
Genome-Wide Association Study Identifies Two Loci for Stripe Rust Resistance in a Durum Wheat Panel from Iran

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Materials
2.2. Disease Evaluations
2.3. DNA Isolation and Genotyping Assay
2.4. GWAS Analysis
3. Results
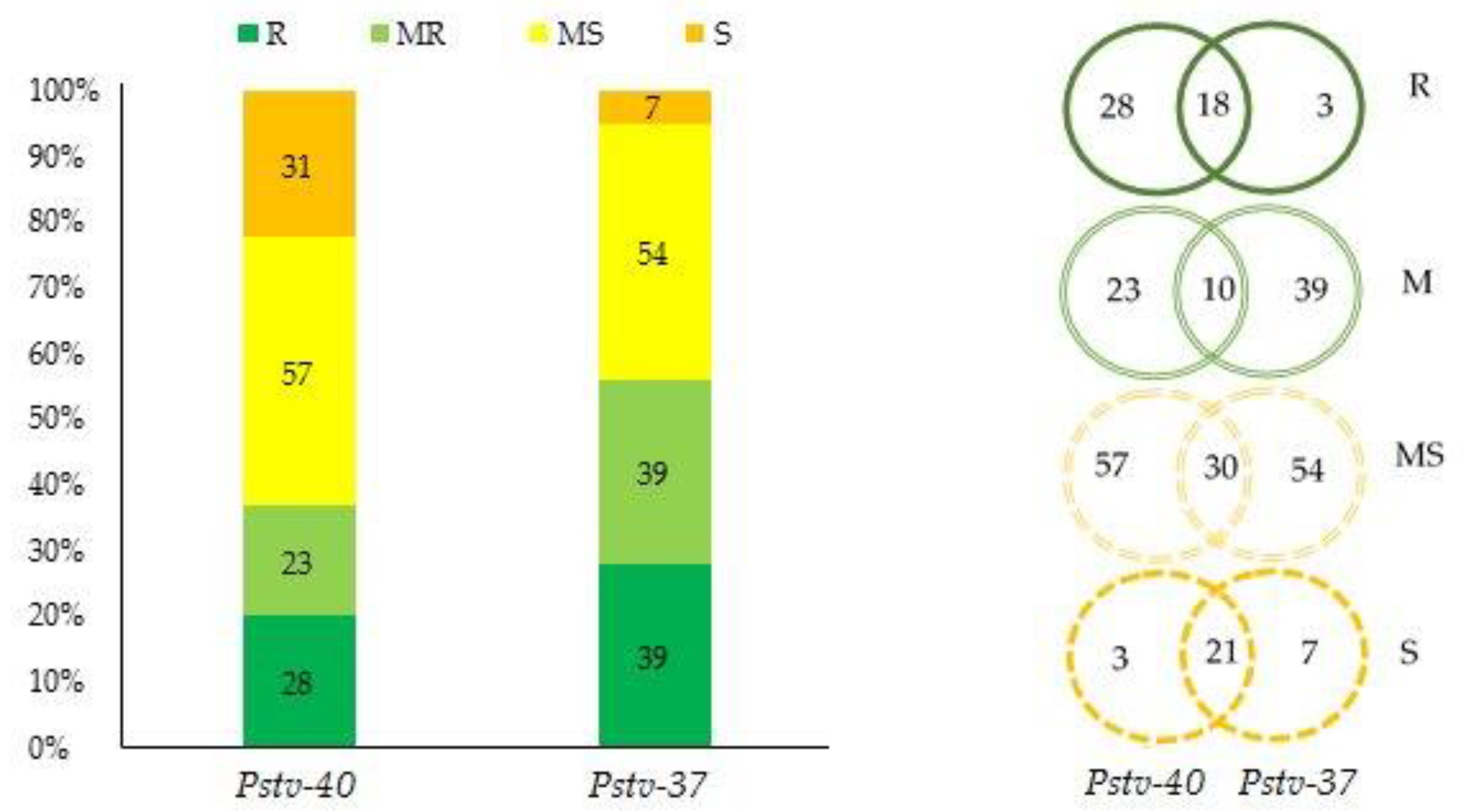
3.1. Response of Durum Wheat Genotypes to Stripe Rust (SR)
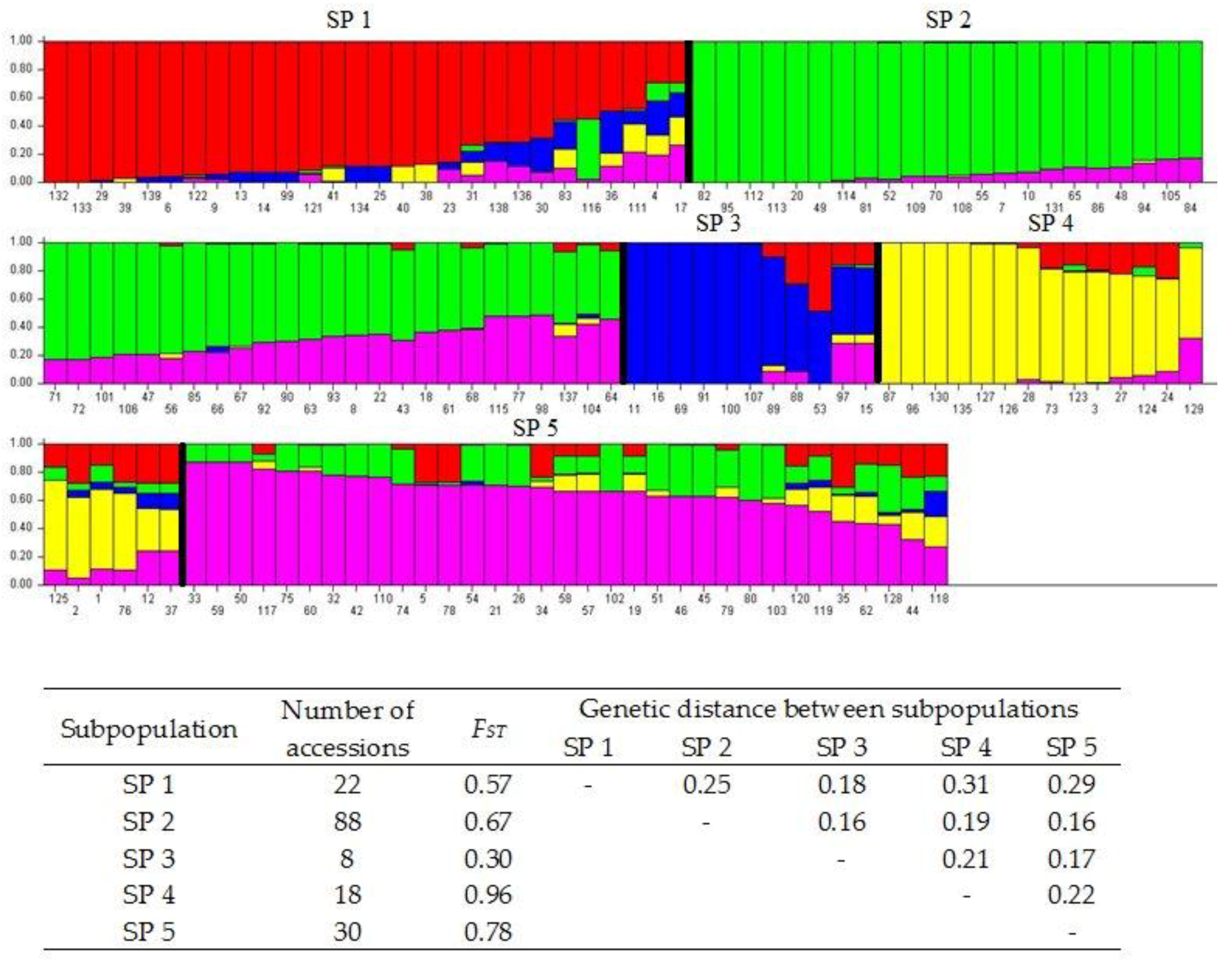
3.2. Population Structure
3.3. LD Decay Analysis and Markers Significantly Associated with Stripe Rust Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cassman, K.G.; Grassini, P. A global perspective on sustainable intensification research. Nat. Sustain. 2020, 3, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Beres, B.L.; Rahmani, E.; Clarke, J.M.; Grassini, P.; Pozniak, C.J.; Geddes, C.M.; Porker, K.D.; May, W.E.; Ransom, J.K. A Systematic Review of Durum Wheat: Enhancing Production Systems by Exploring Genotype, Environment, and Management (G × E × M) Synergies. Front. Plant Sci. 2020, 11, 568657. [Google Scholar] [CrossRef] [PubMed]
- International Grain Council (IGC). Available online: https://www.igc.int/en/default.aspx (accessed on 20 November 2021).
