Antimicrobial Cocktail Combining Specific Peptide Extracts from Native Probiotic Bacteria Hamper Adulteration of Ready-to-Eat Mango Wedges


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbiological Diagnostic Quality of Mango Fruit and Fresh-Cut Wedges Purchased from the Local Market
2.2. Antibiotic Susceptibility Testing
2.3. Lactic Acid Bacterial Strains Growth Conditions, and Preparation of Cell-Free Supernatant (CFS) and Precipitated Peptides (PP)
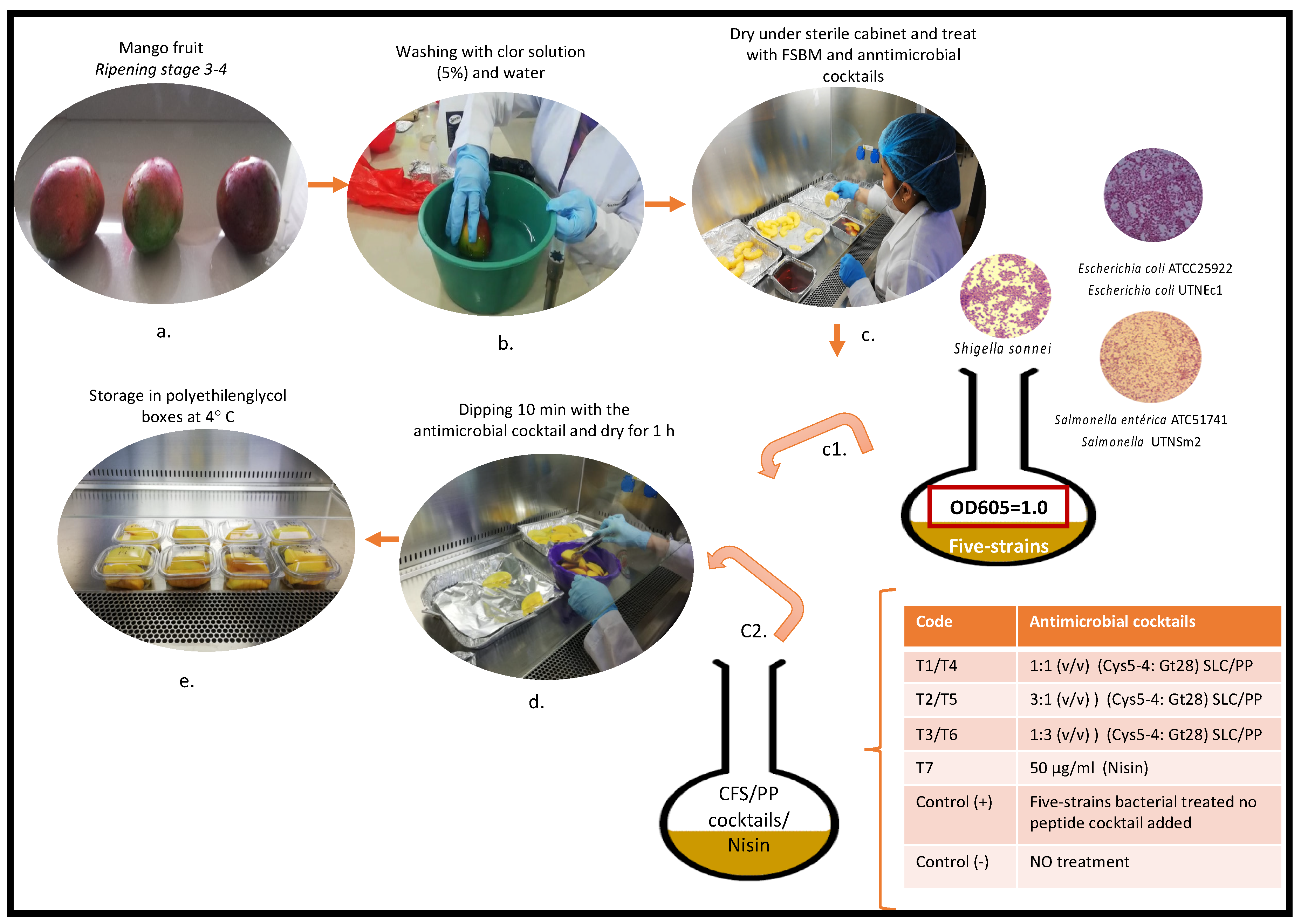
2.4. Composition of Five-Strain Bacterial Mixture (FSBM)
2.5. Establishment of Antimicrobial Cocktails Composition
2.6. Inoculation of Fresh-Cut Mango Wedges with FSBM
2.7. Treatment of Mango Wedges with Antimicrobial Cocktails
2.8. Determination of the Total Cell Counts in Mango Wedges during Storage
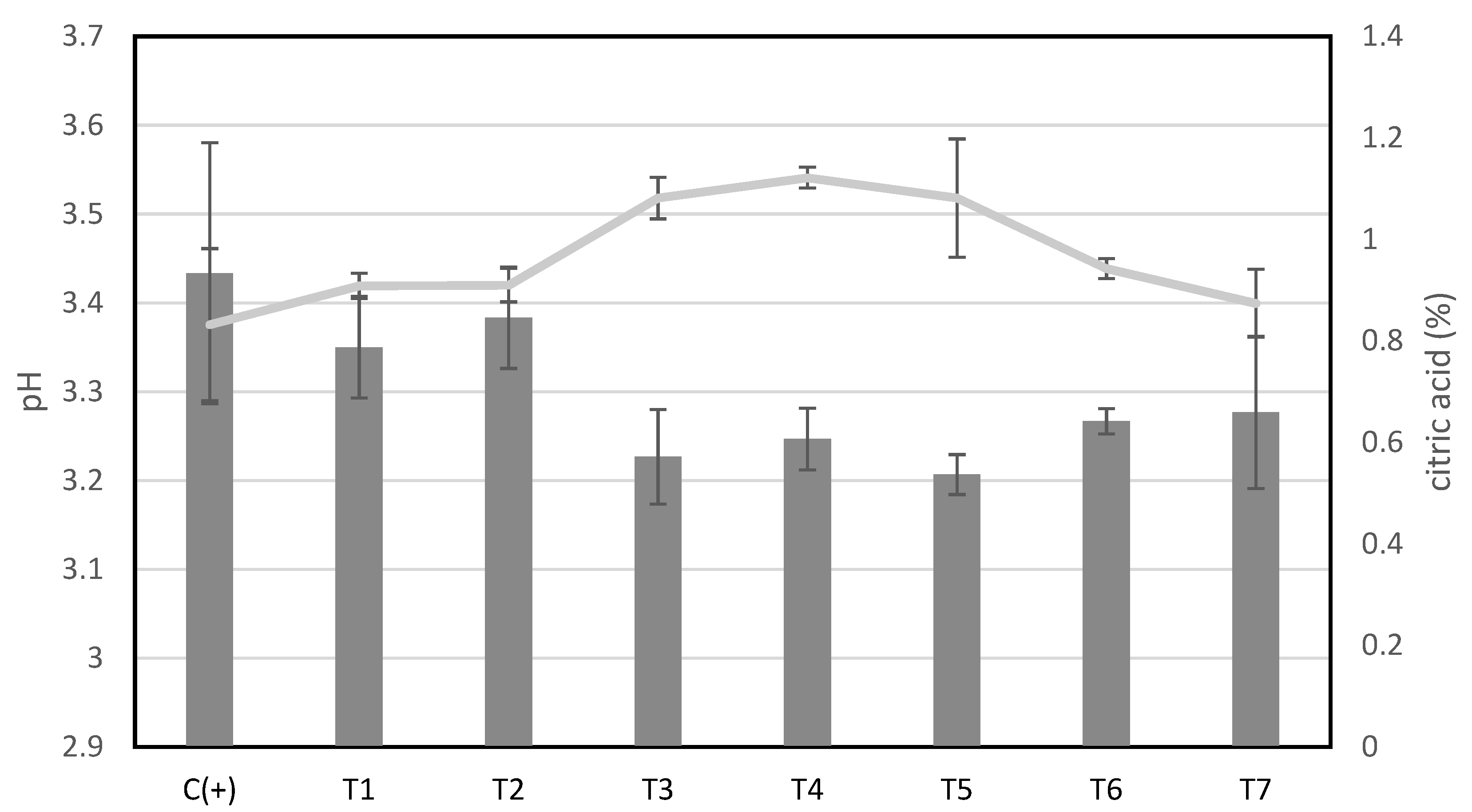
2.9. Monitoring the pH and Acidity of Mango Wedges during Storage
