Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada


Department of Biological Science, University of Alberta, Edmonton, AB T6G 2E9, Canada
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(1), 9; https://0-doi-org.brum.beds.ac.uk/10.3390/d13010009
Submission received: 27 November 2020
/
Revised: 18 December 2020
/
Accepted: 25 December 2020
/
Published: 30 December 2020
(This article belongs to the Special Issue Bird Parasites)
Abstract
:When a species colonizes a new area, it has the potential to bring with it an array of smaller-bodied symbionts. Rock Pigeons (Columba livia Gmelin) have colonized most of Canada and are found in almost every urban center. In its native range, C. livia hosts more than a dozen species of ectosymbiotic arthropods, and some of these lice and mites have been reported from Rock Pigeons in the United States. Despite being so abundant and widely distributed, there are only scattered host-symbiont records for rock pigeons in Canada. Here we sample Rock Pigeons from seven locations across Canada from the west to east (a distance of > 4000 km) to increase our knowledge of the distribution of their ectosymbionts. Additionally, because ectosymbiont abundance can be affected by temperature and humidity, we looked at meteorological variables for each location to assess whether they were correlated with ectosymbiont assemblage structure. We found eight species of mites associated with different parts of the host’s integument: the feather dwelling mites Falculifer rostratus (Buchholz), Pterophagus columbae (Sugimoto) and Diplaegidia columbae (Buchholz); the skin mites: Harpyrhynchoides gallowayi Bochkov, OConnor and Klompen, H. columbae (Fain), and Ornithocheyletia hallae Smiley; and the nasal mites Tinaminyssus melloi (Castro) and T. columbae (Crossley). We also found five species of lice: Columbicola columbae (Linnaeus), Campanulotes compar (Burmeister), Coloceras tovornikae Tendeiro, Hohorstiella lata Piaget, and Bonomiella columbae Emerson. All 13 ectosymbiont species were found in the two coastal locations of Vancouver (British Columbia) and Halifax (Nova Scotia). The symbiont species found in all sampling locations were the mites O. hallae, H. gallowayi, T. melloi and T. columbae, and the lice Colu. columbae and Camp. compar. Three local meteorological variables were significantly correlated with mite assemblage structure: annual minimum and maximum temperatures and maximum humidity in the month the pigeon was collected. Two local meteorological variables, annual maximum and average temperatures, were significantly correlated with louse assemblages. Our results suggest that milder climatic conditions may affect richness and assemblage structure of ectosymbiont assemblages associated with Rock Pigeons in Canada.
1. Introduction
The introduction of a non-native species to a new habitat, whether it be on purpose or by accident, has the potential to also introduce new symbionts [1,2,3]. However, successful colonization of a new area by a host does not automatically mean its symbionts are also successful. ‘Sorting events’ can result in a symbiont failing to become established in a new habitat, which often results in newly introduced hosts having a lower diversity of symbionts than in their native ranges [4,5,6]. Most obviously, for a symbiont to successfully colonize a new habitat it must not “miss the boat” (i.e., it needs to be present in or on at least some of the members of the founding host population). But even if the symbiont is present on members of the founding host population, it still may fail to survive in the new area. If there are too few conspecific symbionts, or if they are highly aggregated among host individuals, the chance at successful establishment decreases [7]. The life cycle of the symbiont also plays a role in successful colonization. Symbionts with indirect life cycles need both intermediate and final hosts, and may have a lower probability of establishment than those with direct life cycles that require only a single host species [5].
Studies of host-symbiont introductions and sorting events have frequently focused on birds and their ectosymbiotic arthropods [5,8,9,10,11,12,13]. Ectosymbionts of birds include feather mites, skin mites, nasal mites, and chewing lice [14,15]. Although not all of these are closely related phylogenetically, they are all permanent ectosymbionts with direct life cycles and have no stages that live off the body of the host. Most of them rely on direct contact between hosts to disperse [16,17], although some species of skin mites and chewing lice can disperse by phoresy on hippoboscid flies [18]. Chewing lice and vane-dwelling feather mites have most often been the subjects of ‘sorting-event’ studies [4,8], while to our knowledge there is no such study that has examined the entire assemblage of ectosymbionts of a host in its novel range.
An avian host that has been incredibly successful expanding its range is the Rock Pigeon (Columbiformes: Columbidae: Columba livia, Gmelin). The original range of the Rock Pigeon, also commonly known as the Rock Dove or just ‘feral pigeon,’ is Eastern Europe, Western Asia, and Northern Africa. Due to human introductions, it is now found on every continent except Antarctica [19]. Within Canada, Rock Pigeons are found year-round in most urban centers as far north as Whitehorse in the Yukon [20]. They were originally brought by French colonists to Nova Scotia’s Port Royal in the early 1600s [21], and with them came at least some of their ectosymbionts. Like many birds, a single Rock Pigeon can host several ectosymbiont species at the same time. However, published ectosymbiont-host records for Rock Pigeons within Canada are scarce and are focused on C. livia populations in only a few provinces; the most detailed records are from Manitoba due to the long-term research of Terry Galloway on feather lice [22,23,24]. Outside of Manitoba, published records of feather lice and other ectosymbionts of C. livia are sparse [25]. This is probably due to a combination of lack of reporting and true absence of particular ectosymbiont species from some regions. For example, there is only one published report of the vane-dwelling feather mite Falculifer rostratus (Buchholz) (Astigmata: Pterolichoidea: Falculiferidae) in Canada, from Rock Pigeons in Ontario [26]; however, in previous surveys of feather mites of birds in Western Canada, one of us (H.P.) observed this mite species in great numbers on C. livia in coastal British Columbia (Vancouver). Conversely, F. rostratus has not been found on Rock Pigeons from Edmonton in the adjacent province of Alberta (H.P., personal observation).
There are several potential explanations for the apparent lack of ectosymbionts on a host that has recently expanded its range. It could be due to sampling error; if a symbiont species is present in low numbers and at low prevalence, it is possible that it has yet to be detected. If so, increased sampling should reveal this and possibly other as-yet unrecorded ectosymbionts. Alternatively, the symbiont may truly be absent from the host in its new range. Such an absence could arise in two ways: (1) the symbiont may have “missed the boat” and was never on any of the host individuals that colonized the new area; (2) the symbiont may have made it to the new area but failed to survive and reproduce. A drawback to being a permanent ectosymbiont is that there is little control over where you go and little control over the abiotic environment you are exposed to. Rock Pigeons are sedentary (non-migratory) and overwinter in the same area that they spend the summer. Returning to the original example, Falculifer rostratus may not be found in Edmonton because the cold, dry winter environment of central Alberta is not suitable for its survival. Abiotic factors have been shown to influence some avian mites and lice [27,28,29]. This has been reported for vane-dwelling feather mites of the family Proctophyllodidae on passerine hosts in the Cantabrian Mountains of Spain [30]. The authors found that as elevation increased, there was a constant decline in the mean number of feather mites on infested hosts. This decline strongly correlated with the decline in temperature along the elevation gradient. Humidity has also been shown to influence ectosymbiont abundance. For example, chewing louse prevalence and intensity on two species of doves (Columbidae) were significantly lower in an arid environment (Tucson, Arizona) than in a humid one (Weslaco, Texas) [27]. Response to humidity appears to be taxon-specific [13], as some louse species are scarce in areas with long humid seasons [31] and some can survive arid periods through their more resistant eggs [28].
In this study we sampled C. livia from seven locations across Canada, including humid coastal areas and drier inland areas, with two goals: (1) to increase our knowledge of the distribution and diversity of ectosymbionts of this important introduced species in Canada; and (2) to test whether geographic variation in ectosymbiont assemblages of C. livia is more suggestive of a history of “missing the boat” or of being influenced by abiotic factors including the cold, dry winter weather that characterizes much of central Canada.
2. Materials and Methods
2.1. Collection and Identification of Ectosymbionts
Rock Pigeons were salvaged from nine rehabilitation centers and one window-kill-monitoring program (see Acknowledgements). The areas sampled spanned Canada from Vancouver on the west coast to Halifax on the east coast, a straight-line distance of ~4400 km (Figure 1): Vancouver, British Columbia; Calgary, Alberta; Edmonton, Alberta; Saskatoon, Saskatchewan; Winnipeg, Manitoba; Kingston, Ontario; Belleville, Ontario; Toronto, Ontario; and Halifax, Nova Scotia (see Table 1 for sample sizes of Rock Pigeons, and Table S1 for latitudes, longitudes, and collection dates). Due to the similar latitude and relatively close proximity of Kingston, Belleville and Toronto (~240 km), data from Rock Pigeons from these locations were combined and are referred to as from Southern Ontario (S. Ontario). A few pigeons came from nearby locations outside of the city in which the rehabilitation center was located.
At the rehabilitation centers the Rock Pigeons were euthanized, individually bagged, then frozen immediately. Rock Pigeons provided by the Fatal Light Awareness Program (FLAP) in Toronto were found dead as window-kills. Depending on the location, frozen pigeons were picked up in person by A.G. or were shipped overnight to the University of Alberta. Ectosymbionts were removed by washing the pigeons according to the procedure outlined in Grossi et al. [33], with the addition of a nasal rinse (20 mL of water was forced through each nostril using a syringe, with backwash being collected in the general washings bucket). Washings were passed through a 43 µm sieve, a mesh size fine enough to retain small skin mites. Ectosymbionts collected from the pigeon washings were stored in 95% ethanol.
A.G. examined each washing under a dissecting microscope (Leica MZ16, Leica Microsystems Inc.. All lice (adults and nymphs) were counted and removed and a subset of each morphospecies of mite was removed. Due to the large number of mites present, they were only identified to species but were not enumerated. Since Rock Pigeons from Toronto, Ontario were collected by FLAP as already dead birds, it was not known how long the birds had been dead prior to collection. Since lice are known to leave the body of a dead host [34], we used louse data from Toronto birds only for calculation of prevalence and not for intensity. Slide-mounted representatives of each louse and mite morphotype were made by A.G. These specimens were first cleared in 85% lactic acid (Fisher Scientific, Fair Lawn, NJ, USA) for 24 h, and then slide mounted in commercially available phenol-free poly-vinyl alcohol medium (PVA; BioQuip Products, Rancho Dominguez, CA, USA). Slides were cured for 4 days at ca. 40 °C on a slide-warmer and then examined using Differential Interference Contrast optics on a Leica DMLB compound microscope. Specimens were identified using relevant taxonomic literature [14,15,25,35,36,37,38,39,40] and exemplars of each species were deposited in the E.H. Strickland Entomological Museum at the University of Alberta (accession numbers UASM80575–UASM80587).
2.2. Ectosymbiont Assemblages
Prevalence, the proportion of host individuals infested with one or more individuals of a particular symbiont species [41], was calculated for each species of ectosymbiont at each sampling location. Mean intensity, the average number of a particular symbiont among infected hosts [41], was calculated for each louse species at each sampling location. We used the Quantitative Parasitology software package [42] to calculate both metrics.
