0.0003; p < 0.03; p < 0.002; p < 0.001). They also demonstrated lower plasma DHA, AA and linoleic acid levels (respectively, p < 0.02; p < 0.05; p < 0.02). |
| Mostafa and Al-Ayadi, 2015 [60] | Cross-sectional study | 100 ASD children: Mean age 6.22 ± 2.1 years 78% males 100 HC: Mean age 5.96 ± 2 years 78% males | / | Plasma FA and carnitine | Reduced levels of plasma carnitine and plasma DHA, AA, linolenic and linoleic acids were found in 66%, 62%, 60%, 43% and 38%, respectively of ASD children. 54% of ASD patients had elevated ω6/ω3 ratio. ASD patients with GI manifestations had significantly increased percentage of reduced serum carnitine (91.7%) and plasma DHA levels (87.5%) than HC (respectively, 42.3%; 38.5%), (respectively, p < 0.001; p < 0.001). |
| Yui et al., 2016 [61] | Cross-sectional study | 28 ASD subjects: Mean age 13.5 ± 4.6 years 71.43% males 21 HC: Mean age 13.9 ± 5.7 years 71.43% males | / | Plasma FA | Plasma EPA, DHA and arachidic acid levels and plasma DHA/AA and EPA/AA ratios were significantly higher in ASD compared to HC (respectively, p = 0.02; p = 0.03; p = 0.04; p = 0.0002; p = 000). Plasma AA and adrenic acid were significantly lower in ASD compared to HC (respectively, p = 0.05; p = 0.04). |
| Yui et al., 2016 [62] | Cross-sectional study | 30 ASD subjects: Mean age 13.6 ± 4.3 years 33.33% males 20 HC: Mean age, 13.2 ± 5.4 years 30% males | / | Plasma FA | The plasma levels of EPA and the plasma ratios of EPA/AA and DHA/AA were significantly higher (respectively, p = 0.1; p = 000; p = 0.000), while the plasma levels of AA and metabolites, such as adrenic acid, were significantly lower in ASD compared to HC (respectively, p = 0.01; p = 0.0; p = 0.004). |
| Parletta et al., 2016 [63] | Cross-sectional study | 85 ASD children 401 ADHD children 79 HC | / | Plasma FA | Children with ADHD and ASD had lower DHA, EPA and AA, higher AA/EPA ratio and lower ω3/ω6 than controls (p < 0.001 except AA between ADHD and controls: p = 0.047). Children with ASD had lower DHA, EPA and AA than children with ADHD (p < 0.001). Childhood Autism Rating Scale scores correlated significantly with DHA, EPA and AA (respectively, p = 0.002; p = 0.038; p = 0.021). |
| Puig-Alcatraz et al., 2016 [64] | Cross-sectional study | 26 ASD children aged 4–13 years 23 HC aged 4–12 years | / | Urinary adipic acid, suberic acid | No increase in the concentration of adipic acid or suberic in children with ASD compared to HC. The increase in adipic acid concentration was significantly and indirectly correlated with the severity of the deficit in socialization and communication skills in ASD children. |
| Wang et al., 2016 [65] | Cross-sectional study | 73 ASD children: Mean age 4.6 ± 0.8 years 80.82% males 63 HC: Mean age 4.1 ± 0.7 years 80.95% males | / | Plasma metabolomics | ASD was associated with 2 metabolites: sphingosine 1-phosphate and DHA (respectively, p < 0.001; p < 0.001). |
| Yui et al., 2016 [66] | Cross-sectional study | 30 ASD subjects: Mean age 13.0 years 20 sex-matched HC: Mean age 13.6 years | / | Plasma FA | ASD had significantly higher plasma DHA/AA and EPA/AA ratios compared to HC. The plasma ceruloplasmin levels in ASD were significantly reduced compared to HC. Multiple linear regression demonstrated that plasma DHA/AA ratio was a fitting model for distinguishing ASD from the HC. |