2.10. Cytoplasmic Membrane Permeabilization
2.11. Cell Membrane Integrity Assay
2.12. Statistical Analysis
3. Results
3.1. Mango Wedges Sold by Mobile Vendors Hold Antibiotic-Resistant Microorganisms
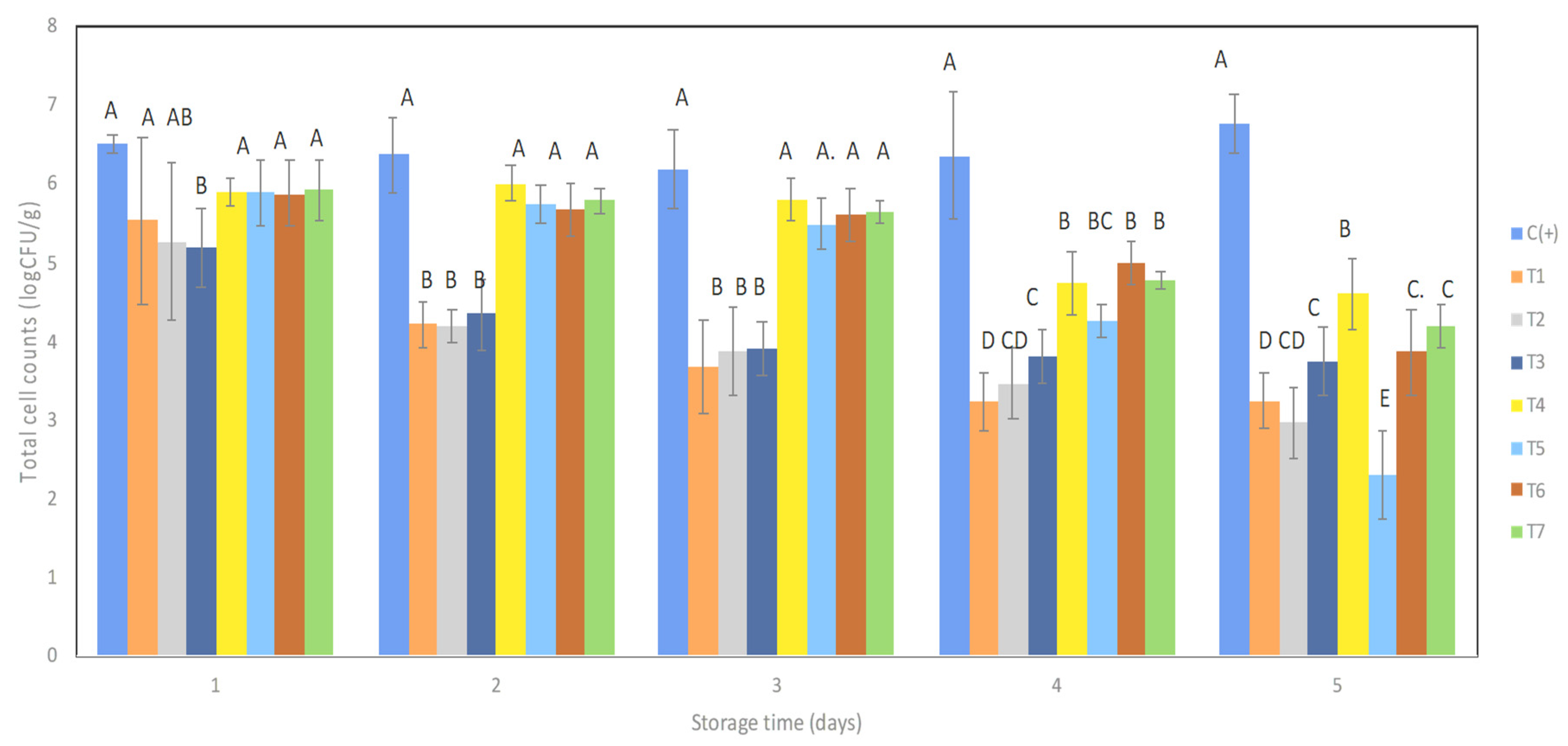
3.2. Antimicrobial Cocktails Diminished the Total FSBM Viability in Mango Wedges during Storage with Refrigeration
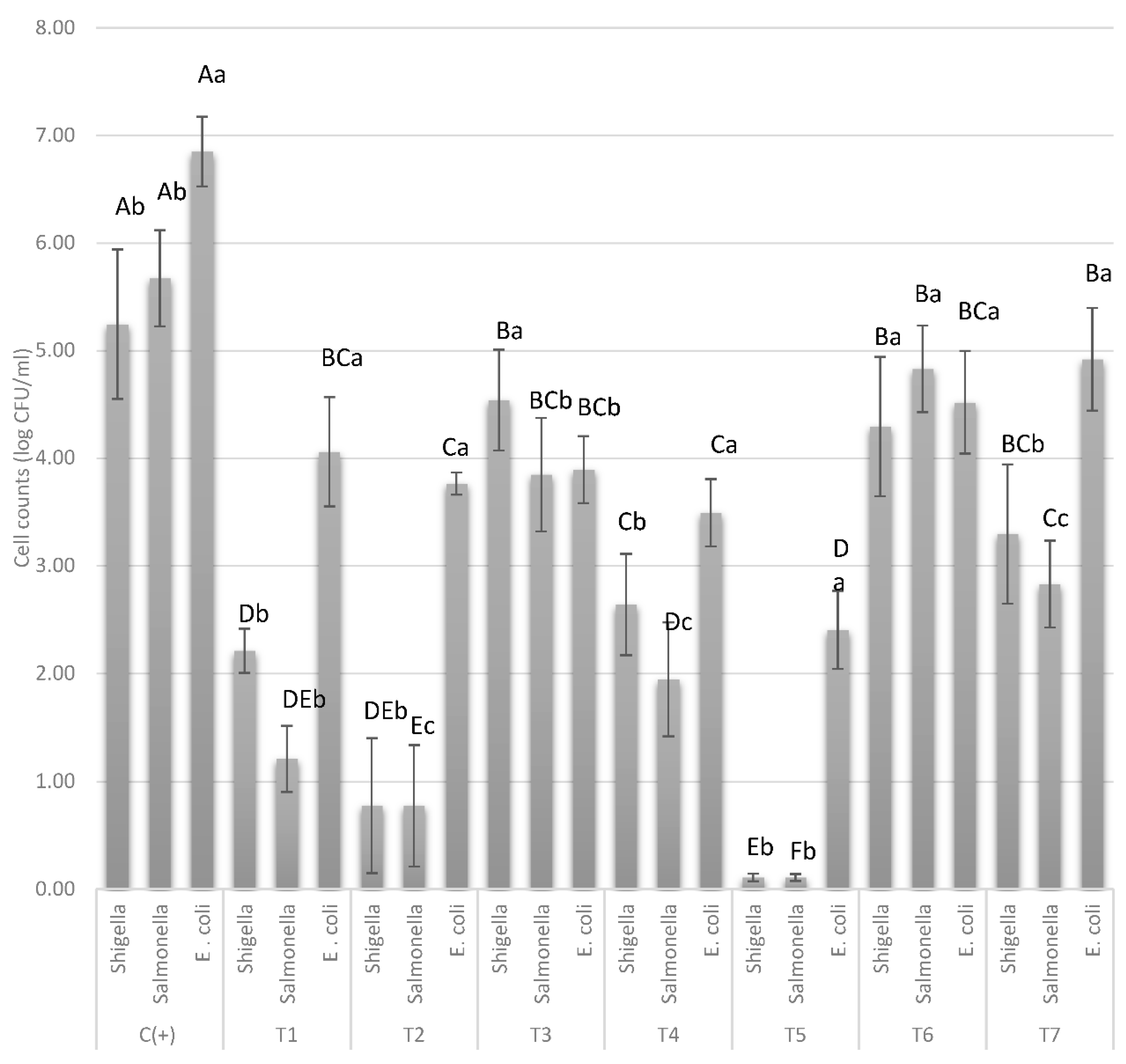
3.3. The Antimicrobial Cocktails T2 and T5 Inhibit Salmonella/Shigella and to Some Extent E. coli Growth in Mango Wedges
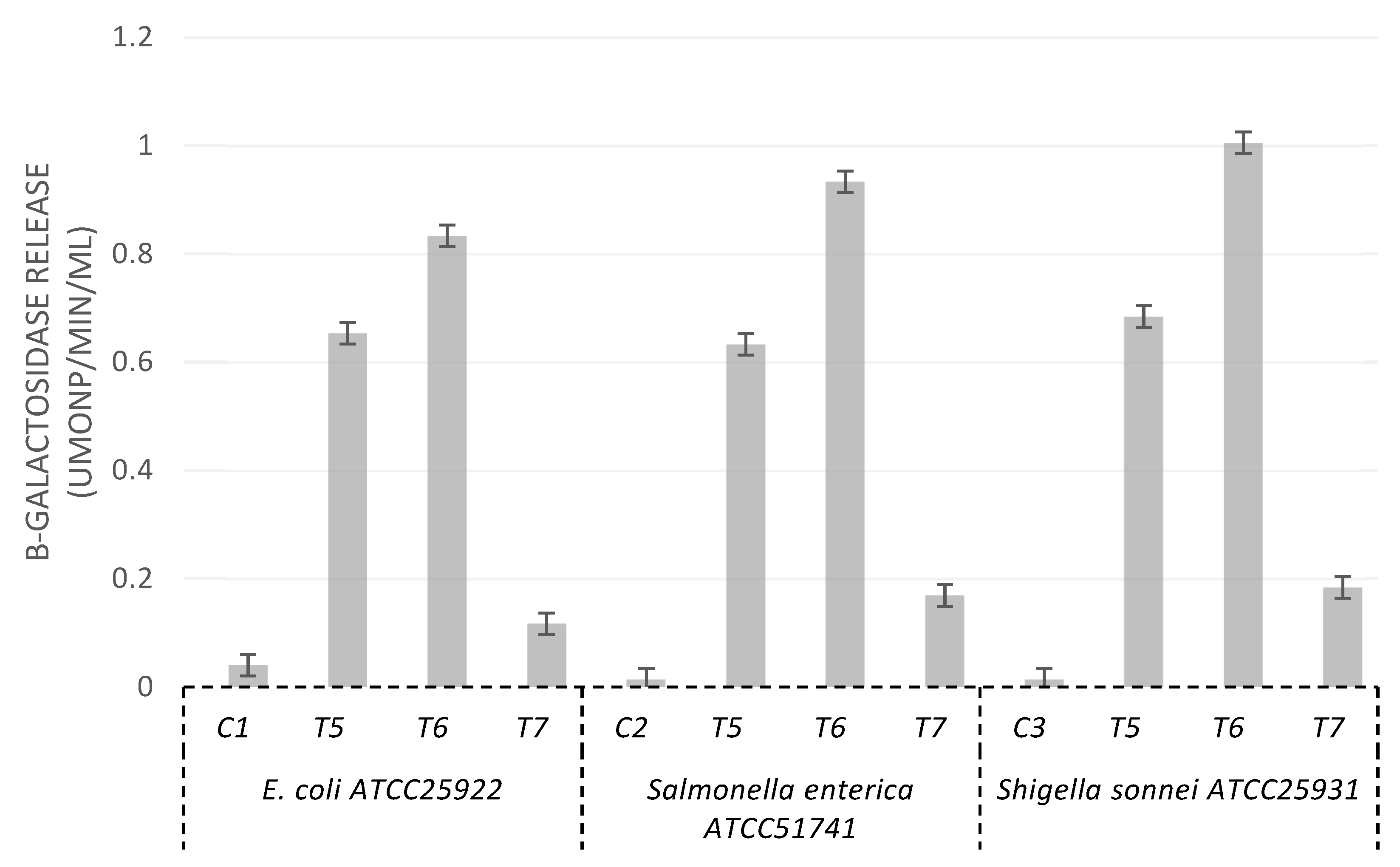
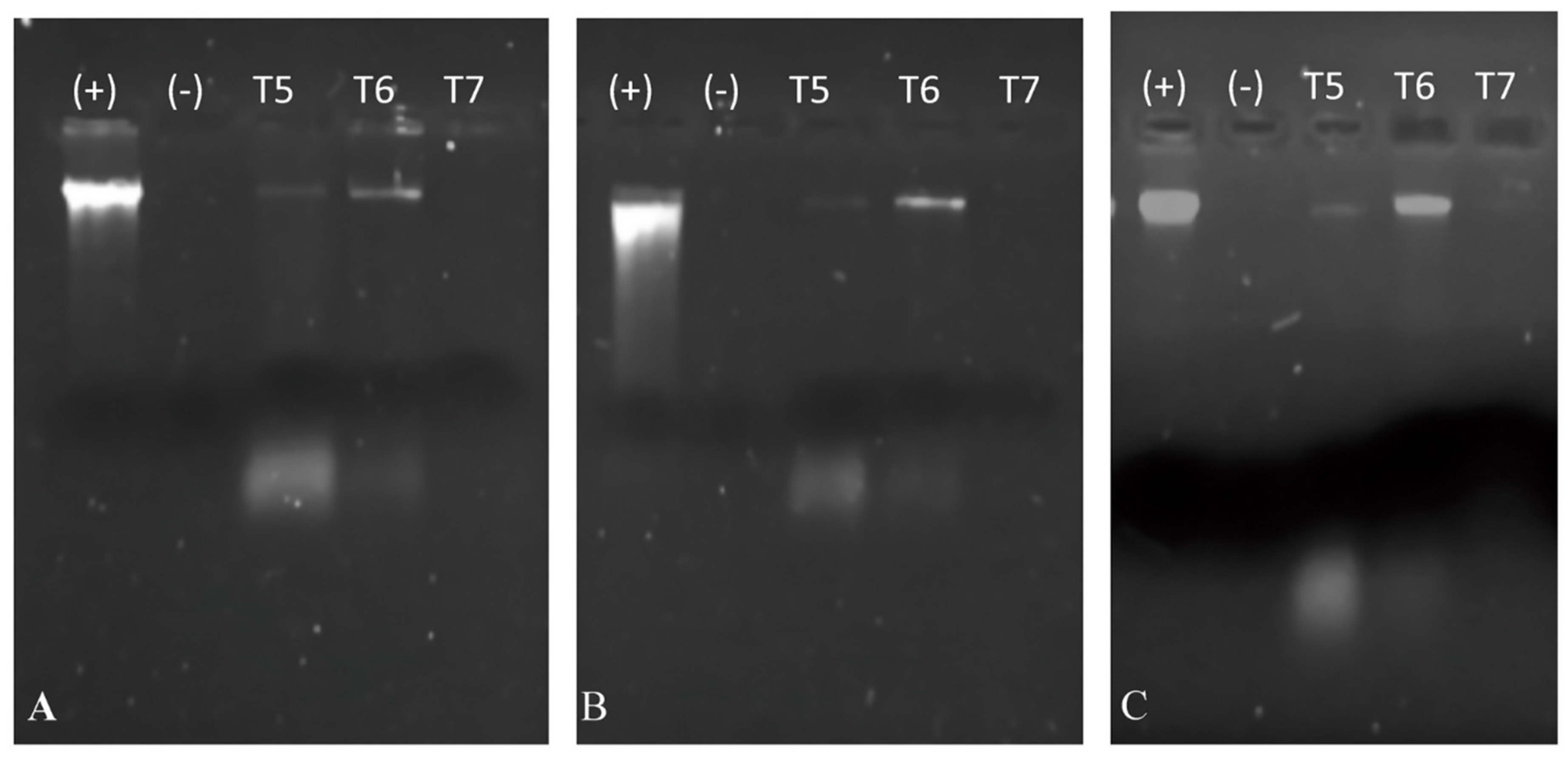
3.4. Cytoplasmic Membrane Permeation and Alteration of Cell Membrane Integrity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bernal, A.C.; Maldonado, M.E.; Urango, L.A.; Franco, M.C.; Rojano, B.A. Sugar mango (Mangifera indica), variety from Colombia: Antioxidant, nutritional and sensorial characteristics. Rev. Chil. Nutr. 2014, 41. [Google Scholar] [CrossRef] [Green Version]