Non-metric multidimensional scaling (NMDS) was used to explore differences in ectosymbiont assemblages among sampling locations. NMDS was conducted on the mite and louse assemblages separately. For these analyses, the mite dataset consisted of presence/absence data and the louse dataset consisted of count data. NMDS plots were produced using the “metaMDS” function in the vegan package v. 2.4–6 [43] in the R statistical program [44]. Jaccard distance was used for the mite prevalence data and the louse count data were first log (x + 1) transformed in R and Bray–Curtis distance was used. To examine observed differences in ectosymbiont assemblages based on sampling location, permutational multivariate analysis of variance (PERMANOVA) was performed on each dissimilarity matrix using the “adonis” function in the vegan package, with 999 permutations. This was followed by post hoc pairwise comparisons between sampling locations, with the p-values adjusted using the Bonferroni correction [45].
2.3. Potential Effects of Local Climate on Ectosymbiont Assemblages
To assess potential effects of meteorological variables on ectosymbiont assemblages, distance-based redundancy analysis (dbRDA) was conducted on the mite data and a redundancy analysis (RDA) on the louse data. A dbRDA was used to analyze the mite data since it was collected as presence/absence, whereas the louse data consisted of counts. Meteorological data were assembled from Environment Canada (https://climate.weather.gc.ca/prods_servs/cdn_climate_summary_e.html). These data consisted of records from the collection location of the pigeon, including the following: minimum, maximum, and average temperature and humidity and total precipitation from the month before each bird was euthanized ( = ‘previous monthly’); from the month that the bird was euthanized ( = ‘monthly’); and for the month that the bird was euthanized plus the previous 11 months ( = ‘annual’). Previous monthly variables were included to account for louse generation time, with the thought that assemblages present at the time of the host’s death could be strongly influenced by past favorable or unfavorable conditions. Some pigeons had no collection date associated with them, and therefore meteorological data could not be associated with those individuals. This caused sample sizes of hosts to differ from those in previous analyses (see Table 1) for several sites: Calgary n = 28, Edmonton n = 18, Saskatoon n = 0, S. Ontario n = 5 (lice), 7 (mites), Halifax n = 21.
Since meteorological variables were measured in units with different scales, these data were standardized. To reduce collinearity among meteorological variables, stepwise reduction was performed based on variance inflation factor (VIF), in which variables with the largest VIF values were removed until only variables with VIF values less then 5 remained [46]. This was done for the mite and louse data separately using the “vif” function from the “car” package in R. For mites, the dbRDA was performed using the “capscale” function in the vegan package using Jaccard distance. For lice, data were first transformed (log (x + 1)) and then an RDA was performed using the “rda” function in the vegan package. In both cases, meteorological variables were selected using forward model choice with the “OrdiR2step” function in the vegan package in R. p-Values of selected meteorological variables were corrected for multiple comparisons using the Holm–Bonferroni method. Variation partitioning was used to determine how much of the variation was explained by the selected meteorological variables vs. spatial variation. The spatial component was calculated by transforming the latitude and longitude of pigeon collection locations using distance-based Moran’s eigenvector maps spatial eigenfunctions with the package “adespatial,” and variation partitioning was carried out with the “varpart” function.
3. Results
3.1. Ectosymbiont Diversity, Prevalence, and Intensity
We examined a total of 162 Rock Pigeons from the seven sites in Canada and recovered 13 ectosymbiont species. Eight of these were mites: the feather-dwelling mites F. rostratus, Pterophagus columbae (Sugimoto), and Diplaegidia columbae (Buchholz); the skin mites Harpyrhynchoides gallowayi Bochkov, OConnor and Klompen, Harpyrhynchoides columbae (Fain), and Ornithocheyletia hallae Smiley; and the nasal mites Tinaminyssus melloi (Castro) and Tinaminyssus columbae (Crossley). In total, 48,025 lice were collected, representing five species: Columbicola columbae (Linnaeus), Campanulotes compar (Burmeister), Coloceras tovornikae Tendeiro, Hohorstiella lata Piaget, and Bonomiella columbae Emerson. Additionally, two other louse species normally associated with passerine birds, Myrsidea sp. (1 female; 1 male; 1 nymph) and Machaerilaemus maestus (Kellogg and Chapman) (2 females) were each found on a single host from Calgary and Nova Scotia, respectively. Due to their occurrence, each on only a single host individual, and the low number of individuals collected, we believe these are ‘stragglers’—lice found on atypical hosts where they are assumed to be unable to maintain populations [47]. Although these lice may be contaminants due to different species of birds being housed together in rehabilitation centers, stragglers have been shown to be the precursor for a host-switching events [47,48], and therefore their presence should be noted.
The feather-dwelling mites F. rostratus, P. columbae, and D. columbae had their highest prevalences in Vancouver and Halifax and while they all had different distributions across sampling locations, they were all absent from Saskatoon (Table 1; Figure 2A–C).
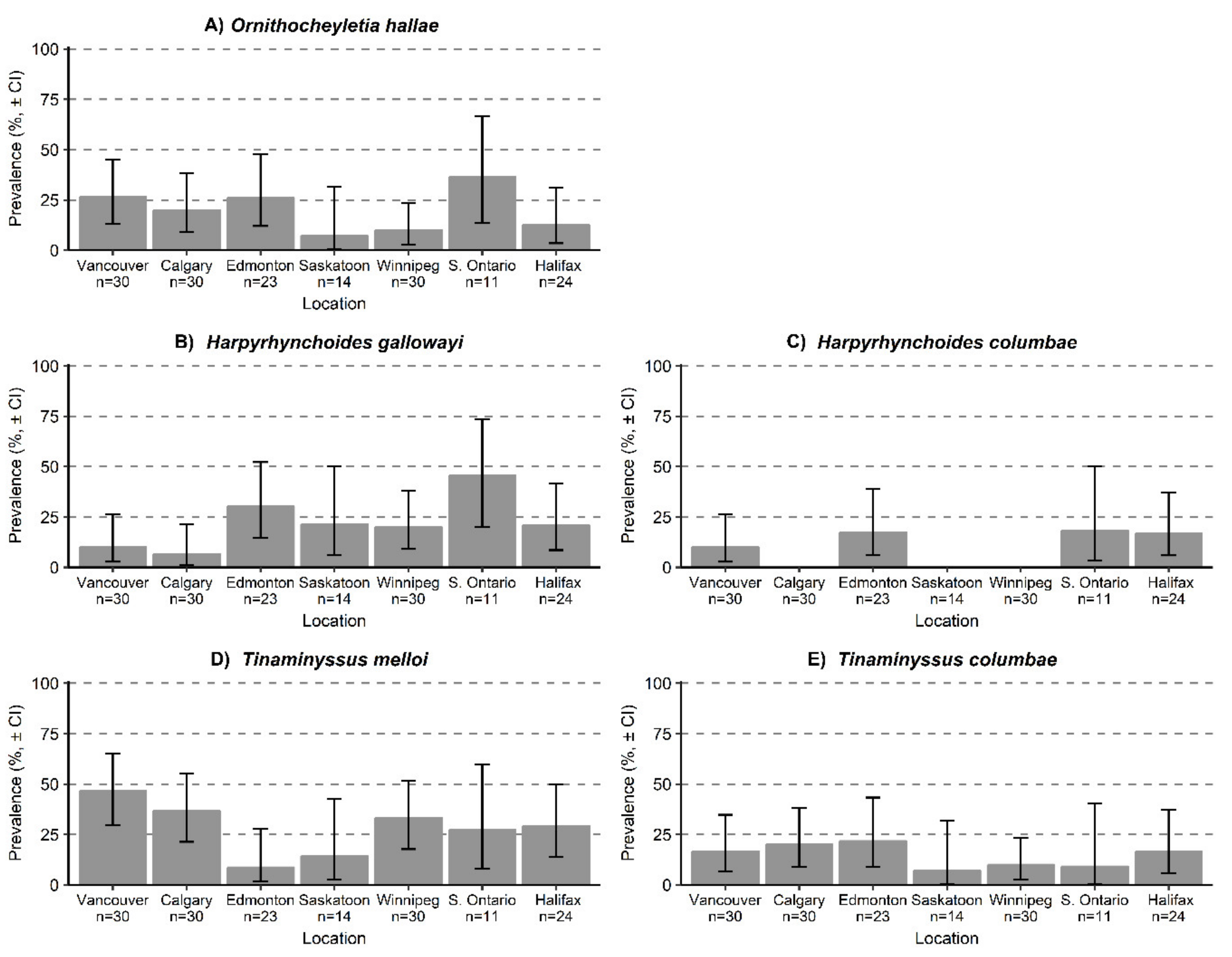
Two of the three skin mite species, O. hallae and H. gallowayi, were found in all locations, while H. columbae was found infesting pigeons from Vancouver, Edmonton, Southern Ontario, and Halifax (Table 1; Figure 3A–C). All three skin mites had their highest prevalences in Southern Ontario. Both species of Tinaminyssus nasal mites were found in every location (Table 1). Tinaminyssus melloi had the highest prevalence in Vancouver and the lowest prevalence in Edmonton (Figure 3D), while T. columbae had the highest prevalence in Edmonton and the lowest in Southern Ontario (Figure 3E).