- Giunta, F.; Pruneddu, G.; Motzo, R. Grain yield and grain protein of old and modern durum wheat cultivars grown under different cropping systems. Field Crop. Res. 2019, 1, 107–120. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Etminan, A.; Abdelrahman, M.; Siddique, K.H.M.; Tran, L.S.P. Assessment of biochemical and physiological parameters of durum wheat genotypes at the seedling stage during polyethylene glycol-induced water stress. Plant Growth Regul. 2020, 92, 81–93. [Google Scholar] [CrossRef]
- Gull, A.; Lone, A.A.; Islam, N.U. Biotic and Abiotic Stresses in Plants. In Abiotic and Biotic Stress in Plants; IntechOpen: London, UK, 2019. [Google Scholar]
- Singla, R.K.; Guimarães, A.G.; Zengin, G. Editorial: Application of plant secondary metabolites to pain neuromodulation. Front. Pharmacol. 2020, 11, 623399. [Google Scholar] [CrossRef]
- Singla, P.; Bhardwaj, R.D.; Kaur, S.; Kaur, J. Stripe rust induced defence mechanisms in the leaves of contrasting barley genotypes (Hordeum vulgare L.) at the seedling stage. Protoplasma 2019, 257, 169–181. [Google Scholar] [CrossRef]
- Chen, Y.E.; Cui, J.M.; Su, Y.Q.; Yuan, S.; Yuan, M.; Zhang, H.Y. Influence of stripe rust infection on the photosynthetic characteristics and antioxidant system of susceptible and resistant wheat cultivars at the adult plant stage. Front. Plant Sci. 2015, 6, 779. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.M. Epidemiology and control of stripe rust (Puccinia striiformis f. sp. tritici) on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Liu, L.; Wang, M.N.; Feng, J.Y.; See, D.R.; Chao, S.M.; Chen, X.M. Combination of all-stage and high-temperature adult-plant resistance QTL confers high level, durable resistance to stripe rust in winter wheat cultivar Madsen. Theor. Appl. Genet. 2018, 131, 1835–1849. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Xia, X.C.; Catalogue of Gene Symbols for Wheat. Supplement. 2017. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf (accessed on 25 June 2019).
- Wang, M.; Chen, X. Stripe rust resistance. In Stripe Rust; Springer: Dordrecht, The Netherlands, 2017; pp. 353–558. [Google Scholar]
- Zaim, M.; El Hassouni, K.; Gamba, F.; Filali-Maltouf, A.; Belkadi, B.; Ayed, S.; Amri, A.; Nachit, M.; Taghouti, M.; Bassi, F. Wide crosses of durum wheat (Triticum durum Desf.) reveal good disease resistance, yield stability, and industrial quality across Mediterranean sites. Field Crops Res. 2017, 214, 219–227. [Google Scholar] [CrossRef]
- Li, H.; Bariana, H.; Singh, D.; Zhang, L.; Dillon, S.; Whan, A.; Urmil, B.; Ayliffe, M. A durum wheat adult plant stripe rust resistance QTL and its relationship with the bread wheat Yr80 locus. Theor. Appl. Genet. 2020, 133, 3049–3066. [Google Scholar] [CrossRef] [PubMed]
- Tam, V.; Patel, N.; Turcotte, M.; Bosse, Y.; Pare, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. 2019, 20, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Mwadzingeni, L.; Shimelis, H.; Rees, D.J.G.; Tsilo, T.J. Genome-wide association analysis of agronomic traits in wheat under drought stressed and non-stressed conditions. PLoS ONE 2017, 12, e0171692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Sun, L.; Hu, X.; Wang, Y.; Zhang, Y.; Nevo, E.; Peng, J.; Sun, D. Associations of canopy leaf traits with SNP markers in durum wheat (Triticum turgidum L. durum (Desf.)). PLoS ONE 2018, 13, e0206226. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, S.; Reynolds, M.P.; Sansaloni, C. Genome-wide association analyses identify QTL hotspots for yield and component traits in durum wheat grown under yield potential, drought, and heat stress environments. Front. Plant Sci. 2018, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.; Giraldo, P.; Gonzalez, J.M. Phenotypic variation in root architecture traits and their relationship with eco-geographical and agronomic features in a core collection of tetraploid wheat landraces (Triticum turgidum L.). Euphytica 2018, 214, 54. [Google Scholar] [CrossRef]