- Guererro, G.H. La produccion del mango ecuatoriano. Perspectiva 2018. Available online: https://perspectiva.ide.edu.ec/investiga/2018/06/26/la-produccion-del-mango-ecuatoriano/ (accessed on 3 June 2020).
- Arequipa, X.B. Diseño de un plan de marketing para la exportación del mango ecuatoriano hacia el mercado español. Bibdigital 2010. Available online: http://bibdigital.epn.edu.ec/handle/15000/1612 (accessed on 8 June 2020).
- Jadán Solís, K.P.; Najarro Quintero, R. Análisis de las políticas públicas agrícolas y la seguridad alimentaria de la provincia de Los Ríos. Rev. Cientif. Ecocien. 2019, 6, 1–16. [Google Scholar] [CrossRef]
- Ministerio De Salud Pública. MSP lidera la primera campaña de capacitación en manipulación de alimentos. Quito: Ministerio de Salud Pública. 2014. Available online: http://www.salud.gob.ec/msp-lidera-la-primera-campana-de-capacitacion-en-manipulacion-de-alimentos/ (accessed on 10 October 2020).
- Nazuka, E.; Inatsu, Y.; Kayasaki, S.; Miyamaru, M. The investigation of bacterial contamination in fresh-cut fruits, and the behavior of Escherichia coli O157: H7 and Salmonella enteritidis on fruit cubes. Jap. J. Food Microbio. 2004, 21, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Branquinho Bordini, M.E.; Asturiano Ristori, C.; Jakabi, M.; Scala Gelli, D. Incidence, internalization and behavior of Salmonella in mangoes, var. Tommy Atkins. Food Control. 2007, 18, 1002–1007. [Google Scholar] [CrossRef]
- Alegre, I.; Abadias, M.; Anguera, M.; Oliveira, M.; Viñas, I. Factors affecting growth of foodborne pathogens on minimally processed apples. Food Microbiol. 2010, 27, 70–76. [Google Scholar] [CrossRef]
- CDC (Center for Disease Control and Prevention). Outbreak of Salmonella Poona Infections Linked to Imported Cucumbers (Final Update). 2016. Available online: https://www.cdc.gov/salmonella/poona-09-15/index.html (accessed on 11 October 2020).
- CDC (Center for Disease Control and Prevention). Outbreak of Listeriosis Linked to Whole Cantaloupes from Jensen Farms, Colorando (Final Update). 2012. Available online: https://www.cdc.gov/listeria/outbreaks/cantaloupes-jensen-farms/index.html (accessed on 6 October 2020).
- CDC (Center for Disease Control and Prevention). Outbreak of Salmonella Infections Linked to Pre-Cut Melons. 2019. Available online: https://www.cdc.gov/salmonella/carrau-04-19/index.html (accessed on 11 October 2020).
- CDC (Center for Disease Control and Prevention). Outbreak of Salmonella Infections Linked to Cut Fruit. 2020. Available online: https://www.cdc.gov/salmonella/javiana-12-19/index.html (accessed on 11 October 2020).