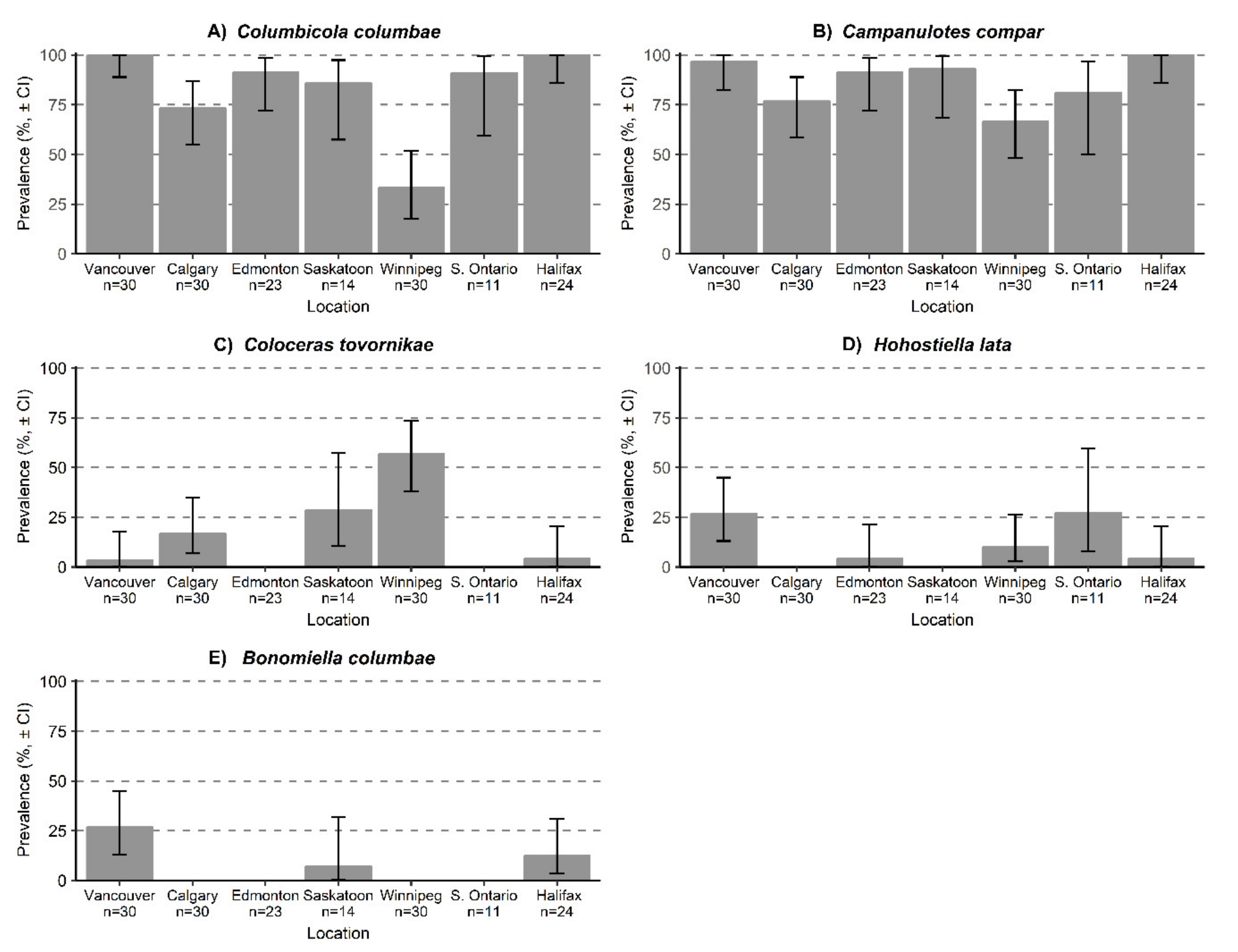
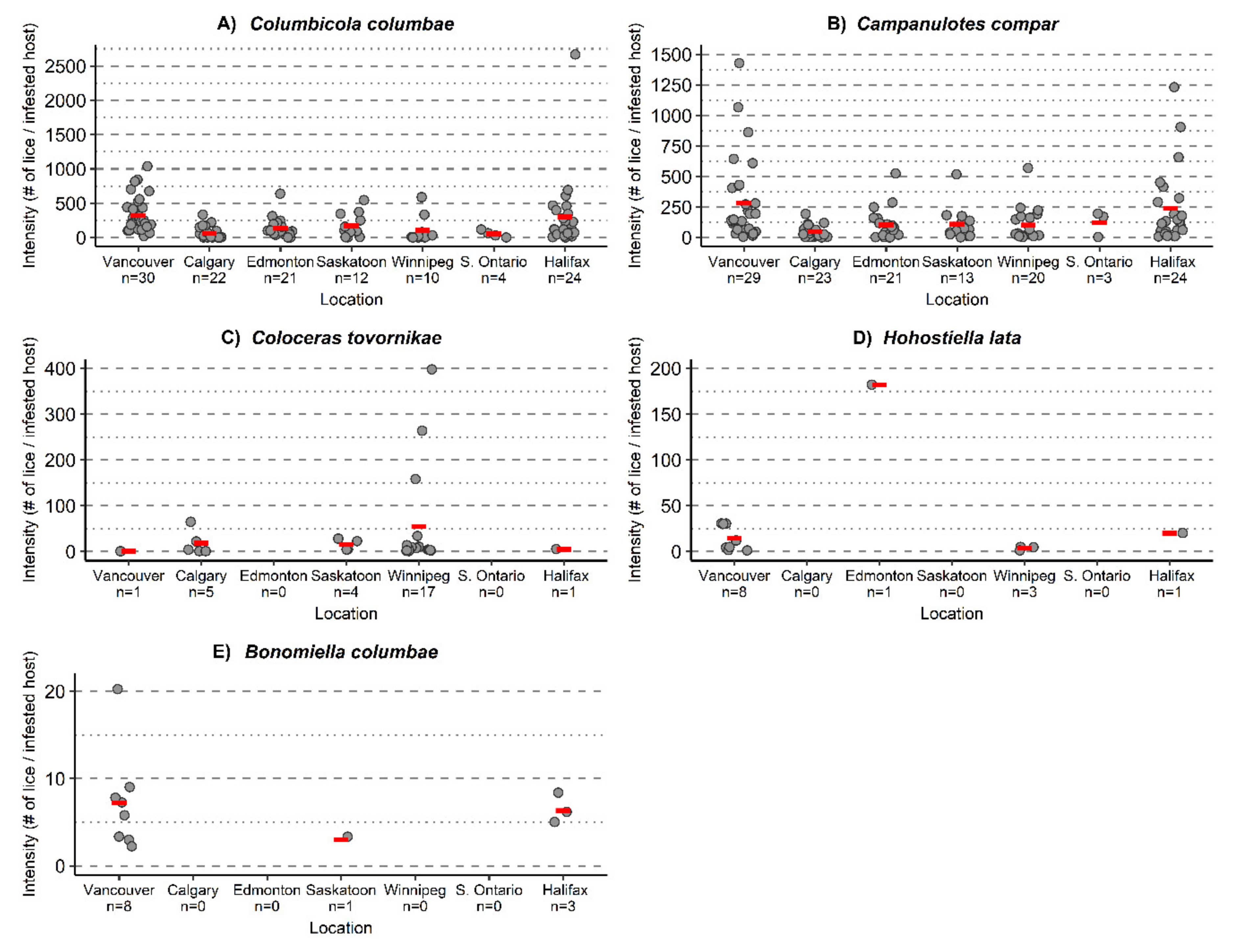
Of the lice, only two species were found in every location: Colu. columbae and Camp. compar (Table 2); both species had high prevalences in all locations (Figure 4A,B). Both species had their highest mean intensities in Vancouver, followed by Halifax: Colu. Columbae—Vancouver (326.1 lice/infested pigeon), Halifax (303.1 lice/infested pigeon); Campanulotes compar—Vancouver (282.0 lice/infested pigeon), Halifax (239.2 lice/infested pigeon) (Figure 5A,B; Table 3). Coloceras tovornikae was found in Vancouver, Calgary, Saskatoon, Winnipeg, and Halifax (Figure 4C) with the highest prevalence and mean intensity in Winnipeg (56.7%, 54.2 lice/infested pigeon, respectively) (Figure 5C). Hohorstiella lata was found in Vancouver, Edmonton, Winnipeg, and Halifax (Figure 4D) with the highest prevalence in Vancouver (26.7%), and the highest mean intensity in Edmonton (182.0 lice/infested pigeon) (Figure 4D); however, this is based on only a single heavily infested individual (Figure 5D). In all other locations the mean intensities for H. lata were less then 20 lice/infested pigeon. Bonomiella columbae was found in Vancouver, Saskatoon, and Halifax; however only one individual pigeon was infested in Saskatoon (Figure 4E). The mean intensity of B. columbae was low with the highest being 7.3 lice/infested pigeon in Vancouver (Figure 5E).
3.2. Assemblage Structure
The NMDS plot for mites based on presence-absence data (Figure 6) shows overlap in the mite assemblages between the different locations. To examine if there were significant differences in assemblages between locations, a PERMANOVA was conducted; this showed that there was a significant difference in assemblage structure between the locations (df = 6, F-value = 7.01, R2 = 0.284, p = 0.001). Pairwise comparisons among locations revealed that mite assemblages in Vancouver were significantly different from every other location except Halifax and that Halifax’s mite assemblages were significantly different from every other location other than Vancouver (Table S2). The louse NMDS plot based on intensity data (Figure 7) shows even more overlap between the locations with most overlaid on each other except for Winnipeg, which was influenced by the high intensities of Colo. tovornikae. The PERMANOVA for the lice was also significant (df = 6, F-value = 4.40, R2 = 0.162, p = 0.001). Pairwise comparisons among locations revealed that louse assemblages in Winnipeg were significantly different from those in Vancouver, Edmonton, and Halifax, and that Vancouver was significantly different from Calgary (Table S2).
3.3. Potential Effects of Local Climate on Ectosymbiont Assemblages
The dbRDA model for mite assemblages was significant (p = 0.001), and three local meteorological variables were found to potentially influence mite assemblages: annual maximum and minimum temperatures and monthly maximum humidity (Figure 8). All three variables were still significant after Holm–Bonferroni correction (Table S3). Through variance partitioning, the meteorological variables alone explained 7.8% of the variation, spatial variables alone explained 0.8%, and the meteorological × spatial interaction explained 11.7% of assemblage composition, leaving 79.7% of the variance unexplained.
The RDA model for lice was also significant (p = 0.001) and five meteorological variables were found to potentially influence louse assemblages: annual maximum, minimum and average temperatures, monthly maximum humidity, and previous monthly minimum temperature (Figure 9). After Holm–Bonferroni correction, only annual maximum and average temperatures remained significant (Table S3). Through variance partitioning, the meteorological variables alone explained 4.8% of the variation, spatial variables alone explained 1.1%, and the environmental × spatial interaction explained 9.6% of assemblage composition, leaving 84.5% of the variance unexplained.
4. Discussion
Based on published literature, of the thirteen ectosymbiont species we found on Rock Pigeons, four (all mites) are new records for Canada: Ornithocheyletia hallae, which has been recorded from Rock Pigeons in Brownsville Texas [36], and Pterophagus columbae, Diplaegidia columbae, and Harpyrhynchoides columbae, which have not been reported from Rock Pigeons in North America. All other mite and louse species were previously recorded from Rock Pigeons in Canada [25,38,39,49,50]. In addition, Passeroptes bispinosa (Banks), a skin mite, was previously recorded on Rock Pigeons in Ontario [51], but was not found infesting any of the pigeons used in this study. There are fifteen additional species of permanent ectosymbionts that have been recorded from Rock Pigeons worldwide that were not found in this survey. Two louse species, Colpocephalum turbinatum Denny and Columbicola tschulyschman Eichler, have been recorded from Rock Pigeons in Louisiana and Utah, respectively [52,53]. The remaining thirteen ectosymbionts have never been recorded on Rock Pigeons within North America: three skin mites (Myialges anchora Sergent and Trouessart, Myialges lophortyx [Furmann and Tharshis], and Rivoltasia bifurcata [Rivolta]), four feather-dwelling mites (Megninia cubitalis Mégnin, Pterophagoides paradoxus Gaud and Barré, Pterophagus strictus Mégnin, and Pterolichus obtusus Robin) and six louse species (Coloceras aegypticum [Kellogg and Paine], Coloceras damicorne [Nitzsch], Coloceras israelensis [Tenderio], Coloceras liviae [Tenderio], Colpocephalum afrozeae Naz, Sychra and Rizvi, and Physconelloides zenaidurae [McGregor]) [14,18,21,54,55,56,57,58,59,60,61].
