
Decreased SLC27A5 Suppresses Lipid Synthesis and Tyrosine Metabolism to Activate the Cell Cycle in Hepatocellular Carcinoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Human Proteome Map (HPM)
2.2. The Cancer Genome Atlas (TCGA)
2.3. Gene Expression Omnibus (GEO)
2.4. Gene Set Enrichment Analysis (GSEA)
2.5. The Kaplan-Meier Plotter (KM Plotter)
2.6. The Human Protein Atlas (HPA)
2.7. Cell Culture and Stable Knockdown Cell Lines
2.8. RNA Extraction and Real-Time PCR Analysis (qPCR)
2.9. Reagent
2.10. Statistics
3. Results
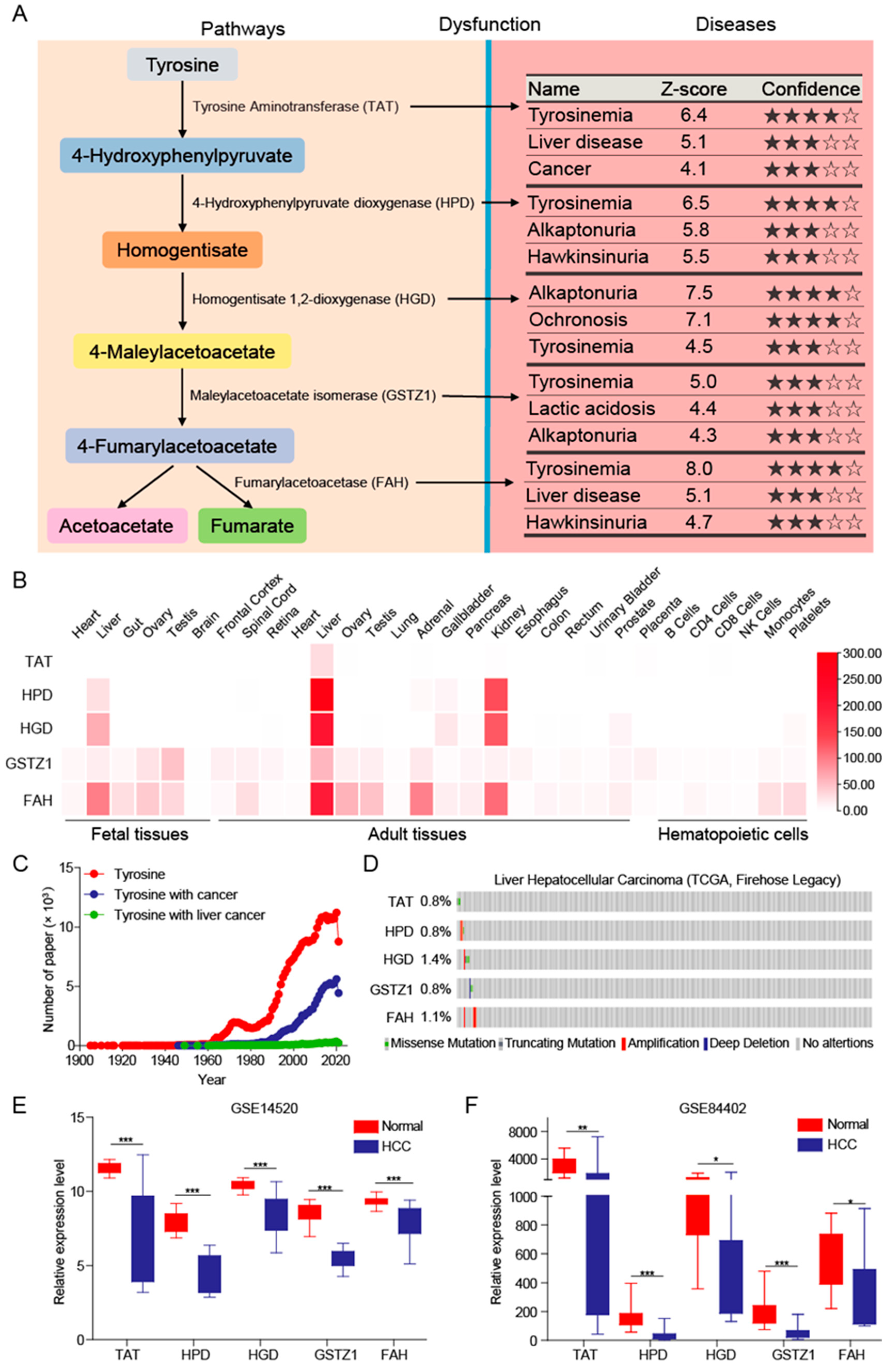
3.1. Tyrosine Metabolism Pathway Was Inhibited in HCC
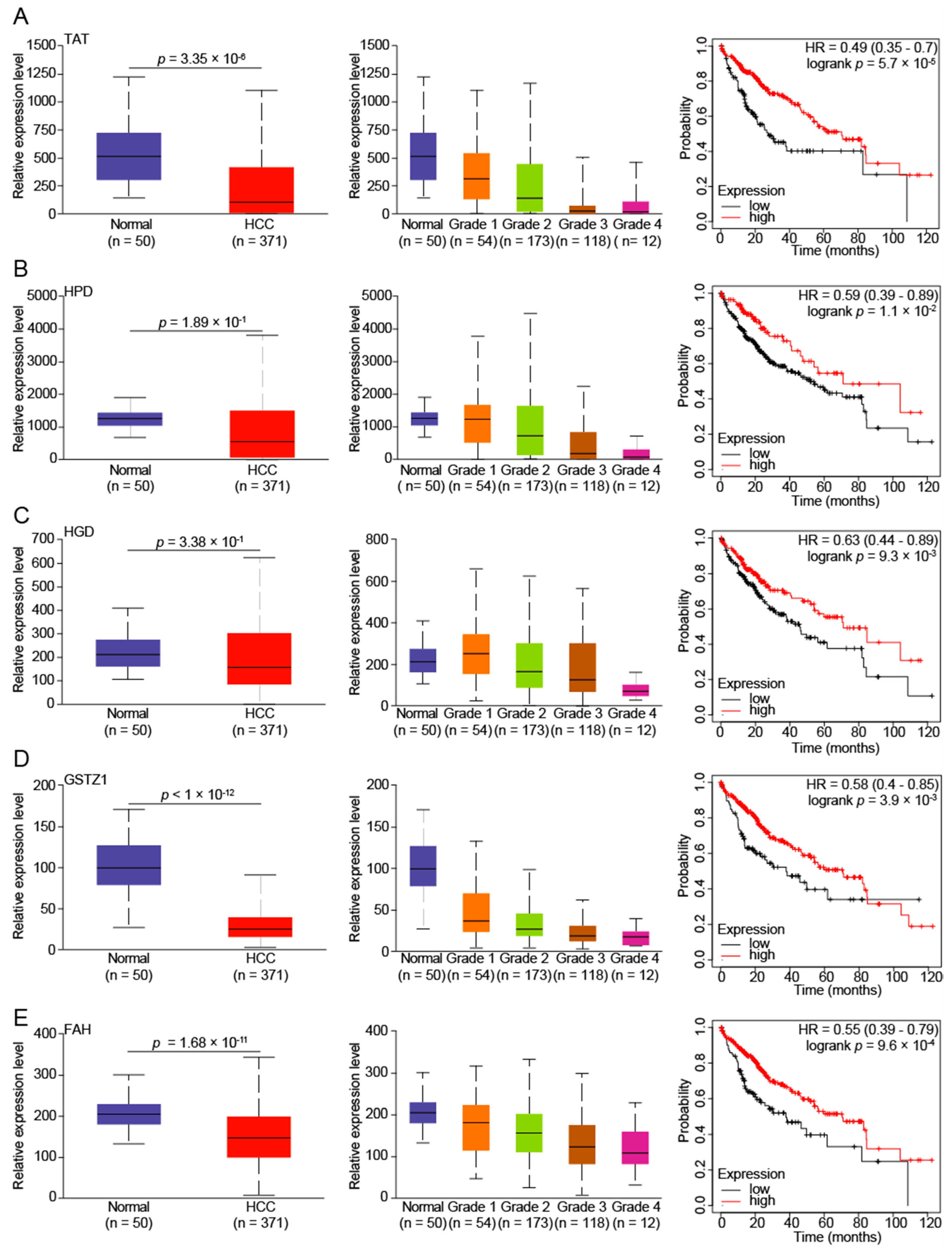
3.2. Reduced Tyrosine Metabolism Predicts Poor Prognosis in HCC Patients