- CDC (Center for Disease Control and Prevention). Outbreak of Salmonella Infections Linked to Gavi Brand Whole, Fresh Papayas. 2020. Available online: https://www.cdc.gov/salmonella/uganda-06-19/index.html (accessed on 11 October 2020).
- Rojas-Graü, M.A.; Soliva-Fortuny, R.; Martín-Belloso, O. Edible coatings to incorporate active ingredients to fresh-cut fruits: A review. Trends Food Sci. Technol. 2009, 20, 438–447. [Google Scholar] [CrossRef]
- Oliveira, M.; Abadias, M.; Colás-Medà, P.; Usall, J.; Viñas, I. Biopreservative methods to control the growth of foodborne pathogens on fresh-cut lettuce. Int. J. Food Microbiol. 2015, 214, 4–11. [Google Scholar] [CrossRef]
- Rai, M.; Pandit, R.; Gaikwad, S.; Kövics, G. Antimicrobial peptides as natural bio-preservative to enhance the shelf-life of food. J. Food Sci. Technol. 2016, 53, 3381–3394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Meza, J.E.; Ochoa-Zarzosa, A.; Barboza-Corona, J.E.; Bideshi, D.K. Antimicrobial peptides: Current and potential applications in biomedical therapies. Biomed Res. Int. 2015, 367243. [Google Scholar] [CrossRef] [PubMed]
- Upendra, R.S.; Khandelwal, P.; Jana, K.; Ajay Kumar, N.; Gayathri Devi, M.; Stephaney, M.L. Bacteriocin production from indigenous strains of lactic acid bacteria isolated from selected fermented food sources. Int. J. Pharma. Res. Health Sci. 2016, 4, 982–990. Available online: www.pharmahealthsciences.net (accessed on 20 October 2020).
- Saravanakumar, K.; Sathiyaseelan, A.; Mariadoss, A.V.A.; Chelliah, R.; Hu, X.; Oh, D.H.; Wang, M.H. Lactobacillus rhamnosusGG and Biochemical Agents Enrich the Shelf Life of Fresh-Cut Bell Pepper (Capsicum annuum L. var. grossum (L.) Sendt). Foods 2020, 9, 1252. [Google Scholar] [CrossRef] [PubMed]
- Tumbarski, Y.; Nikolova, R.; Petkova, N.; Ivanov, I.; Lante, A. Biopreservation of fresh strawberries by carboxymethyl cellulose edible coatings enriched with a bacteriocin from Bacillus methylotrophicus BM47. Food Technol. Biotechnol. 2019, 57, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenea, G.N.; Pozo Delgado, T. Antimicrobial peptides from Lactobacillus plantarum UTNGt2 prevent harmful bacteria growth on fresh tomatoes. J. Microbiol. Biotechnol. 2019, 29, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N.; Olmedo, D.; Ortega, C. Peptide-based formulation from lactic acid bacteria Impairs the pathogen growth in Ananas comosus (Pineapple). Coatings 2020, 10, 457. [Google Scholar] [CrossRef]
- Tenea, G.N.; Hurtado, P.; Ortega, C. Inhibitory effect of substances produced by native Lactococcus lactis strains of tropical fruits towards food pathogens. Prev. Nutr. Food Sci. 2018, 23, 260–268. [Google Scholar] [CrossRef] [PubMed]
- NTE INEN 1529-15:2009. Control Microbiológico de los Alimentos. Toma, Envío y Preparación de Muestras Para el Análisis Microbiológico. Instituto Ecuatoriano de Normalización (INEN), 14. Available online: https://archive.org/stream/ec.nte.1529.15.1996/ec.nte.1529.15.1996_djvu.txt (accessed on 10 July 2020).
- NTE INEN 1529-8: 2015.Norma. Técnica. Ecuatoriana. Control Microbiológico de los Alimentos. Detección y Recuento de Escherichia coli Presuntiva por la Técnica del Número más Probable. pp. 1–13. Available online: https://archive.org/stream/ec.nte.1529.8.1990#mode/2up (accessed on 10 July 2020).
- García Martínez, R.; López Jiménez, A.; Saucedo Veloz, C.; Salazar-García, S.; Suárez Espinosa, J. Maduración y calidad de frutos de mango ‘Kent’ con tres niveles de fertilización. Rev. Mex. Cienc. Agric. 2015, 6, 665–678. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-09342015000400001&lng=es&tlng=es (accessed on 13 October 2020).
- ISO, 6579: 2002 (E). Microbiology-General Guidance on Methods for the Detection of Salmonella, 4th ed.; International Organization for Standardization: Geneve, Switzerland, 2002; Available online: https://www.iso.org/standard/29315.html (accessed on 10 July 2020).
- Tenea, G.N.; Guana, J.M. Inhibitory substances produced by native Lactobacillus plantarum UTNUTNCys5-4 control microbial population growth in meat. J. Food Qual. 2019, a9516981. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety authority (EFSA). Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar] [CrossRef]
- Tenea, G.N. Peptide extracts from native lactic acid bacteria generate ghost cells and spheroplasts upon interaction with salmonella enterica, as promising food antimicrobials. BioMed Res. Int. 2020, 6152356. [Google Scholar] [CrossRef]
- Alimi, B.A.; Workneh, T.S. Consumer awareness and willingness to pay for safety of street foods in developing countries: A review. Int. J. Consum. Stud. 2016, 40, 242–248. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic acid bacteria as antibacterial agents to extend the shelf life of fresh and minimally processed fruits and vegetables: Quality and safety aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Hansen, J.N.; Sandine, W.E. Nisin as a model food preservative. Crit. Rev. Food Sci. Nutr. 1994, 34, 69–93. [Google Scholar] [CrossRef]
- Artes-Hernandez, F.; Martinez-Hernandez, G.R.; Aguayo, E.; Gomes, P.A.; Artes, F. Fresh-Cut Fruit and Vegetables: Emerging Eco-friendly Techniques for Sanitation and Preserving Safety. In Postharvest Handling; Kahramanoglu, I., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- FAO (Food and Agricultural Organization of the United States). Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; FAO: Rome, Italy, 2016; Available online: http://www.fao.org/3/a-a0512e.pdf (accessed on 15 July 2020).