The distribution of ectosymbionts was not homogeneous across all sampling locations. There were two locations, Vancouver and Halifax, in which all ectosymbiont species found in this study were present. It should be noted that prevalence was not invariably high at these two locations, as some ectosymbiont species were only found infesting one host individual (Table 1 and Table 2). Mite assemblages in Vancouver and Halifax were significantly different from those in other sampling locations (Table S2); however, this pattern was not seen in lice. In addition to high ectosymbiont richness in the two coastal sites, the mite species Pterophagus columbae was only found in these locations. It is perhaps not surprising that pigeons in Halifax are host to this high diversity of ectosymbiont species since this is a known point of introduction [21]. The equally high diversity in Vancouver, and sharing of the otherwise rare species P. columbae, was not expected as these two locations are the most distant from each other. However, both Vancouver and Halifax are port cities, and therefore have coastal elements influencing their climates. It is also possible that C. livia and its ectosymbionts could have been introduced independently to Vancouver via importation of pigeons from Europe; however, we know of no such importation records. Some of the Vancouver population of pigeons (and populations of ectosymbionts) may be descended from birds brought into ports in the Pacific Northwest of the United States. If such importation records exist in grey literature (e.g., government documents), they may provide clues as to the timing and source of imported pigeons in this area.
Of the ectosymbionts collected in this study, one louse (Colu. columbae) and one skin mite (O. hallae) are known to be phoretic on the Pigeon Louse Fly Pseudolynchia canariensis (Macquart) (Diptera: Hippoboscidae) [18,53]. Pseudolynchia canariensis has been recorded from North America and is most abundant in warm climates [62,63]. Pseudolynchia canariensis was not found on any of the pigeons examined in this study. To our knowledge there is only a single Canadian record of P. canariensis in the Canadian National Collection of Insects, Arachnids and Nematodes (CNC), from Windsor in Southern Ontario (personal commentary, Jeffrey Skevington, CNC). Phoresy on this particular species of louse fly is therefore unlikely to be an important mode of dispersal for pigeon ectosymbionts in Canada; however, it is possible that other hippoboscid species may occasionally serve as phoretic hosts.
For this study Rock Pigeons were mainly acquired from rehabilitation centers, which presents two main drawbacks. First, there is no control over the number of pigeons that were turned in to the rehabilitation centers, which resulted in an uneven sample size of hosts across sampling locations and seasons. Therefore, it is possible that ectosymbionts in some locations were not detected, especially in locations that have smaller sample sizes, and for ectosymbiont species that have low prevalence. Second, there was no control over the health of the pigeons sampled. If the reason the pigeon was admitted to the center (injury, illness) also hindered its ability to preen and scratch, this could cause their ectosymbiont loads to be higher than those found on healthy pigeons [64]. The vane-dwelling feather mite Falculifer rostratus was never observed on pigeons from Edmonton prior to our study (H. Proctor, unpublished), and was not found on the 23 pigeons we examined from Edmonton. This species was also absent from pigeons from Saskatoon. Falculifer rostratus as well as the down-dwelling Diplaegidia columbae had highest intensities in Vancouver and Halifax, and the vane mite Pterophagus columbae was only found in these locations. All three of these mite species live on feathers and therefore are more exposed to local climatic changes than mites that live closer to the surface of the host’s body. In contrast, skin mites and nasal mites that may be less exposed to external temperature and humidity did not have elevated prevalences in coastal sampling locations and were found infesting pigeons in almost every location sampled.
The meteorological variables that were significant in our analysis of mite assemblages were annual maximum and minimum temperatures and monthly maximum precipitation. Both temperature and humidity have been shown to influence feather-dwelling mites. As mentioned in the Introduction, one study [29] found that mean number of vane-dwelling feather mites on passerine hosts declined as temperatures declined [30]. With regard to the impact of humidity, vane-dwelling mites do not feed on fluids produced by their host and their sources of food may not provide adequate amounts of water [65]. Instead, it is likely that much of their water comes from the atmosphere; if true, this would explain why locations like Edmonton and Saskatoon that are very dry in the winter appear to be less suitable for these feather mites.
Columbicola columbae and Campanulotes camper were the only louse species found infesting pigeons in all seven sampling locations. Both species generally had high prevalences, with Colu. columbae achieving 100% prevalence on pigeons sampled from Vancouver and Halifax. The other louse species (Colo. tovornkiae, H. lata, and B. columbae) all had patchy distributions with, on average, much lower mean intensities then those shown by Colu. columbae and Camp. camper. The meteorological variables that were significant in our analysis of louse assemblages were annual maximum and average temperatures. Nelson and Murray [66] experimentally manipulated the temperature that Colu. columbae, Camp. compar, and H. lata were raised at and found that temperatures between 32–40 °C were optimal for oviposition and egg development.
While annual changes in louse intensities were not examined in this study, Galloway and Lamb [67] examined 542 Rock Pigeons from Winnipeg, Manitoba, over a 10-year period. They found that Colu. columbae and Camp. compar both had annual peaks in September in abundance of approximately 100 lice per bird with the lowest abundances from February to March. In contrast, the less abundant species Coloceras tovornikae and H. lata both peaked in March with approximately six lice per bird, and C. tovornikae had a second peak in August. Unfortunately a similar study has not been done on the mites infesting Rock Pigeons, however other species of vane-dwelling feather mites also show annual changes in abundance [68,69,70].
In both our study and that of Galloway and Lamb [67], Colo. tovornikae, B. columbae, and H. lata had prevalences usually less than 30% and mean intensities usually less than 20 lice per bird. Their patchy distribution may therefore be due to a naturally low prevalence and intensity, which would both make it harder for them to successfully establish in new areas and make them more difficult to detect in surveys even when they are present. In the city of Boston (MA), in the northeastern United States, Brown [71] examined 72 Rock Pigeons and found H. lata on 41.7%. Although this is a slightly higher prevalence then what is seen in Canada, there was still a low intensity of 6.2 lice per bird. In contrast, in a study conducted in Pakistan, which is part of the native range for Rock Pigeons, H. lata had a prevalence of 51.4% from 68 examined Rock Pigeons and a much higher mean intensity of 230.1 lice per bird [72]. Additionally, in this study they found high mean intensities of Colu. columbae (438.2 lice per bird) and Camp. compar (614.3 lice per bird) while having mid-range prevalences of 70.4% and 58.8%, respectively [72]. However, the trend of high mean intensities does not carry through the whole native range. A study that looked at 50 pigeons in the Canary Islands found Colu. columbae had a prevalence of 100% and mean intensity of 111.4 lice per bird [73], while a study in Nigeria (n = 240) Colu. columbae had a prevalence of 63.8% and a mean intensity of 17.9 lice per bird [74]. Even within their native range, it appears that infestation parameters vary depending on location. It should be noted that in Boston, Colu. columbae and Camp. compar had prevalences and mean intensities that fell within the range that was seen in Canada [71].
The two ‘straggler’ louse species found in this study, Myrsidea sp. and Machaerilaemus maestus, appear to be true stragglers as they have never been recorded on Rock Pigeons before. While these probably do not represent preludes to host-switching events, host switching has occurred on Rock Pigeons before. Gunabopicobia zumpti (Lawrence) (Syringophilidae), a quill mite previously only recorded from Streptopelia capicola (Sundevall) and Streptopelia senegalensis (Linnaeus) (Columbiformes) in South Africa, was found infesting Rock Pigeons in the United States in 1999 [75] and subsequently in Poland [76].
5. Conclusions
Ectosymbiont-host records such as the ones reported here, which are based on numerous locations with basic infestation parameters, give us a baseline of information on what is present. This information is essential for recognizing host-switching events, range extensions, and changes in life histories.
While it appears that some ectosymbiont species missed the boat when Rock Pigeons were introduced to, and spread throughout, Canada (see Discussion, first paragraph), a fair number have been successful in establishing themselves with their host. In this study 13 ectosymbiont species were found infesting pigeons in Canada. Pigeons in the two coastal locations had every ectosymbiont species identified in this study. We could find no other published studies that look at large-scale distributions of multispecies assemblages of ectosymbionts on avian hosts. It is therefore unclear if the ’bi-coastal’ phenomenon we observed is restricted to ectosymbionts of C. livia or if this trend of higher ectosymbiont diversity in coastal vs. inland regions is also true for other widespread host species.
The variation seen in ectosymbiont assemblages does not suggest that mites and lice of Rock Pigeons are failing to reach different parts of Canada due to ‘missing the boat’, with perhaps the exceptions of the lice Colo. tovornikae, B. columbae, and H. lata. Local climatic differences explain statistically significant variation in ectosymbiont assemblages and appear to affect feather-dwelling mites most strongly. However, large portion of unexplained variation (~80% for mites and ~85% for lice) remains. Ideally, one would have a more structured sampling scheme with regular collections from pigeons throughout the year, and manipulative experiments to determine how much influence temperature and humidity have on the ectosymbionts.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1424-2818/13/1/9/s1, Table S1. Collection information for Columba livia; Table S2. Results from pairwise permutational analysis of variance; Table S3. Environmental variables chosen through forward selection.
Author Contributions
Conceptualization, A.G. and H.P.; methodology, A.G.; formal analysis, A.G.; data curation, A.G.; writing—original draft preparation, A.G.; writing—review and editing, H.P.; supervision, H.P.; funding acquisition, H.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant to H.P.
Institutional Review Board Statement
Ethical review and approval were waived for this study, due to the animals already being deceased before they came into our possession.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to their being used for manuscripts in preparation.