- Alahmad, S.; El Hassouni, K.; Bassi, F.M.; Dinglasan, E.; Youssef, C.; Quarry, G.; Aksoy, A.; Mazzucotelli, E.; Juhász, A.; Able, J.A.; et al. A major root architecture QTL responding to water limitation in durum wheat. Front. Plant Sci. 2019, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, A.A.; Pour-Aboughadareh, A.; Mansouri, S.; Hosseini, A. Genome-wide association analysis of root system architecture features and agronomic traits in durum wheat. Mol. Breed. 2020, 40, 55. [Google Scholar] [CrossRef]
- Pradhan, A.K.; Kumar, S.; Singh, A.K.; Budhlakoti, N.; Mishra, D.C.; Chauhan, D.; Mittal, S.; Grover, M.; Kumar, S.; Gangwar, O.P.; et al. Identification of QTLs/defense genes effective at seedling stage against prevailing races of wheat stripe rust in India. Front. Genet. 2020, 11, 572975. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, 7191. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Li, J.; Cheng, Y.; Yao, F.; Long, L.; Wang, Y.; Wu, Y.; Li, J.; Wang, J.; Jiang, Q. Genome-wide association study reveals new loci for yield-related traits in Sichuan wheat germplasm under stripe rust stress. BMC Genom. 2019, 20, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Line, R.F.; Qayoum, A. Virulence, aggressiveness, evolution, and distribution of races of Puccinia striiformis (the cause of stripe rust of wheat) in North America. USDA-ARS Tech. Bull. 1992, 1788, 44. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploidy wheat genomic diversity using a high-density 90000 single nucleotide polymorphism array. Plant. Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformation 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Hwang, E.Y.; Song, Q.; Jia, G.; Specht, J.E.; Hyten, D.L.; Costa, J.; Cregan, P.B. A genome-wide association study of seed protein and oil content in soybean. BMC Genom. 2014, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.K.; Kumar, A.; Grover, N.; Ellur, R.K.; Bollinedi, H.; Krishnan, S.G.; Bhowmick, P.K.; Vinod, K.K.; Nagarajan, M.; Singh, A.K. Genome-Wide Association Study Reveals Marker-Trait Associations for Early Vegetative Stage Salinity Tolerance in Rice. Plants 2021, 10, 559. [Google Scholar] [CrossRef]
- Teshome, D.T.; Zharare, G.E.; Naidoo, S. The Threat of the Combined Effect of Biotic and Abiotic Stress Factors in Forestry Under a Changing Climate. Front. Plant Sci. 2020, 11, 601009. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, H.; Wu, N.; Ma, J.; Yuan, M.; Zhang, Z.; Yuan, S.; Zhang, H. Effects of Stripe Rust Infection on the Levels of Redox Balance and Photosynthetic Capacities in Wheat. Int. J. Mol. Sci. 2020, 21, 268. [Google Scholar] [CrossRef] [Green Version]
- Wheat & Small Grains. Washington State University. Available online: https://smallgrains.wsu.edu/disease-resources/foliar-fungal-diseases/stripe-rust/ (accessed on 10 February 2022).
- Bansal, U.K.; Kazi, A.G.; Singh, B.; Hare, R.A.; Bariana, H.S. Mapping of durable stripe rust resistance in a durum wheat cultivar Wollaroi. Mol. Breeding 2014, 33, 51–59. [Google Scholar] [CrossRef]