| Cariou et al., 2018 [67] | Cross-sectional study | 839 adult psychiatric patients: group 1: hypobetalipoproteinemia (HBL) Mean age 35 ± 10 years 65% male group 2: non-HBL Mean age 44 ± 14 years 59% male | / | Plasma TC, HDL-C, LDL-C, TG | Psychiatric patients with HBL were characterized by a higher frequency of specific developmental disorders (including autism) (p = 0.011). |
| Howsmon et al., 2018 [68] | Cross-sectional study | 63 ASD children: Median age 7.8 years: 49 HC: Median age 10.0 years | / | RBC membranes FA composition | FA do not allow for classification at the individual level. |
| Toscano et al., 2018 [69] | Clinical trial | 64 ASD children aged 6–12 years: experimental group: n = 46 control group: n = 18 | 48-week exercise-based intervention | Plasma TC, HDL, LDL | The experimental group showed beneficial effects on metabolic indicators (TC, HDL, LDL), autism traits and parent-perceived quality of life. |
| Benachenhou et al., 2019 [70] | Cross-sectional study | 79 ASD children: Mean age 19.4 ± 12.1 years 81% male 79 HC: Mean age 19.4 ± 12.0 years 81% male | / | Plasma TC, HDL-C, TG, LDL-C | TC levels below the 10th centile were associated with a higher rate of ASD-associated ID (OR = 3.33; 95% CI: 1.26–8.00) and anxiety/depression (OR = 4.74; 95% CI: 1.40–15.73). |
| Hassan et al., 2019 [71] | Cross-sectional study | 63 ASD children 63 age- and sex-matched HC | / | Plasma TC | The serum levels of TC was significantly lower among ASD when compared with HC (p < 0.05). |
| Blazewicz et al., 2020 [33] | Clinical trial | 57 ASD children: 100% male group 1: low-fat diet (LFD), n = 14 Mean age 16.6 years group 2: gluten-casein-free diet (GF-CF), n = 10 Mean age 17.8 years group 3: regular diet (RD), n = 35 Mean age 17.3 years 36 HC: Mean age 17.6 years 100% male RD | Different type of diet: LFD, GF-CF, RD | Plasma CRP, TC, HDL-C, TG | First assessment: in ASD subjects compared to HC: increased BMI, CRP and TC/HDL and decreased HDL-C for all types of diets (p < 0.05) increased TG in the group of LFD (p = 0.003) and RD individual (p < 0.001) increased non-HDL-C in the group of GF–CF (p = 0.008) and RD subjects (p < 0.001) Second assessment: increased levels of TC, non HDL-C and TC/HDL and decreased level of HDL-C for all ASD individuals regardless of diets used (p < 0.05) BMI and CRP increased only for individuals on LFD (BMI: p = 0.029; CRP: p < 0.001) and RD (BMI: p < 0.001; CRP: p > 0.001) |
| Usui et al., 2020 [72] | Cross-sectional study | 152 ASD children 122 HC | / | Plasma FA, lipoprotein analysis | 48 metabolites were identified in the plasma of ASD children by lipidomics (linoleic acid: p = 0.0133; linolenic acid: p = 0.0141; EPA: p = 0.0147; oleic acid: p = 0.0284; EPA: p = 0.0327; AA: p = 0.0395). Among these, increased FA, such as ω3 and ω6, showed correlations with clinical social interaction score and ASD diagnosis (p < 0.05). Specific reductions of plasma VLDL and APOB in ASD were found by large-scale lipoprotein analysis. |
| Yui et al., 2020 [73] | Cross-sectional study | 11 ASD subjects: Mean age 12.3 ± 5.4 years 27.27% males 7 HC: Mean age 10.0 ± 4.1 years 57.14% males | / | Plasma FA, MDA-LDL, superoxide dismutase | Plasma levels of MDA-LDL, EPA, DHA and DHA/AA ratios were significantly higher, while plasma superoxide dismutase levels were significantly lower in ASD than in HC (respectively, p = 0.034; p = 0.000; p = 0.000; p = 0.000; p = 0.006). Multiple linear regression and adaptative Lasso analysis revealed association of increased plasma DHA levels with the Aberrant Behavior Checklists scores and increased plasma MDA-LDL levels. |