Acknowledgments
We thank the staff and volunteers of all the wildlife rehabilitation centers and window-kill monitoring programs for providing Rock Pigeons (listed geographically from west to east): Wildlife Rescue Association of BC (Burnaby, British Columbia), Calgary Wildlife Rehabilitation Society (Calgary, Alberta), WILDNorth - Northern Alberta Wildlife Rehabilitation and Rescue (Edmonton, Alberta), Living Sky Wildlife Rehabilitation (Saskatoon, Saskatchewan), Wildlife Haven Rehabilitation Centre (Ile des Chênes, Manitoba), Prairie & Wildlife Rehabilitation Centre (Winnipeg, Manitoba), Sandy Pines Wildlife Centre (Napanee, Ontario), Fatal Light Awareness Program (FLAP, Toronto, Ontario), Hope for Wildlife (Seaforth, Nova Scotia) and Cobequid Wildlife Rehabilitation Centre (Brookfield, Nova Scotia). We also thank. Ildiko Szabo from the University of British Columbia Beaty Biodiversity Museum and Terry Galloway from the University of Manitoba for helping to acquire Rock Pigeons. Sergey Mironov (Zoological Institute of the Russian Academy of Sciences) kindly helped us with taxonomic issues associated with the feather mite genera Diplaegidia and Pterophagus. We finally thank Jeffrey Skevington, from the Canadian National Collection of Insects, Arachnids and Nematodes who generously looked up collection records for hippoboscid flies in Canada.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Torchin, M.E.; Mitchell, C.E. Parasites, pathogens, and invasions by plants and animals. Front. Ecol. Environ. 2004, 2, 183–190. [Google Scholar] [CrossRef]
- Lymbery, A.J.; Morine, M.; Kanani, H.G.; Beatty, S.J.; Morgan, D.L. Co-invaders: The effects of alien parasites on native hosts. Int. J. Parasitol. Parasites Wildl. 2014, 3, 171–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheath, D.J.; Williams, C.F.; Reading, A.J.; Robert Britton, J. Parasites of non-native freshwater fishes introduced into England and Wales suggest enemy release and parasite acquisition. Biol. Invasions 2015, 17, 2235–2246. [Google Scholar] [CrossRef] [Green Version]
- Macleod, C.J.; Paterson, A.M.; Tompkins, D.M.; Duncan, R.P. Parasites lost-do invaders miss the boat or drown on arrival? Ecol. Lett. 2010, 13, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.M.; Palma, R.L.; Gray, R.D. Drowning on arrival, missing the boat, and x-events: How likely are sorting events? In Tangled Trees: Phylogeny, Cospeciation and Coevolution; Page, R.D., Ed.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 287–309. [Google Scholar]
- Torchin, M.E.; Lafferty, K.D.; Dobson, A.P.; Mckenzie, V.J.; Kuris, A.M. Introduced species and their missing parasites. Lett. Nat. 2003, 421, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R. Evolutionary Ecology of Parasites, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Dabert, J. Feather mites (Astigmata; Pterolichoidea, Analgoidea) and birds as models for cophylogenetic studies. Phytophaga 2004, 14, 409–424. [Google Scholar]
- Dabert, J.; Dabert, M.; Mironov, S.V. Phylogeny of feather mite subfamily avenzoariinae (Acari: Analgoidea: Avenzoariidae) inferred from combined analyses of molecular and morphological data. Mol. Phylogenet. Evol. 2001, 20, 124–135. [Google Scholar] [CrossRef]
- Dabert, J. The feather mite family Syringobiidae Trouessart, 1896 (Acari, Astigmata, Pterolichoidea). 2. Phylogeny and host-parasite evolutionary relationships. Acta Parasitol. 2003, 48, 185–233. [Google Scholar]
- Banks, J.C.; Palma, R.L.; Paterson, A.M. Cophylogenetic relationships between penguins and their chewing lice. J. Evol. Biol. 2006, 19, 156–166. [Google Scholar] [CrossRef]
- Johnson, K.P.; Kennedy, M.; McCracken, K.G. Reinterpreting the origins of flamingo lice: Cospeciation or host-switching? Biol. Lett. 2006, 2, 275–278. [Google Scholar] [CrossRef]
- Gustafsson, D.R.; Zou, F. Gallancyra gen. nov. (Phthiraptera: Ischnocera), with an overview of the geographical distribution of chewing lice parasitizing chicken. Eur. J. Taxon. 2020, 685, 1–36. [Google Scholar] [CrossRef]
- Price, R.D.; Hellenthal, R.A.; Palma, R.L.; Johnson, K.P.; Clayton, D.H. The Chewing Lice: World Checklist and Biological Overview; Illinois Natural History Survey Special Publication: Champaign, IL, USA, 2003. [Google Scholar]
- Gaud, J.; Atyeo, W.T. Feather mites of the world (Acarina, Astigmata): The supraspecific taxa. Mus. R. Afr. Cent. Ann. Sci. Zool. 1996, 277, 1–193. [Google Scholar]
- Marshall, A. The Ecology Ectoparasitic Insects; Academic Press: London, UK, 1981. [Google Scholar]
- Proctor, H.C. Feather mites (Acari: Astigmata): Ecology, behavior, and evolution. Annu. Rev. Entomol. 2003, 48, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha Amaral, H.L.; Bergmann, F.B.; Silveira, T.; dos Santos, P.R.S.; Krüger, R.F. Pseudolynchia canariensis (Diptera: Hippoboscidae): Distribution pattern and phoretic association with skin mites and chewing lice of Columba livia (Aves: Columbidae). J. Nat. Hist. 2013, 47, 2927–2936. [Google Scholar] [CrossRef]
- Goodwin, D. Pigeons and Doves of the World, 2nd ed.; British Museum (Natural History): London, UK, 1977. [Google Scholar]
- EBird eBird: An online database of bird distribution and abundance (web application). eBird, Cornell Lab of Ornithology: Ithaca, NY, USA. Available online: http://www.ebird.org. (accessed on 10 December 2019).
- Schorger, A.W. Introduction of the domestic pigeon. Auk 1952, 69, 462–463. [Google Scholar] [CrossRef]
- Galloway, T.D.; Lamb, R.J. Infestation parameters for chewing lice (Phthiraptera: Amblycera, Ischnocera) infesting owls (Aves: Strigidae, Tytonidae) in Manitoba, Canada. Can. Entomol. 2019, 151, 608–620. [Google Scholar] [CrossRef]
- Lamb, R.J.; Galloway, T.D. Abundance and stability of populations of chewing lice (Phthiraptera: Amblycera and Ischnocera) infesting two species of woodpeckers (Aves: Piciformes: Picidae). Can. Entomol. 2018, 150, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Galloway, T.D. Phthiraptera of Canada. Zookeys 2019, 819, 301–310. [Google Scholar] [CrossRef]
- Knee, W.; Proctor, H.; Galloway, T. Survey of nasal mites (Rhinonyssidae, Ereynetidae, and Turbinoptidae) associated with birds in Alberta and Manitoba, Canada. Can. Entomol. 2008, 140, 364–379. [Google Scholar] [CrossRef]
- Mitchell, W.R. Observations on poultry diseases in Ontario. Ontario Vet. Coll. Rep. 1953, 1952, 78–79. [Google Scholar]
- Moyer, B.R.; Drown, D.M.; Clayton, D.H. Low humidity reduces ectoparasite pressure: Implications for host life history evolution. Oikos 2002, 97, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, C.M.; Valera, F.; Barbosa, A.; Moreno, E. Thriving in an arid environment: High prevalence of avian lice in low humidity conditions. Écoscience 2007, 12, 241–249. [Google Scholar] [CrossRef]
- Dowling, D.K.; Richardson, D.S.; Blaakmeer, K.; Komdeur, J. Feather mite loads influenced by salt exposure, age and reproductive stage in the Seychelles Warbler Acrocephalus sechellensis. J. Avian Biol. 2001, 32, 364–369. [Google Scholar] [CrossRef]
- Meléndez, L.; Laiolo, P.; Mironov, S.; García, M.; Magaña, O.; Jovani, R. Climate-driven variation in the intensity of a host-symbiont animal interaction along a broad elevation gradient. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabiyi, T.P. Association between duration of humid season and geographical distribution patterns of different species of chewing lice (Mallophaga: Insecta) infesting domestic chickens in Nigeria. J. Parasitol. 1996, 82, 1034–1036. [Google Scholar] [CrossRef]
- Shorthouse, D.P. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. Available online: www.simplemappr.net (accessed on 25 September 2020).
- Grossi, A.A.; Sharanowski, B.J.; Galloway, T.D. Anatoecus species (Phthiraptera: Philopteridae) from Anseriformes in North America and taxonomic status of Anatoecus dentatus and Anatoecus icterodes. Can. Entomol. 2014, 146, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Petryszak, A.; Rościszewska, M.; Bonczar, Z.; Szwalec, R. Observations on the vitality of lice from dead pigeons. Wiad. Parazytol. 1996, 42, 337–347. (In Polish) [Google Scholar]
- Gaud, J. Acariens Sarcoptiformes plumicoles parasites sur les oiseaux Lariformes et Columbiformes d’Afrique. Ann. Mus. R. Afr. Cent. Sér. 8 Sci. Zool. 1976, 214, 1–101. (In French) [Google Scholar]
- Smiley, R.L. A review of the family Cheyletiellidae (Acarina). Ann. Entomol. Soc. Am. 1970, 63, 1056–1078. [Google Scholar] [CrossRef]
- Bochkov, A.V.; Galloway, T.D. Parasitic cheyletoid mites (Acari: Cheyletoidea) associated with passeriform birds (Aves: Passeriformes) in Canada. Can. J. Zool. 2001, 79, 2014–2028. [Google Scholar] [CrossRef]
- Emerson, K.C. A new species of Mallophaga from the Pigeon. Florida Entomol. 1957, 40, 63–64. [Google Scholar] [CrossRef]
- Galloway, T.D.; Palma, R.L. Serendipity with chewing lice (Phthiraptera: Menoponidae, Philopteridae) infesting rock pigeons and mourning doves (Aves: Columbiformes: Columbidae) in Manitoba, with new records for North America and Canada. Can. Entomol. 2008, 140, 208–218. [Google Scholar] [CrossRef]
- Adams, R.J.; Price, R.D.; Clayton, D.H. Taxonomic revision of Old World members of the feather louse genus Columbicola (Phthiraptera: Ischnocera), including descriptions of eight new species. J. Nat. Hist. 2005, 39, 3545–3618. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own term: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Peter, S.; et al. Vegan: Community Ecology Package. R package v 2.4–6 2018. Available online: https://cran.ism.ac.jp/web/packages/vegan/vegan.pdf (accessed on 1 September 2020).
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/. (accessed on 1 September 2020).