- Rosewarne, G.M.; Herrera Foessel, S.A.; Singh, R.P.; Huerta Espino, J.; Lan, C.X.; He, Z.H. Quantitative trait loci of stripe rust resistance in wheat. Theor. Appl. Genet. 2013, 126, 2427–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokore, F.E.; Ruan, Y.; Mccartney, C.; Knox, R.E.; Pei, X.; Aboukhaddour, R.; Randhawa, H.; Ammar, K.; Meyer, B.; Cuthbert, R.D.; et al. High density genetic mapping of stripe rust resistance in a ‘Strongfield’/‘Blackbird’ durum wheat population. Can. J. Plant Pathol. 2021, 43, 242–255. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Kianersi, F.; Poczai, P.; Moradkhani, H. Potential of wild relatives of wheat: Ideal genetic resources for future breeding programs. Agronomy 2021, 11, 1656. [Google Scholar] [CrossRef]
- Atwell, S.; Huang, Y.S.; Vilhjálmsson, B.J.; Willems, G.; Horton, M.; Li, Y.; Meng, D.; Platt, A.; Tarone, A.M.; Hu, T.T.; et al. Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines. Nature 2010, 465, 627–631. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Thornsberry, J.M.; Buckler, E.S. Structure of linkage disequilibrium in plants. Annu. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Cory, A.T.; Haile, J.; Kutcher, H.R.; Ammar, K.; Loladze, A.; Huerta-Espino, J.; et al. Genetic analysis of resistance to stripe rust in durum wheat (Triticum turgidum L. var. durum). PLoS ONE 2018, 13, e0203283. [Google Scholar] [CrossRef]
- Kabbaj, H.; Sall, A.T.; Al-Abdallat, A.; Geleta, M.; Amri, A.; Filali-Maltouf, A.; Belkadi, B.; Ortiz, R.; Bassi, F.M. Genetic diversity within a global panel of durum wheat (Triticum durum) landraces and modern germplasm reveals the history of alleles exchange. Front. Plant Sci. 2017, 8, 1277. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Mesquita, A.C.O.; Figueiró, A.D.A.; Wu, X.; Manjunatha, S.; Wickland, D.P.; Hudson, M.E.; Juliatti, F.C.; Clough, S.J. Genome-wide association mapping of resistance to a Brazilian isolate of Sclerotinia sclerotiorum in soybean genotypes mostly from Brazil. BMC Genom. 2017, 18, 849. [Google Scholar] [CrossRef] [Green Version]
- Voss-Fels, K.; Frisch, M.; Qian, L.; Kontowski, S.; Friedt, W.; Gottwald, S.; Snowdon, R.J. Subgenomic diversity patterns caused by directional selection in bread wheat gene pools. Plant Genome 2015, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Maccaferri, M.; Rynearson, S.; Letta, T.; Zegeye, H.; Tuberosa, R.; Chen, X.; Pumphrey, M. Novel sources of stripe rust resistance identified by genomewide association mapping in Ethiopian durum wheat (Triticum turgidum ssp. durum). Front. Plant Sci. 2017, 8, 774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H.; Shafqat, N. Genome-wide association analyses for yield and yield-related traits in bread wheat (Triticum aestivum L.) under pre-anthesis combined heat and drought stress in field conditions. PLoS ONE 2019, 14, e0213407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, I.; Saripalli, G.; Kumar, K.; Kumar, A.; Singh, R.; Batra, R.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Meta-QTLs and candidate genes for strip rust resistance in wheat. Sci. Rep. 2021, 11, 22923. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, A.; Chhokar, V.; Gangwar, O.P.; Bhardwaj, S.C.; Sivasamy, M.; Prasad, S.V.; Prakasha, T.L.; Khan, H.; Singh, R.; et al. Genome-wide association studies in diverse spring wheat panel for stripe, stem and leaf rust resistance. Front. Plant Sci. 2020, 11, 748. [Google Scholar] [CrossRef]
- Debibakas, S.; Rocher, S.; Garsmeur, O.; Toubi, L.; Roques, D.; D’Hont, A.; Hoarau, J.Y.; Daugrois, J.H. Prospecting sugarcane resistance to sugarcane yellow leaf virus by genome-wide association. Theor. Appl. Genet. 2014, 127, 1719–1732. [Google Scholar] [CrossRef] [Green Version]