| TC Serum/Plasma Levels | Increased | Decreased |
|---|---|---|
| Dziobek et al., 2005 (in Asperger syndrome)—cross-sectional study [42] | Goldenberg et al., 2003—cross-sectional study [40] | |
| Blazewicz et al., 2020—prospective study [33] | Tierney et al., 2006—cross-sectional study [45] | |
| Schengrund et al., 2012—cross-sectional study [52] | ||
| Hassan et al., 2018—cross-sectional study [71] | ||
| Benachenhou et al., 2019—cross-sectional study [70] |
| ω6/ω3 Serum/Plasma Ratio | Increased | Decreased |
|---|---|---|
| Vancassel et al., 2001—cross-sectional study [39] | Jory, 2015—cross-sectional study [59] | |
| Ghezzo et al., 2013—cross-sectional study [54] | Sliwinski et al., 2006—cross-sectional study [44] | |
| Esparham et al., 2015—cross-sectional study [58] | ||
| Mostafa and Al-Ayadhi, 2015—cross-sectional study [60] |
| DHA Serum/Plasma Levels | Increased | Decreased |
|---|---|---|
| Bell et al., 2004—prospective trial [41] | Meguid et al., 2008—clinical trial [46] | |
| Sliwinski et al., 2006—cross-sectional study [44] | Wiest et al., 2009—cross-sectional study [48] | |
| Esparham et al., 2015—cross-sectional study [58] | Ghezzo et al., 2013—cross-sectional study [54] | |
| Yui et al., 2016—cross-sectional study [61] | Brigandi et al., 2015—cross-sectional study [57] | |
| Yui et al., 2016—cross-sectional study [62] | Jory, 2015—cross-sectional study [59] | |
| Yui et al., 2020—cross-sectional study [73] | Mostafa and Al-Ayadhi, 2015—cross-sectional study [60] | |
| Parletta et al., 2016—cross-sectional study [63] | ||
| Wang et al., 2016—cross-sectional study [65] |
| AA Serum/Plasma Levels | Increased | Decreased |
|---|---|---|
| Yui et al., 2016—cross-sectional study [62] | Bell et al., 2004 – prospective trial [41] | |
| Yui et al., 2016—cross-sectional study [66] | Meguid et al., 2008—clinical trial [46] | |
| Brigandi et al., 2015—cross-sectional study [57] | ||
| Jory, 2015—cross-sectional study [59] | ||
| Mostafa and Al-Ayadhi, 2015—cross-sectional study [60] | ||
| Yui et al., 2016—cross-sectional study [61] | ||
| Parletta et al., 2016—cross-sectional study [63] |
| EPA Serum/Plasma Levels | Increased | Decreased |
|---|---|---|
| Bell et al., 2004—prospective trial [41] | Ghezzo et al., 2013—cross-sectional study [54] | |
| Yui et al., 2016—cross-sectional study [61] | Parletta et al., 2016—cross-sectional study [63] | |
| Yui et al., 2016—cross-sectional study [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, C.M.; Buoli, M.; Ciappolino, V.; Agostoni, C.; Brambilla, P. The Role of Cholesterol and Fatty Acids in the Etiology and Diagnosis of Autism Spectrum Disorders. Int. J. Mol. Sci. 2021, 22, 3550. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073550
Esposito CM, Buoli M, Ciappolino V, Agostoni C, Brambilla P. The Role of Cholesterol and Fatty Acids in the Etiology and Diagnosis of Autism Spectrum Disorders. International Journal of Molecular Sciences. 2021; 22(7):3550. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073550
Chicago/Turabian StyleEsposito, Cecilia Maria, Massimiliano Buoli, Valentina Ciappolino, Carlo Agostoni, and Paolo Brambilla. 2021. "The Role of Cholesterol and Fatty Acids in the Etiology and Diagnosis of Autism Spectrum Disorders" International Journal of Molecular Sciences 22, no. 7: 3550. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073550