- Bland, M.; Altman, D.G. Multiple significance tests: The Bonferroni method. BMJ 1995, 310, 170. [Google Scholar] [CrossRef] [Green Version]
- Simon, S. A Modern Approach to Regression with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Ròzsa, L. Speciation patterns of ectoparasites and “straggling” lice. Int. J. Parasitol. 1993, 23, 859–864. [Google Scholar] [CrossRef]
- Kerness Whiteman, N.; Santiago-Alarcon, D.; Johnson, K.P.; Parker, P.G. Differences in straggling rates between two genera of dove lice (Insecta: Phthiraptera) reinforce population genetic and cophylogenetic patterns. Int. J. Parasitol. 2004, 34, 1113–1119. [Google Scholar] [CrossRef]
- Rayner, J.A. Parasites of wild birds in Quebec. Sci. Agric. 1932, 12, 307–309. [Google Scholar] [CrossRef]
- Bochkov, A.V.; O’Connor, B.M.; Klompen, H. A review of the mite subfamily Harpirhynchinae (Acariformes: Harpirhynchidae)—Parasites of New World birds (Aves: Neognathae). Zootaxa 2015, 4023, 1–130. [Google Scholar] [CrossRef] [Green Version]
- Banks, N. New Canadian mites. Proc. Entomol. Soc. Washingt. 1909, 11, 133–143. [Google Scholar]
- Hill, W.W.; Tuff, D.W. A review of the Mallophaga parasitizing the Columbiformes of North America North of Mexico. J. Kans. Entomol. Soc. 1978, 51, 307–327. [Google Scholar]
- Harbison, C.W.; Bush, S.E.; Malenke, J.R.; Clayton, D.H. Comparative transmission dynamics of competing parasite species. Ecology 2008, 89, 3186–3194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaud, J.; Petitot, M.L. Sacroptides plumicoles des oiseaux du Maroc. Ann. Parasitol. Hum. Comparée 1948, 23, 35–46. (In French) [Google Scholar] [CrossRef]
- Millthorpe, A.P.; Eves, N. Pigeon mites and human infestation. New Zeal. Entomol. 1971, 5, 90–92. [Google Scholar] [CrossRef]
- Oudemans, A.C. Description du Myialges anchora Sergent et Trouessart 1907 (Acarien). Ann. Parasitol. Hum. Comp. 1935, 13, 5–11. [Google Scholar] [CrossRef]
- Dubinin, V. Studies in the adaptation of ectoparasites. 2. Ecological adaptations of the mallophagans and feather mites. Parazitol. Sb. 1947, 9, 191–222. (In Russian) [Google Scholar]
- Gaud, J.; Barré, N. Les genres Pterophagoides et Byersalges (Falculiferidae, Pterolichoidea) parasites plumicoles des Columbidae. Acarologia 1988, 29, 63–71. (In French) [Google Scholar]
- Dubinin, V. Fauna of the USSR—Arachnida. Part 3. Family Pterolichidae 1. Zool. Inst. Acad. Sci. USSR 1956, 6, 1–363. (In Russian) [Google Scholar]
- Cerny, V. Die parasitischen Arthropoden der synanthropen Taubenvögel einer Grossstadt. Angew. Parasitol. 1970, 11, 244–248. [Google Scholar]
- Naz, S.; Sychra, O.; Rizvi, S.A. New records and a new species of chewing lice (Phthiraptera, Amblycera, Ischnocera) found on Columbidae (Columbiformes) in Pakistan. Zookeys 2012, 174, 79–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knab, F. Four European Diptera established in North America. Insecutor Inscitiae Menstruus 1916, 4, 1–4. [Google Scholar]
- Martin, M. Life history and habits of the Pigeon Louse (Colimbicola columbae L.). Can. Entomol. 1934, 66, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Clayton, D.H.; Moyer, B.R.; Bush, S.E.; Jones, T.G.; Gardiner, D.W.; Rhodes, B.B.; Goller, F. Adaptive significance of avian beak morphology for ectoparasite control. Proc. Biol. Sci. 2005, 272, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doña, J.; Proctor, H.; Serrano, D.; Johnson, K.P.; Oploo, A.O.; Huguet-Tapia, J.C.; Ascunce, M.S.; Jovani, R. Feather mites play a role in cleaning host feathers: New insights from DNA metabarcoding and microscopy. Mol. Ecol. 2019, 28, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.C.; Murray, M.D. The distribution of Mallophaga on the domestic pigeon (Columba livia). Int. J. Parasitol. 1971, 1, 21–29. [Google Scholar] [CrossRef]
- Galloway, T.D.; Lamb, R.J. Seasonal population dynamics of four species of chewing lice (Phthiraptera: Menoponidae, Philopteridae) on feral pigeons (Aves: Columbiformes: Columbidae). Can. Entomol. 2015, 147, 712–722. [Google Scholar] [CrossRef] [Green Version]
- Pap, P.L.; Vágási, C.I.; Osváth, G.; Mureşan, C.; Barta, Z. Seasonality in the uropygial gland size and feather mite abundance in house sparrows Passer domesticus: Natural covariation and an experiment. J. Avian Biol. 2010, 41, 653–661. [Google Scholar] [CrossRef]
- Blanco, G.; Frías, O. Symbiotic feather mites synchronize dispersal and population growth with host sociality and migratory disposition. Ecography 2001, 24, 113–120. [Google Scholar] [CrossRef]
- Hamstra, T.L.; Badyaev, A.V. Comprehensive investigation of ectoparasite community and abundance across life history stages of avian host. J. Zool. 2009, 278, 91–99. [Google Scholar] [CrossRef]
- Brown, N.S. A survey of the arthropod parasites of pigeons (Columba livia) in Boston. J. Parasitol. 1971, 57, 1379–1380. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Anser Rizvi, S.; Sychra, O. The high rate of infestation of chewing lice (Phthiraptera) in Rock Pigeons (Columba livia Gmelin 1789) in Pakistan. Trop. Zool. 2010, 23, 21–28. [Google Scholar]
- Forond, A.P.; Valladares, B.; Rivera-Medin, A.J.A.; Figueruel, O.E.; Abre, U.N.; Casanov, A.J.C. Parasites of Columba livia (Aves: Columbiformes) in Tenerife (Canary Islands) and their role in the conservation biology of the Laurel pigeons. Parasite 2004, 11, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adang, K.L.; Oniye, S.J.; Ezealor, A.U.; Abdu, P.A.; Ajanusi, O.J. Ectoparasites of domestic pigeon (Columba livia domestica, Linnaeus) in Zaria, Nigeria. Res. J. Parasitol. 2008, 3, 79–84. [Google Scholar] [CrossRef]
- Bochkov, A.; Williams, G.; Proctor, H. First record of Picobia zumpti (Acari: Syringophilidae) from quills of the rock pigeon in North America and description of the male. Belg. J. Entomol. 2006, 7, 121–127. [Google Scholar]
- Kaszewska, K.; Skoracki, M.; Hromada, M. A review of the quill mites of the genus Gunabopicobia Skoracki and Hromada (Acariformes: Prostigmata: Syringophilidae) associated with birds of the order Columbiformes. Int. J. Acarol. 2018, 44, 288–299. [Google Scholar] [CrossRef]
Figure 1.
Map of Canada with marked sampling locations. Created using SimpleMappr [32].
Figure 1.
Map of Canada with marked sampling locations. Created using SimpleMappr [32].

Figure 2.
Prevalence of feather dwelling mites: (A) Falculifer rostratus, (B) Pterophagus columbae, and (C) Diplaegidia columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of C. livia examined from each location.
Figure 2.
Prevalence of feather dwelling mites: (A) Falculifer rostratus, (B) Pterophagus columbae, and (C) Diplaegidia columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of C. livia examined from each location.

Figure 3.
Prevalence of the non-feather dwelling mites: (A) Ornithocheyletia hallae, (B) Harpyrhynchoides gallowayi, (C) Harpyrhynchoides columbae, (D) Tinaminyssus melloi, and (E) Tinaminyssus columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of C. livia examined from each location.
Figure 3.
Prevalence of the non-feather dwelling mites: (A) Ornithocheyletia hallae, (B) Harpyrhynchoides gallowayi, (C) Harpyrhynchoides columbae, (D) Tinaminyssus melloi, and (E) Tinaminyssus columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of C. livia examined from each location.

Figure 4.
Prevalence of feather lice: (A) Columbicola columbae, (B) Campanulotes compar, (C) Coloceras tovornikae, (D) Hohorstiella lata, and (E) Bonomiella columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of C. livia examined from each location.
Figure 4.
Prevalence of feather lice: (A) Columbicola columbae, (B) Campanulotes compar, (C) Coloceras tovornikae, (D) Hohorstiella lata, and (E) Bonomiella columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of C. livia examined from each location.

Figure 5.
Mean intensity (red bar), each gray point represents the number of lice on an infested host. Feather lice: (A) Columbicola columbae, (B) Campanulotes compar, (C) Coloceras tovornikae, (D) Hohorstiella lata, and (E) Bonomiella columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of infested C. livia from each location.
Figure 5.
Mean intensity (red bar), each gray point represents the number of lice on an infested host. Feather lice: (A) Columbicola columbae, (B) Campanulotes compar, (C) Coloceras tovornikae, (D) Hohorstiella lata, and (E) Bonomiella columbae infesting Columba livia sampled from seven locations across Canada, 95% confidence intervals. Sample sizes refer to number of infested C. livia from each location.

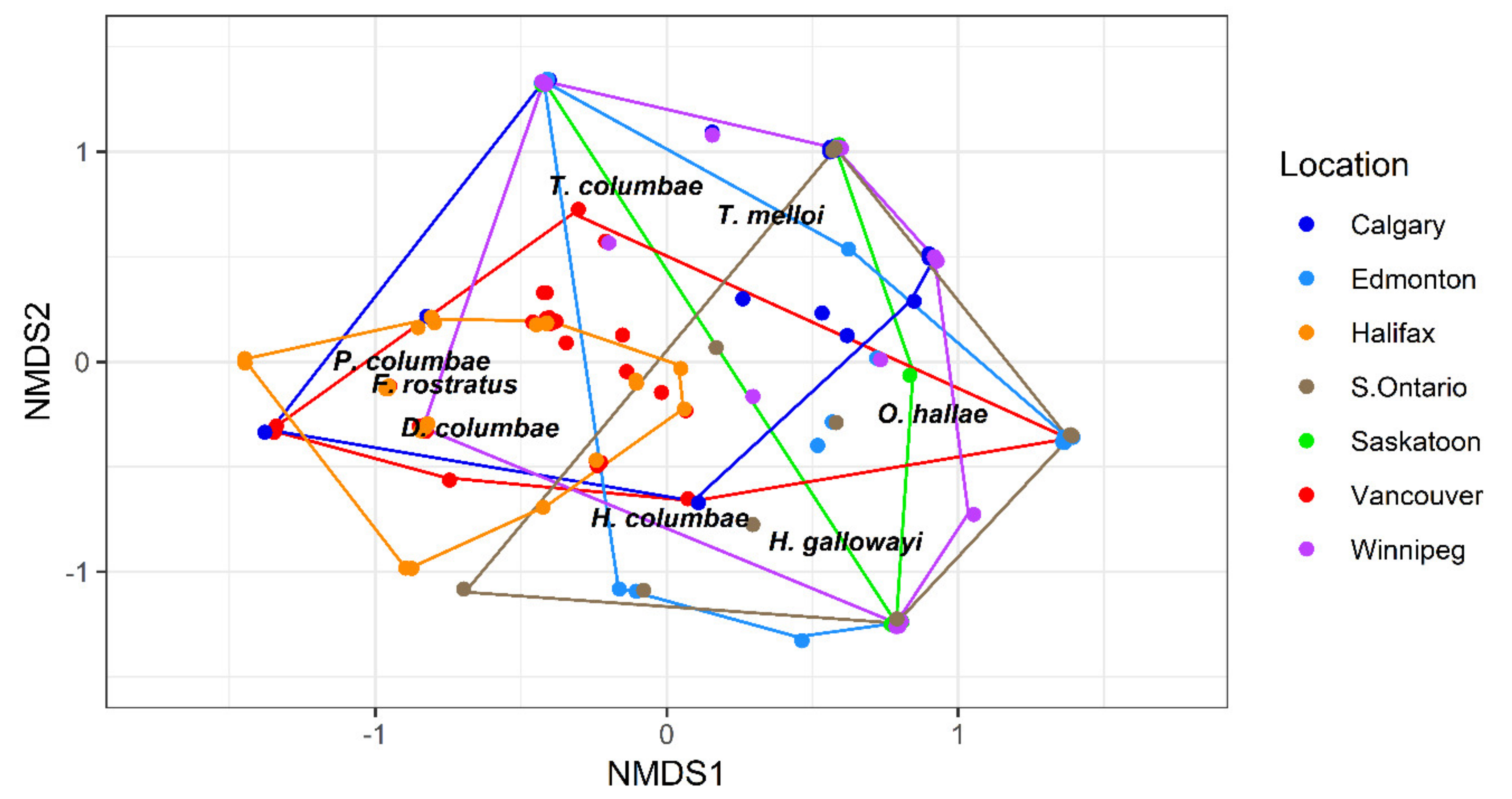
Figure 6.