- Zegeye, H.; Rasheed, A.; Makdis, F.; Badebo, A.; Ogbonnaya, F.C. Genome-wide association mapping for seedling and adult plant resistance to stripe rust in synthetic hexaploid wheat. PLoS ONE 2014, 9, e105593. [Google Scholar] [CrossRef] [Green Version]
- Muleta, K.T.; Rouse, M.N.; Rynearson, S.; Chen, X.; Buta, B.G.; Pumphrey, M.O. Characterization of molecular diversity and genome-wide mapping of loci associated with resistance to stripe rust and stem rust in Ethiopian bread wheat genotypes. BMC Plant Biol. 2017, 17, 134. [Google Scholar] [CrossRef]
- Ye, X.; Li, J.; Cheng, Y.; Yao, F.; Long, L.; Yu, C.; Wang, Y.; Wu, Y.; Li, J.; Wang, J.; et al. Genome-wide association study of resistance to stripe rust (Puccinia striiformis f. sp. tritici) in Sichuan wheat. BMC Plant Biol. 2019, 19, 147. [Google Scholar]



{kind=link}
{kind=link}
| No. | Gene Bank Code | Region | Pstv-37 | Pstv-40 | No. | Gene Bank Code | Region | Pstv-37 | Pstv-40 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | IUGB-00650 | Iran, Moghan | MS | MR | 71 | IUGB-00720 | Iran, Unknown | MS | MS |
| 2 | IUGB-00651 | Iran, Gorgan | R | R | 72 | IUGB-00721 | Iran, Unknown | S | S |
| 3 | IUGB-00652 | Iran, Mianeh | MS | MR | 73 | IUGB-00722 | Iran, Unknown | R | R |
| 4 | IUGB-00653 | Iran, Shirvan | S | S | 74 | IUGB-00723 | Iran, Unknown | MS | MR |
| 5 | IUGB-00654 | Iran, Unknown | MS | S | 75 | IUGB-00724 | Iran, Unknown | S | S |
| 6 | IUGB-00655 | Iran, Unknown | S | MR | 76 | IUGB-00726 | Iran, Unknown | S | S |
| 7 | IUGB-00656 | Iran, Unknown | S | MS | 77 | IUGB-00727 | Iran, Unknown | R | R |
| 8 | IUGB-00657 | Iran, Mashhad | S | S | 78 | IUGB-00728 | Iran, Bam | R | MS |
| 9 | IUGB-00658 | Iran, Mashhad | S | S | 79 | IUGB-00729 | Iraq | R | R |
| 10 | IUGB-00659 | Iran, Mashhad | MS | R | 80 | IUGB-00730 | Iran, Shoushtar | MS | MS |
| 11 | IUGB-00660 | Iran, Mashhad | MS | MS | 81 | IUGB-00731 | Iran, Kermanshah | R | S |
| 12 | IUGB-00661 | Iran, Mashhad | MS | MR | 82 | IUGB-00732 | Iran, Kermanshah | MS | S |
| 13 | IUGB-00662 | Iran, Mashhad | S | MS | 83 | IUGB-00733 | Iran, Golpaygan | S | S |
| 14 | IUGB-00663 | Iran, Mashhad | S | MS | 84 | IUGB-00734 | Iran, Unknown | S | MS |
| 15 | IUGB-00664 | Iran, Mashhad | S | MS | 85 | IUGB-00735 | Iran, Unknown | S | MS |
| 16 | IUGB-00665 | Iran, Mashhad | MR | MR | 86 | IUGB-00736 | Iran, Unknown | MS | MS |
| 17 | IUGB-00666 | Iran, Mashhad | S | MS | 87 | IUGB-00737 | Iran, Unknown | MR | R |
| 18 | IUGB-00667 | Iran, Galuran | S | MS | 88 | IUGB-00738 | Iran, Unknown | MS | R |
| 19 | IUGB-00668 | Iran, Galuran | S | S | 89 | IUGB-00740 | Iran, Mashhad | S | S |
| 20 | IUGB-00669 | Iran, Kabkali | S | MS | 90 | IUGB-00741 | Iran, Mashhad | MR | MS |
| 21 | IUGB-00670 | Iran, Shetaban | S | S | 91 | IUGB-00742 | Iran, Mashhad | MR | R |
| 22 | IUGB-00671 | Iran, Zigh Abad | S | MS | 92 | IUGB-00743 | Iran, Mashhad | S | MS |
| 23 | IUGB-00672 | Iran, Mahidasht | MR | R | 93 | IUGB-00744 | Iran, Mashhad | S | MS |
| 24 | IUGB-00673 | Iran, Unknown | MR | MR | 94 | IUGB-00745 | Iran, Mashhad | MS | MS |
| 25 | IUGB-00674 | Iran, Songhor | R | R | 95 | IUGB-00746 | Iran, Mashhad | S | S |
| 26 | IUGB-00675 | Iran, Kangavar | R | R | 96 | IUGB-00747 | Iran, Mashhad | MS | MS |
| 27 | IUGB-00676 | Iran, Aleshtar | MS | MR | 97 | IUGB-00750 | Iran, Mashhad | R | R |
| 28 | IUGB-00677 | Iran, Azna | S | MS | 98 | IUGB-00751 | Iran, Mashhad | R | R |
| 29 | IUGB-00678 | Iran, Delfan | R | R | 99 | IUGB-00752 | Iran, Mashhad | R | R |