- Favaro, L.; Penna, A.L.B.; Todorov, S.D. Bacteriocinogenic LAB from cheeses–application in biopreservation? Trends Food Sci. Technol. 2015, 41, 37–48. [Google Scholar] [CrossRef]
- Arqués, J.L.; Rodríguez, E.; Nuñez, M.; Medina, M. Combined effect of reuterin and lactic acid bacteria bacteriocins on the inactivation of food-borne pathogens in milk. Food Control. 2011, 22, 457–461. [Google Scholar] [CrossRef]
- Bari, M.L.; Ukuku, D.O.; Kawasaki, T.; Inatsu, Y.; Isshiki, K.; Kawamoto, S. Combined efficacy of nisin and pediocin with sodium lactate, citric acid, phytic acid, and potassium sorbate and EDTA in reducing the Listeria monocytogenes population of inoculated fresh-cut produce. J. Food Prot. 2005, 68, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Okuku, D.O.; Fett, W.F. Effect of nisin in combination with EDTA, sodium lactate, and potassium sorbate for reducing Salmonella on whole and fresh-cut cantaloupe. J. Food Prot. 2004, 67, 2143–2150. [Google Scholar] [CrossRef]
- Ragazzo-Sanchez, J.A.; Sanchez-Prado, L.; Gutiérrez-Martínez, P.; Luna-Solano, G.; Gomez-Gil, B.; Calderon-Santoyo, M. Inhibition of Salmonella spp. isolated from mango using bacteriocin-like produced by lactobacilli, CyTA. J. Food. 2009, 7, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Penteado, A.L.; Eblen, B.S.; Miller, A.J. Evidence of Salmonella internalization into fresh mangos during simulated postharvest insect disinfestations procedures. J. Food Prot. 2004, 67, 181–184. [Google Scholar] [CrossRef]
- Qadri, O.S.; Yousuf, B.; Srivastava, A.K. Fresh-cut fruits and vegetables: Critical factors influencing microbiology and novel approaches to prevent microbial risks—A review. Cogent Food Agric. 2015, 1, 1121606. [Google Scholar] [CrossRef]
- Bezerra, A.C.D.; Reis, R.B.; Bastos, D.H.M. Microbiological quality of hamburgers sold in the streets of Cuiabá–MT, Brazil and vendor hygiene- awareness. Food Sci. Technol. 2010, 30, 520–524. [Google Scholar] [CrossRef] [Green Version]
- Lues, J.F.R.; Rasephei, M.R.; Venter, P.; Theron, M.M. Assessing food safety and associated food handling practices in street food vending. Int. J. Environm. Health Res. 2006, 16, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Vincenti, S.; Raponi, M.; Sezzatini, R.; Giubbini, G.; Laurenti, P. Enterobacteriaceae antibiotic resistance in ready-to-eat foods collected from hospital and community canteens: Analysis of Prevalence. J. Food Prot. 2018, 81, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, Potent Antimicrobial Peptides and the Fight against Multi Drug Resistant Species: Resistance Is Futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaneghah, A.M.; Hashemi, S.M.B.; Limbo, S. Antimicrobial agents and packaging systems in antimicrobial active food packaging: An overview of approaches and interactions. Food Bioprod. Process. 2018, 111, 1–19. [Google Scholar] [CrossRef]
- Bouttefroy, A.; Millière, J.B. Nisin-curvaticin 13 combinations for avoiding the regrowth of bacteriocin resistant cells of Listeria monocytogenes ATCC 15313. Int. J. Food Microbiol. 2000, 62, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus plantarum Strains as a Bio-Control Strategy against Food-Borne Pathogenic Microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.; Silva, S.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Gravel, J.; Paradis-Bleau, C.; Schmitzer, A.R. Adaptation of a bacterial membrane permeabilization assay for quantitative evaluation of benzalkonium chloride as a membrane-disrupting agent. Medchemcomm. 2017, 8, 1408–1413. [Google Scholar] [CrossRef]
- Helander, I.M.; Mattila-Sandholm, T. Permeability barrier of the gram-negative bacterial outer membrane with special reference to nisin. Inter. J. Food Microbiol. 2000, 60, 153–161. [Google Scholar] [CrossRef]