Non-metric multidimensional scaling plot of mite assemblages (presence-absence data) infesting Columba livia (n = 162) sampled from seven locations across Canada. Locations of mite names indicate the direction of their vectors. Stress = 0.1180, points have been jittered (width = 0.02, height = 0.02) to minimize visual overlap. Vancouver mite assemblages are significantly different from every other location except Halifax and Halifax was different from every other location other than Vancouver.
Figure 6.
Non-metric multidimensional scaling plot of mite assemblages (presence-absence data) infesting Columba livia (n = 162) sampled from seven locations across Canada. Locations of mite names indicate the direction of their vectors. Stress = 0.1180, points have been jittered (width = 0.02, height = 0.02) to minimize visual overlap. Vancouver mite assemblages are significantly different from every other location except Halifax and Halifax was different from every other location other than Vancouver.

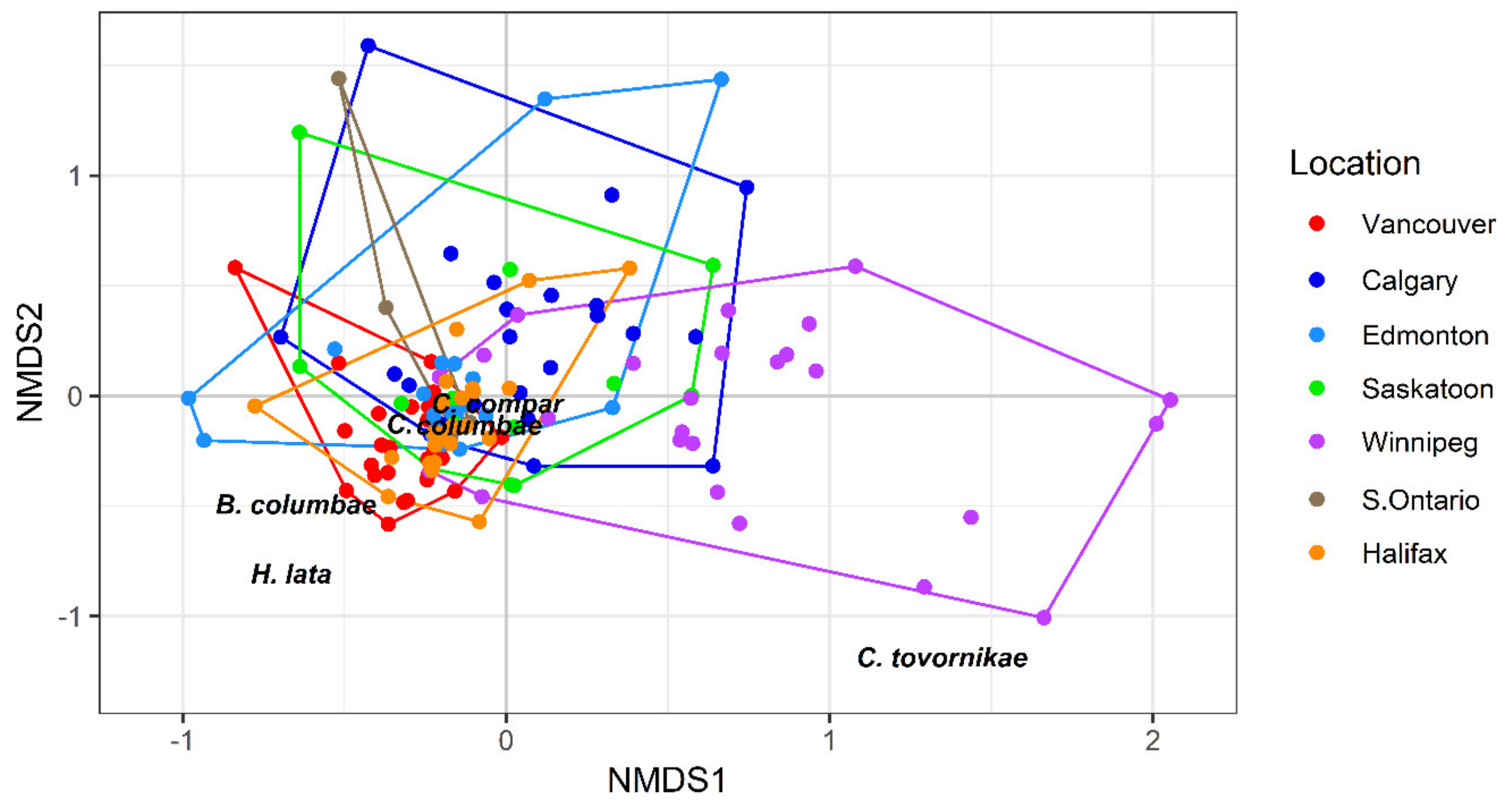
Figure 7.
Non-metric multidimensional scaling plot of louse assemblages (log [x + 1] abundance data) infesting Columba livia (n = 162) sampled from seven locations across Canada. Locations of louse names indicate the direction of their vectors. Stress = 0.1189. Locations that have significantly different louse assemblages are Vancouver and Calgary, Vancouver and Winnipeg, Edmonton and Winnipeg, and Winnipeg and Halifax.
Figure 7.
Non-metric multidimensional scaling plot of louse assemblages (log [x + 1] abundance data) infesting Columba livia (n = 162) sampled from seven locations across Canada. Locations of louse names indicate the direction of their vectors. Stress = 0.1189. Locations that have significantly different louse assemblages are Vancouver and Calgary, Vancouver and Winnipeg, Edmonton and Winnipeg, and Winnipeg and Halifax.

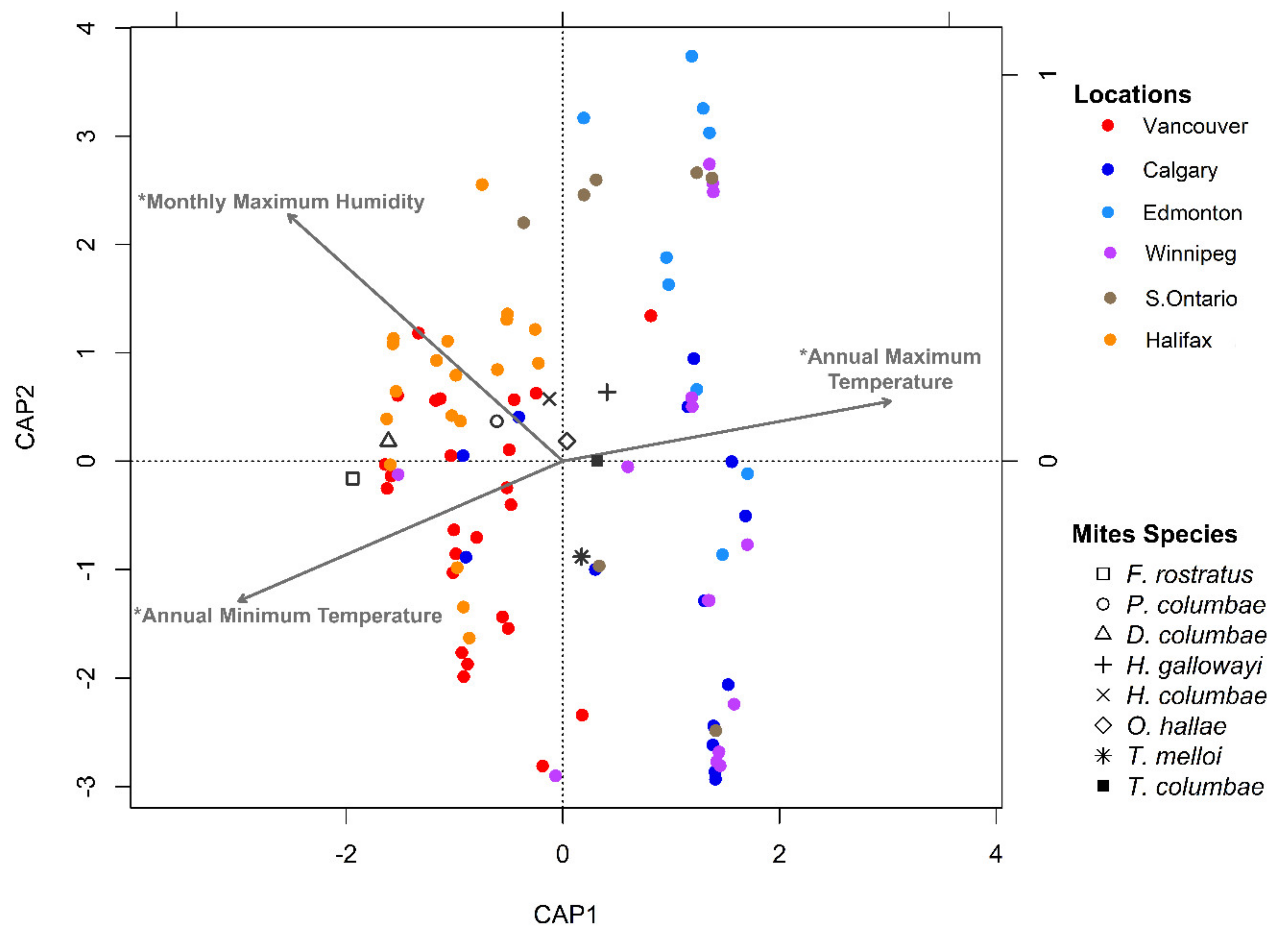
Figure 8.
Distance-based redundancy analysis plot of mite assemblages (presence-absence data) from Columba livia sampled from six locations across Canada. Vectors are meteorological variables chosen through forward selection. An asterisk (*) indicates a p-value < 0.05 after correction for multiple comparisons.
Figure 8.
Distance-based redundancy analysis plot of mite assemblages (presence-absence data) from Columba livia sampled from six locations across Canada. Vectors are meteorological variables chosen through forward selection. An asterisk (*) indicates a p-value < 0.05 after correction for multiple comparisons.