| 30 | IUGB-00679 | Iran, Mehran | MR | MR | 100 | IUGB-00753 | Iran, Mashhad | MR | MR |
| 31 | IUGB-00680 | Iran, Shebab | MS | S | 101 | IUGB-00754 | Iran, Mashhad | R | R |
| 32 | IUGB-00681 | Yugoslavia | MS | MR | 102 | IUGB-00755 | Iran, Unknown | MS | MS |
| 33 | IUGB-00682 | Afghanistan | S | S | 103 | IUGB-00757 | Iran, Unknown | MS | MS |
| 34 | IUGB-00683 | Iran, Dehgolan | S | MS | 104 | IUGB-00758 | Iran, Unknown | MS | MS |
| 35 | IUGB-00684 | Iran, Marivan | S | S | 105 | IUGB-00759 | Iran, Lorestan | MS | MS |
| 36 | IUGB-00685 | Portugal | R | R | 106 | IUGB-00760 | Iran, Lorestan | R | R |
| 37 | IUGB-00686 | Afghanistan | S | MS | 107 | IUGB-00761 | Iran, Unknown | S | MS |
| 38 | IUGB-00687 | Bulgaria | S | S | 108 | IUGB-00762 | Iran, Paveh | MS | MS |
| 39 | IUGB-00688 | Argentina | R | R | 109 | IUGB-00763 | Iran, Kermanshah | MR | MR |
| 40 | IUGB-00689 | Australia | MS | MS | 110 | IUGB-00764 | Iran, Unknown | MR | MS |
| 41 | IUGB-00690 | Bulgaria | S | MS | 111 | IUGB-00765 | Iran, Kermanshah | MS | R |
| 42 | IUGB-00691 | Iran, Lorestan | MS | MS | 112 | IUGB-00766 | Iran, Kermanshah | MS | MS |
| 43 | IUGB-00693 | Iran, Dezful | MR | MR | 113 | IUGB-00767 | Iran, Kermanshah | MS | MS |
| 44 | IUGB-00694 | Iran, Lorestan | MS | MS | 114 | IUGB-00768 | Iran, Kermanshah | S | MS |
| 45 | IUGB-00695 | Iran, Lorestan | MS | R | 115 | IUGB-00769 | Iran, Kermanshah | MS | MS |
| 46 | IUGB-00696 | Iran, Lorestan | S | S | 116 | IUGB-00770 | Iran, Gachsaran | MR | MS |
| 47 | IUGB-00697 | Iran, Lorestan | S | S | 117 | IUGB-00771 | Iran, Kermanshah | MS | MS |
| 48 | IUGB-00698 | Iran, Lorestan | R | R | 118 | IUGB-00772 | Iran, Hamadan | MS | S |
| 49 | IUGB-00699 | Iran, Lorestan | R | R | 119 | IUGB-00773 | Iran, Eizeh | S | R |
| 50 | IUGB-00700 | Iran, Lorestan | S | MS | 120 | IUGB-00774 | Iran, Eizeh | S | S |
| 51 | IUGB-00701 | Iran, Lorestan | S | MS | 121 | IUGB-00775 | Iran, Dezful | S | S |
| 52 | IUGB-00702 | Iran, Lorestan | MS | MS | 122 | IUGB-00776 | Iran, Dezful | MS | MR |
| 53 | IUGB-00703 | Iran, Lorestan | MS | MS | 123 | IUGB-00777 | Iran, Ardebil | MS | MR |
| 54 | IUGB-00704 | Iran, Kermanshah | S | S | 124 | IUGB-00778 | Iran, Ardebil | S | MS |
| 55 | IUGB-00705 | Iran, Kermanshah | R | R | 125 | IUGB-00779 | Iran, Ardebil | MS | MS |
| C56 | IUGB-00706 | Iran, Lorestan | MR | MR | 126 | IUGB-00780 | Iran, Ahar | MS | MS |
| 57 | IUGB-00707 | Iran, Lorestan | MR | MR | 127 | IUGB-00781 | Iran, Ahar | MS | S |
| 58 | IUGB-00708 | Iran, Lorestan | MS | MS | 128 | IUGB-00782 | Iran, Lorestan | S | MS |
| 59 | IUGB-00709 | Iran, Lorestan | MS | MS | 129 | IUGB-00783 | Iran, Lorestan | MS | MR |
| 60 | IUGB-00710 | Iran, Lorestan | MS | S | 130 | IUGB-00784 | Iran, East Azarbayjan | MS | MS |
| 61 | IUGB-00711 | Iran, Unknown | MS | S | 131 | IUGB-00785 | Iran, Lorestan | MS | MS |
| 62 | IUGB-00712 | Iran, Lorestan | MS | MS | 132 | IUGB-00786 | Italy | MR | MR |
| 63 | IUGB-00713 | Iran, Unknown | MS | MS | 133 | IUGB-00787 | Italy | R | MR |
| 64 | IUGB-00714 | Iran, Unknown | S | S | 134 | IUGB-00788 | Italy | MR | MR |
| 65 | IUGB-00715 | Iran, Unknown | MS | MS | 135 | IUGB-00790 | Italy | S | MS |
| 66 | IUGB-00716 | Iran, Unknown | MS | MR | 136 | IUGB-00791 | Italy | MR | R |
| 67 | IUGB-00717 | Iran, Lorestan | MS | MS | 137 | IUGB-00792 | Italy | R | R |
| 68 | IUGB-00718 | Iran, Unknown | MS | S | 138 | IUGB-00793 | Italy | MS | S |
| 69 | IUGB-00718 | Iran, Unknown | MS | S | 139 | IUGB-00945 | Iran, Dareh Shahr | MS | MR |
| 70 | IUGB-00719 | Iran, Unknown | S | S |