- Heesterbeek, D.A.C.; Martin, N.I.; Velthuizen, A.; Duijst, M.; Ruyken, M.; Wubbolts, R.; Rooijakkers, S.H.M.; Bardoel, B.W. Complement-dependent outer membrane perturbation sensitizes Gram-negative bacteria to Gram-positive specific antibiotics. Sci. Rep. 2019, 9, 3074. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mango Fruit Samples | Indicator Microorganisms Counts (CFU/g) | ||||||
|---|---|---|---|---|---|---|---|
| Total Aerobes | E. coli spp. | Salmonella spp. * | Shigella spp. | Enterobacter spp. | Yeasts | Molds | |
| PE2V1 | 1.00 × 103 | 3.00 × 102 | ND | ND | 4.00 × 102 | 1.40 × 103 | ND |
| PE3V1 | 1.30 × 102 | 6.00 × 102 | ND | ND | 1.25 × 103 | 3.80 × 101 | ND |
| PE4V1 | 1.80 × 103 | 3.00 × 102 | ND | ND | 1.43 × 104 | 4.00 × 102 | ND |
| PL2V1 | 1.00 × 101 | ND | ND | ND | ND | ND | ND |
| PL3V1 | 2.97 × 101 | 2.15 × 101 | ND | ND | 3.00 × 101 | 7.00 ×100 | ND |
| PL4V1 | 2.40 × 101 | 3.00 × 102 | ND | ND | 4.00 × 101 | 4.00 × 101 | ND |
| PE2V2 | 7.70 × 103 | 7.10 × 103 | 3.25 × 103 | 1.25 × 103 | 2.15 × 103 | 2.30 × 103 | 3.00 × 100 |
| PE3V2 | 3.08 × 104 | 2.40 × 103 | 5.80 × 103 | 2.25 × 103 | 1.70 × 104 | 5.40 × 103 | 2.00 × 102 |
| PE4V2 | 1.93 × 104 | 1.65 × 104 | 4.55 × 103 | 1.80 × 103 | 1.14 × 104 | 3.90 × 103 | 2.00 × 102 |
| PL2V2 | 6.00 × 102 | 5.00 × 102 | 7.50 × 102 | 4.00 × 102 | 4.00 × 102 | 1.00 × 102 | ND |
| PL3V2 | 2.40 × 103 | 3.20 × 103 | 9.00 × 102 | 5.00 × 102 | 8.75 × 103 | 1.40 × 102 | ND |
| PL4V2 | 8.10 × 103 | 4.64 × 103 | 1.20 × 102 | 1.70 × 103 | 1.10 × 103 | 1.30 × 102 | ND |
| Mango Wedges Sample Source | Indicator Microorganisms Counts (CFU/g) | |||||
|---|---|---|---|---|---|---|
| Total Aerobes | E. coli. spp. | Yeasts/Molds | Shigella spp. | Salmonella spp. * | Enterobacter spp. | |
| V3 | 1.30 × 105 | 7.00 × 102 | 6.30 × 103/ND | ND | ND | 1.00 × 102 |
| V4 | 1.68 × 106 | 1.74 × 104 | 3.03 × 102/ND | 2.55 × 103 | 2.00 × 101 | 1.16 × 106 |
| Breakpoints (CLSI, 2017) R/I/S (mm) | Antimicrobials/Antibiotic Class | % Resistance | ||||
|---|---|---|---|---|---|---|
| V3 | V4 | |||||
| Enterobacter spp. (n = 30) | E. coli spp. (n = 30) | Enterobacter spp. (n = 30) | E. coli spp. (n = 30) | Shigella spp./Salmonella spp. (n = 20/20) | ||
| ≤13/14–17/≥18 | Kanamycin 30/Aminoglycosides | 66.67 | 66.67 | 100 | 66.67 | 66.67/25 |
| ≤12/13–14/≥15 | Gentamycin 10/Aminoglycosides | 100 | 100 | 100 | 100 | 100/25 |
| ≤13/14–16/≥17 | Ampicillin 10/β-lactam-Lactamase inhibitor combinations | 100 | 100 | 100 | 100 | 33.33/100 |
| ≤13/14–17/≥18 | Amoxicillin 25/Penicillin like antibiotics | 100 | 100 | 100 | 100 | 100/100 |
| ≤11/12–14/≥15 | Tetracycline 30/Tetracycline | 66.67 | 66.67 | 66.67 | 33.33 | 33.33/0 |
| ≤16/17–19/≥20 | Cefuroxime 30/Cephalosporins | 66.67 | 33.33 | 100 | 100 | 33.33/100 |
| Antimicrobial Cocktails | % of Total Cell Counts Reduction | % of Shigella Counts Reduction | % of Salmonella Counts Reduction | % of E. coli Counts Reduction |
|---|---|---|---|---|
| T1 | 52.10 | 57.89 | 78.70 | 40.77 |
| T2 | 56.03 | 85.23 | 86.35 | 45.08 |
| T3 | 44.51 | 13.46 | 32.20 | 43.19 |
| T4 | 31.93 | 49.69 | 65.70 | 49.03 |
| T5 | 55.61 | 97.93 | 98.09 | 64.93 |
| T6 | 42.88 | 18.14 | 14.87 | 34.05 |
| T7 | 37.85 | 37.22 | 40.13 | 28.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tenea, G.N.; Olmedo, D. Antimicrobial Cocktail Combining Specific Peptide Extracts from Native Probiotic Bacteria Hamper Adulteration of Ready-to-Eat Mango Wedges. Appl. Sci. 2021, 11, 2246. https://0-doi-org.brum.beds.ac.uk/10.3390/app11052246
Tenea GN, Olmedo D. Antimicrobial Cocktail Combining Specific Peptide Extracts from Native Probiotic Bacteria Hamper Adulteration of Ready-to-Eat Mango Wedges. Applied Sciences. 2021; 11(5):2246. https://0-doi-org.brum.beds.ac.uk/10.3390/app11052246
Chicago/Turabian StyleTenea, Gabriela N., and Daniela Olmedo. 2021. "Antimicrobial Cocktail Combining Specific Peptide Extracts from Native Probiotic Bacteria Hamper Adulteration of Ready-to-Eat Mango Wedges" Applied Sciences 11, no. 5: 2246. https://0-doi-org.brum.beds.ac.uk/10.3390/app11052246