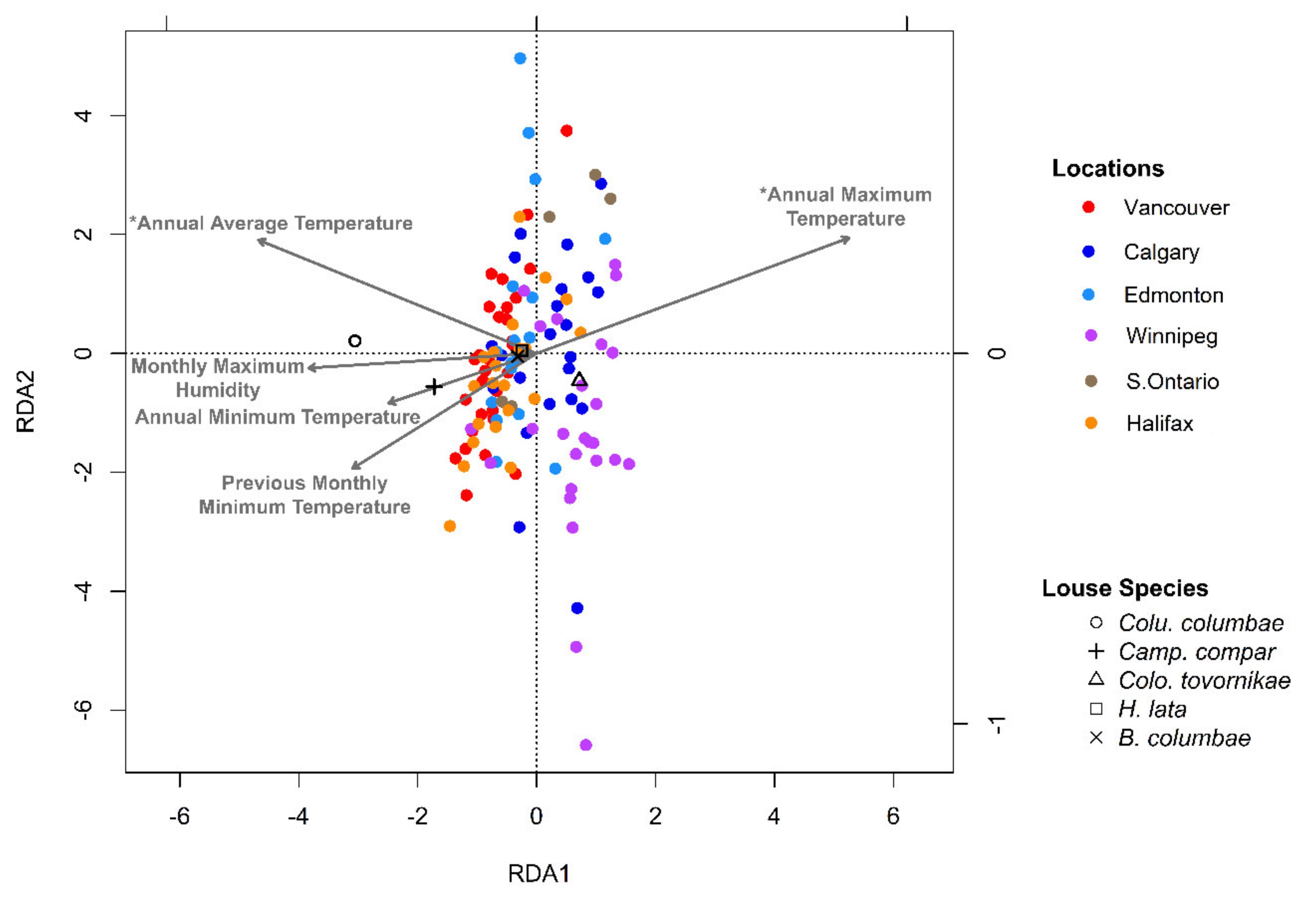
Figure 9.
Redundancy analysis plot of louse assemblages (abundance data log (x + 1) transformed) from Columba livia sampled from six locations across Canada. Vectors are meteorological variables chosen through forward selection. An asterisk (*) indicates a p-value < 0.05 after correction for multiple comparisons.
Figure 9.
Redundancy analysis plot of louse assemblages (abundance data log (x + 1) transformed) from Columba livia sampled from six locations across Canada. Vectors are meteorological variables chosen through forward selection. An asterisk (*) indicates a p-value < 0.05 after correction for multiple comparisons.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Prevalence of mites found on Columba livia collected from seven locations across Canada. 95% confidence intervals in parentheses.
Table 1.
Prevalence of mites found on Columba livia collected from seven locations across Canada. 95% confidence intervals in parentheses.
| Vancouver (n = 30) | Calgary (n = 30) | Edmonton (n = 23) | Saskatoon (n = 14) | Winnipeg (n = 30) | S. Ontario (n = 11) | Halifax (n = 24) | |
|---|---|---|---|---|---|---|---|
| Falculiferidae | |||||||
| Falculifer rostratus | 93.3 (78.7–98.8) | 10.0 (2.8–26.3) | 0 | 0 | 6.7 (1.2–21.3) | 9.1 (0.5–40.4) | 83.3 (62.8–94.1) |
| Pterophagus columbae | 13.3 (4.7–29.8) | 0 | 0 | 0 | 0 | 0 | 41.7 (23.4–62.8) |
| Analgidae | |||||||
| Diplaegidia columbae | 76.7 (58.4–88.8) | 6.7 (1.2–21.3) | 8.7 (1.6–27.8) | 0 | 6.7 (1.2–21.3) | 27.3 (7.9–59.6) | 83.3 (62.8–94.1) |
| Harpirhynchidae | |||||||
| Harpyrhynchoides gallowayi | 10.0 (2.8–26.3) | 6.7 (1.2–21.3) | 30.4 (14.5–52.2) | 21.4 (6.1–50.0) | 20.0 (9–38) | 45.5 (20.0–73.5) | 20.8 (8.6–41.5) |
| Harpyrhynchoides columbae | 10.0 (2.8–26.3) | 0 | 17.3 (6.2–38.9) | 0 | 0 | 8.2 (3.3–50.0) | 16.7 (5.9–37.2) |
| Cheyletidae | |||||||
| Ornithocheyletia hallae | 26.7 (13.1–44.9) | 20.0 (9.1–38.2) | 26.1 (12.0–47.8) | 7.1 (0.4–31.7) | 10.0 (2.8–23.6) | 36.4 (13.5–66.7) | 12.5 (3.5–31.0) |
| Rhinonyssidae | |||||||
| Tinaminyssus melloi | 46.7 (29.8–65.2) | 36.7 21.4–55.1) | 8.7 (1.6–27.8) | 14.2 (2.6–42.6) | 33.3 (17.7–51.7) | 27.3 (7.9–59.6) | 29.2 (13.9–50.0) |
| Tinaminyssus columbae | 16.7 (6.8–34.7) | 20.0 (9.1–38.2) | 21.7 (9.0–43.3) | 7.1 (0.4–31.7) | 10.0 (2.8–23.4) | 9.1 (0.5–40.4) | 16.7 (5.9–37.2) |
Table 2.
Prevalence of lice found on Columba livia collected from seven locations across Canada. 95% confidence intervals in parentheses.
Table 2.
Prevalence of lice found on Columba livia collected from seven locations across Canada. 95% confidence intervals in parentheses.
| Vancouver (n = 30) | Calgary (n = 30) | Edmonton (n = 23) | Saskatoon (n = 14) | Winnipeg (n = 30) | S. Ontario (n = 11) | Halifax (n = 24) | |
|---|---|---|---|---|---|---|---|
| Philopteridae | |||||||
| Columbicola columbae | 100 (88.9–100) | 73.3 (55.1–86.9) | 91.3 (72.2–98.4) | 85.7 (57.4–97.4) | 33.3 (17.7–51.7) | 90.9 (59.6–99.5) | 100 (86.1–100) |
| Campanulotes compar | 96.7 (82.3–99.8) | 76.7 (58.4–88.8) | 91.3 (72.2–98.4) | 92.9 (68.3–99.6) | 66.7 (48.3–82.3) | 81.1 (50.0–96.7) | 100.0 (86.1–100) |
| Coloceras tovornikae | 3.3 (0.2–17.7) | 16.7 (6.8–34.8) | 0 | 28.6 (10.4–57.4) | 56.7 (38.2–73.7) | 0 | 4.2 (0.2–20.4) |
| Menoponidae | |||||||
| Hohorstiella lata | 26.7 (13.1–44.9) | 0 | 4.3 (0.2–21.3) | 0 | 10.0 (2.8–26.3) | 27.3 (7.9–59.6) | 4.2 (0.2–20.4) |
| Bonomiella columbae | 26.7 (13.1–44.9) | 0 | 0 | 7.1 (0.4–31.7) | 0 | 0 | 12.5 (3–31.0) |
Table 3.
Mean intensity of lice found on Columba livia collected from seven locations across Canada. 95% confidence intervals in parentheses and ranges in brackets.
Table 3.
Mean intensity of lice found on Columba livia collected from seven locations across Canada. 95% confidence intervals in parentheses and ranges in brackets.
| Vancouver | Calgary | Edmonton | Saskatoon | Winnipeg | S. Ontario | Halifax | |
|---|---|---|---|---|---|---|---|
| Philopteridae | |||||||
|
Columbicola columbae | 326.1 (246.7–427.7) [22–841] | 64.2 (33.9–112.0) [1–2026] | 136.2 (95.1–219.1) [1–640] | 173.6 (97.5–287.7) [4–542] | 107.5 (20.3–280.6) [1–9] | 53.8 (16.5–96.3) [2–118] | 303.1 (171.0–696.3) [1–2669] |
|
Campanulotes compar | 282.0 (186.0–441.6) [6–1428] | 47.8 (32.2–71.9) [1–194] | 104.8 (67.3–175.1) [1–524] | 110.1 (66.5–199.2) ^ [2–318] | 102.8 (61.7–183.6)[4–568] | 122.7× [4–195] | 239.2 (148.5–400.3) [1–2669] |
| Coloceras tovornikae | 1.0 * | 18.6 (2.2–44.2) [1–65] | - | 15.0 (4.3–25.5) [4–28] | 54.2 (22.3–117.6) ^ [1–404] | - | 5.0 * |
| Menoponidae | |||||||
| Hohorstiella lata | 14.5 (6.4–23.8) [1–31] | - | 182.0 * | - | 3.7 × [1–5] | - | 20.0 * |
|
Bonomiella columbae | 7.3 (4.5–12.4) [2–20] | - | - | 3.0 * | - | - | 6.3 (5.0–7.3) [5–8] |
* only one host infested; × sample size too small to calculate confidence interval; ^ 90% confidence interval, due to small sample size.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grossi, A.; Proctor, H. Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada. Diversity 2021, 13, 9. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010009
AMA Style
Grossi A, Proctor H. Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada. Diversity. 2021; 13(1):9. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010009
Chicago/Turabian StyleGrossi, Alexandra, and Heather Proctor. 2021. "Variation in Ectosymbiont Assemblages Associated with Rock Pigeons (Columba livia) from Coast to Coast in Canada" Diversity 13, no. 1: 9. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010009
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.