| Chromosome | Numbers of SNPs | Length (cM) | * Average Linkage Disequilibrium (r2) between Pair of Markers | ||||
|---|---|---|---|---|---|---|---|
| D < 5 cM | D = 5–10 cM | D = 10–20 cM | D = 20–50 cM | D > 50 cM | |||
| 1A | 181 | 106 | 0.214 | 0.094 | 0.069 | 0.048 | 0.035 |
| 1B | 249 | 101.5 | 0.127 | 0.044 | 0.043 | 0.039 | 0.034 |
| 2A | 206 | 119.7 | 0.147 | 0.041 | 0.092 | 0.037 | 0.032 |
| 2B | 391 | 144.2 | 0.149 | 0.083 | 0.065 | 0.043 | 0.041 |
| 3A | 184 | 163.4 | 0.264 | 0.099 | 0.051 | 0.052 | 0.047 |
| 3B | 340 | 137 | 0.139 | 0.052 | 0.055 | 0.038 | 0.035 |
| 4A | 168 | 161.8 | 0.238 | 0.029 | 0.33 | 0.023 | 0.039 |
| 4B | 150 | 107 | 0.147 | 0.044 | 0.036 | 0.044 | 0.048 |
| 5A | 193 | 106.7 | 0.217 | 0.077 | 0.049 | 0.037 | 0.061 |
| 5B | 270 | 179.6 | 0.25 | 0.062 | 0.038 | 0.033 | 0.031 |
| 6A | 240 | 120.2 | 0.317 | 0.047 | 0.153 | 0.04 | 0.034 |
| 6B | 231 | 110.4 | 0.127 | 0.046 | 0.041 | 0.038 | 0.039 |
| 7A | 251 | 165.9 | 0.149 | 0.043 | 0.043 | 0.031 | 0.04 |
| 7B | 267 | 142 | 0.145 | 0.049 | 0.042 | 0.041 | 0.032 |
| Trait | Marker | Position | Chr. | Marker | Position | Chr. | Marker | Position | Chr. |
|---|---|---|---|---|---|---|---|---|---|
| Pstv-37 | BobWhite_c53978_99 | 115 | 6A | IACX8074 | 97 | 1B | wsnp_Ex_c2617_4864441 | 61 | 4A |
| Pstv-40 | 115 | 6A | 97 | 1B | 61 | 4A | |||
| Pstv-37 | BobWhite_c7786_376 | 64 | 2B | IACX8294 | 116 | 7B | wsnp_Ex_c2617_4864955 | 61 | 4A |
| Pstv-40 | 64 | 2B | 116 | 7B | 61 | 4A | |||
| Pstv-37 | BobWhite_c8428_346 | 63 | 1A | IACX9290 | 97 | 1B | wsnp_Ex_c54395_57291841 | 61 | 4A |
| Pstv-40 | 63 | 1A | 97 | 1B | 61 | 4A | |||
| Pstv-37 | CAP7_c4064_162 | 108 | 5A | Kukri_c93635_290 | 61 | 4A | wsnp_Ex_c6044_10590220 | 61 | 4A |
| Pstv-40 | 108 | 5A | 61 | 4A | 61 | 4A | |||
| Pstv-37 | Ex_c5759_628 | 63 | 1A | RAC875_c5556_328 | 45 | 1B | wsnp_Ex_c7002_12063325 | 115 | 6A |
| Pstv-40 | 63 | 1A | 45 | 1B | 115 | 6A | |||
| Pstv-37 | Excalibur_c24041_794 | 63 | 1A | Tdurum_contig12525_769 | 116 | 7B | wsnp_Ex_c7002_12063380 | 115 | 6A |
| Pstv-40 | 63 | 1A | 116 | 7B | 115 | 6A | |||
| Pstv-37 | Excalibur_c32735_603 | 64 | 4A | Tdurum_contig44173_572 | 55 | 6B | wsnp_Ex_c7550_12907422 | 61 | 4A |
| Pstv-40 | 64 | 4A | 55 | 6B | 61 | 4A | |||
| Pstv-37 | Excalibur_c7002_314 | 115 | 6A | Tdurum_contig47269_904 | 55 | 6B | wsnp_Ex_c831_1625061 | 9 | 5B |
| Pstv-40 | 115 | 6A | 55 | 6B | 9 | 5B | |||
| Pstv-37 | Excalibur_rep_c110429_536 | 140 | 7B | Tdurum_contig7981_70 | 55 | 6B | wsnp_Ku_c16522_25425455 | 55 | 6B |
| Pstv-40 | 140 | 7B | 55 | 6B | 55 | 6B | |||
| Pstv-37 | GENE-0416_480 | 97 | 1B | wsnp_Ex_c12818_20334501 | 61 | 4A | wsnp_Ku_c30381_40208899 | 61 | 4A |
| Pstv-40 | 97 | 1B | 61 | 4A | 61 | 4A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehrabi, A.A.; Steffenson, B.J.; Pour-Aboughadareh, A.; Matny, O.; Rahmatov, M. Genome-Wide Association Study Identifies Two Loci for Stripe Rust Resistance in a Durum Wheat Panel from Iran. Appl. Sci. 2022, 12, 4963. https://0-doi-org.brum.beds.ac.uk/10.3390/app12104963
Mehrabi AA, Steffenson BJ, Pour-Aboughadareh A, Matny O, Rahmatov M. Genome-Wide Association Study Identifies Two Loci for Stripe Rust Resistance in a Durum Wheat Panel from Iran. Applied Sciences. 2022; 12(10):4963. https://0-doi-org.brum.beds.ac.uk/10.3390/app12104963
Chicago/Turabian StyleMehrabi, Ali Ashraf, Brian J. Steffenson, Alireza Pour-Aboughadareh, Oadi Matny, and Mahbubjon Rahmatov. 2022. "Genome-Wide Association Study Identifies Two Loci for Stripe Rust Resistance in a Durum Wheat Panel from Iran" Applied Sciences 12, no. 10: 4963. https://0-doi-org.brum.beds.ac.uk/10.3390/app12104963