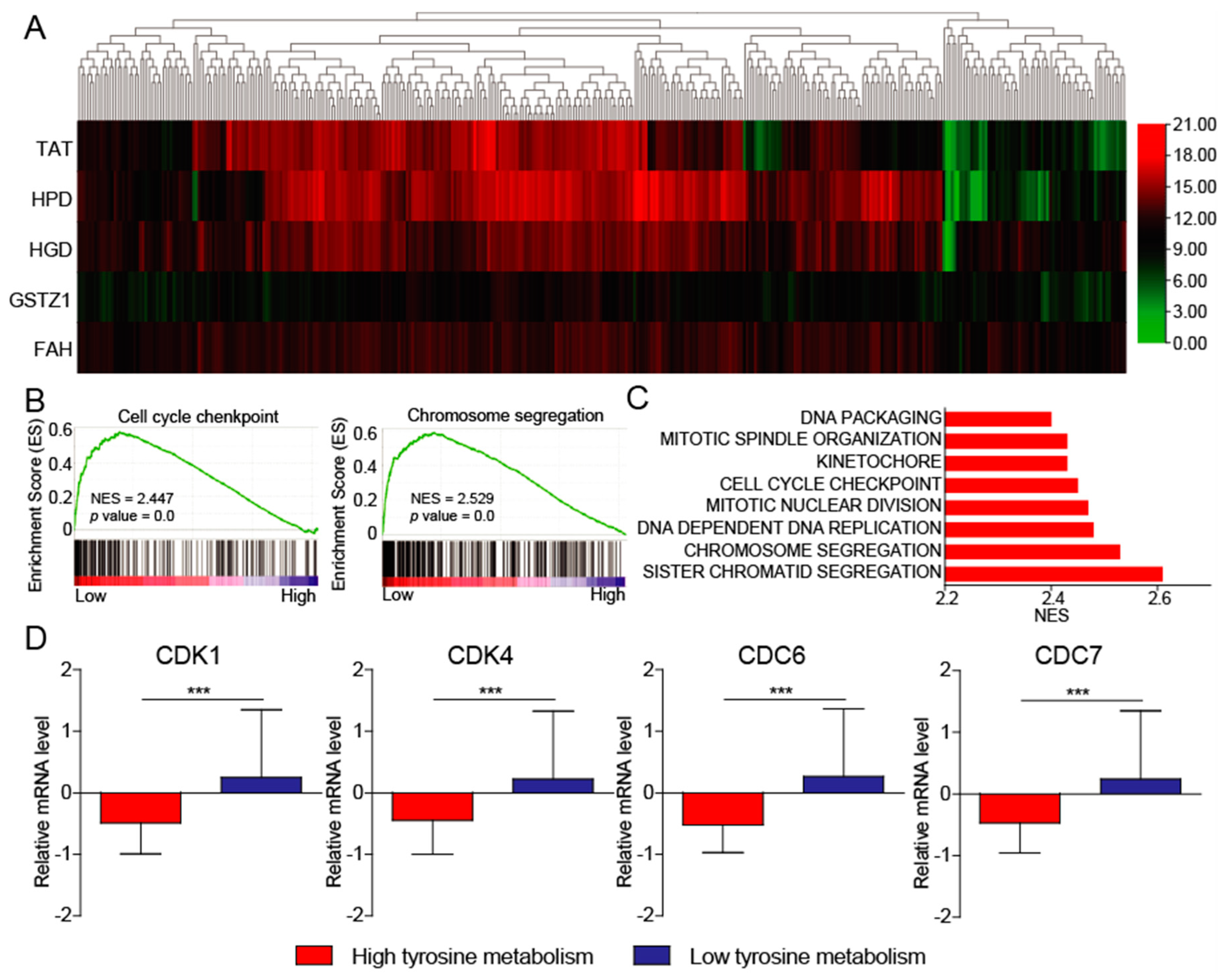
3.3. Tyrosine Metabolism Regulates Cell Cycle
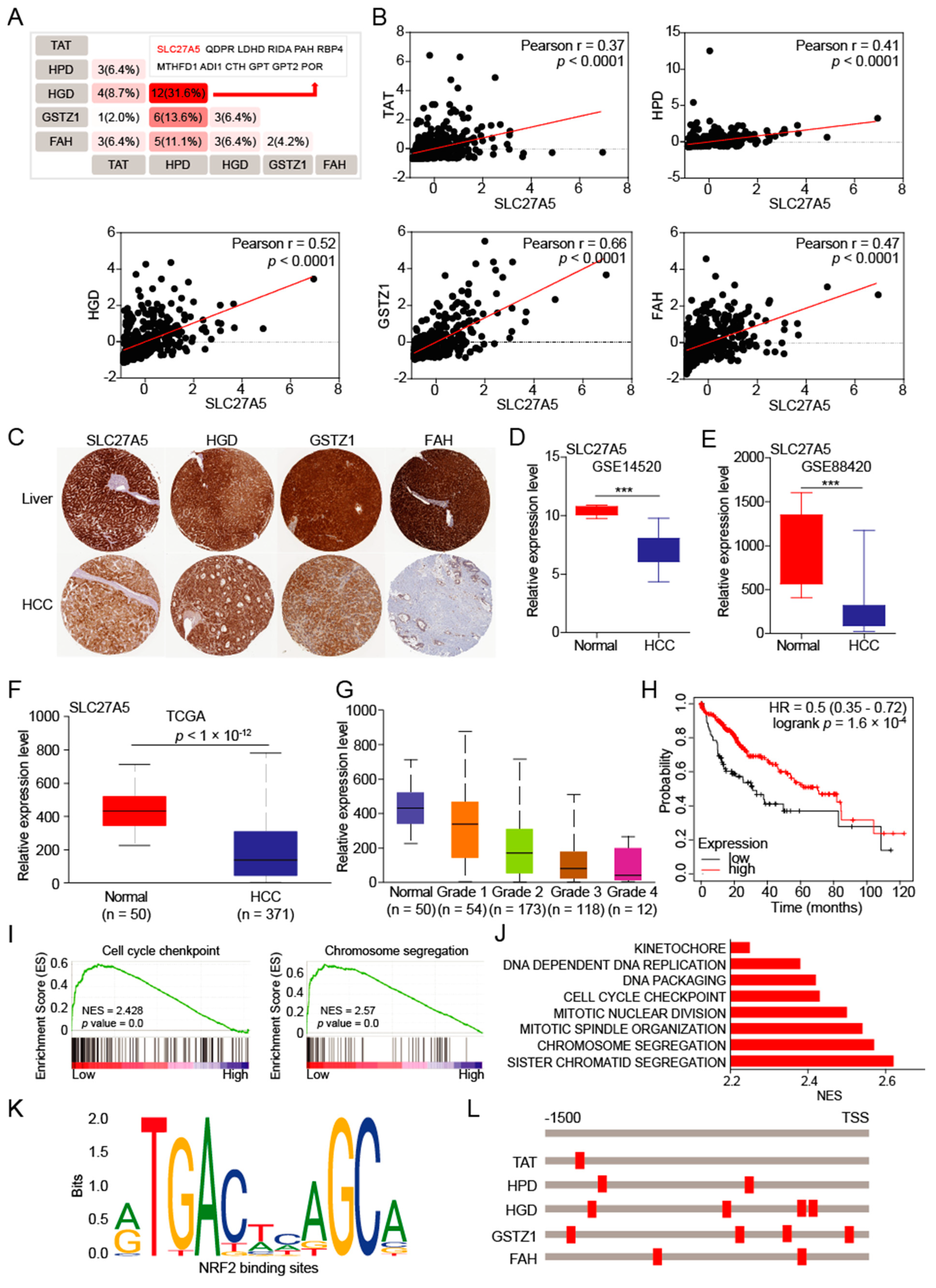
3.4. The SLC27A5 Is Positively Correlated with Expression of Tyrosine-metabolizing Enzymes and Regulates Cell Cycle
3.5. KnockdownSLC27A5 Promotes Growth of Human Hepatoma Cell
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tabe, Y.; Lorenzi, P.L.; Konopleva, M. Amino acid metabolism in hematologic malignancies and the era of targeted therapy. Blood 2019, 134, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, P.C. Acetyl-CoA Metabolism and Histone Acetylation in the Regulation of Aging and Lifespan. Antioxidants 2021, 10, 572. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Lu, Z.; Li, Z.; Sheng, H.; Fan, J.; Qi, Q. 4-hydroxyphenylpyruvate dioxygenase promotes lung cancer growth via pentose phosphate pathway (PPP) flux mediated by LKB1-AMPK/HDAC10/G6PD axis. Cell Death Dis. 2019, 10, 525. [Google Scholar] [CrossRef]
- Li, J.; Wang, Q.; Yang, Y.; Lei, C.; Yang, F.; Liang, L.; Chen, C.; Xia, J.; Wang, K.; Tang, N. GSTZ1 deficiency promotes hepatocellular carcinoma proliferation via activation of the KEAP1/NRF2 pathway. J. Exp. Clin. Cancer Res. 2019, 38, 438. [Google Scholar] [CrossRef] [PubMed]
- Hickey, R.D.; Mao, S.A.; Glorioso, J.; Lillegard, J.B.; Fisher, J.E.; Amiot, B.; Rinaldo, P.; Harding, C.O.; Marler, R.; Finegold, M.J.; et al. Fumarylacetoacetate hydrolase deficient pigs are a novel large animal model of metabolic liver disease. Stem Cell Res. 2014, 13, 144–153. [Google Scholar] [CrossRef]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Ben-Sahra, I.; Manning, B.D. mTORC1 signaling and the metabolic control of cell growth. Curr. Opin. Cell Biol. 2017, 45, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Condon, K.J.; Sabatini, D.M. Nutrient regulation of mTORC1 at a glance. J. Cell Sci. 2019, 132, jcs222570. [Google Scholar] [CrossRef]
- Anderson, C.M.; Stahl, A. SLC27 fatty acid transport proteins. Mol. Asp. Med. 2013, 34, 516–528. [Google Scholar] [CrossRef] [Green Version]
- Enooku, K.; Tsutsumi, T.; Kondo, M.; Fujiwara, N.; Sasako, T.; Shibahara, J.; Kado, A.; Okushin, K.; Fujinaga, H.; Nakagomi, R.; et al. Hepatic FATP5 expression is associated with histological progression and loss of hepatic fat in NAFLD patients. J. Gastroenterol. 2020, 55, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Pal Pathak, D.; Asthana, S. Bile acids mediated potential functional interaction between FXR and FATP5 in the regulation of Lipid Metabolism. Int. J. Biol. Sci. 2020, 16, 2308–2322. [Google Scholar] [CrossRef]
- Doege, H.; Baillie, R.A.; Ortegon, A.M.; Tsang, B.; Wu, Q.; Punreddy, S.; Hirsch, D.; Watson, N.; Gimeno, R.E.; Stahl, A. Targeted deletion of FATP5 reveals multiple functions in liver metabolism: Alterations in hepatic lipid homeostasis. Gastroenterology 2006, 130, 1245–1258. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, B.; Doege, H.; Punreddy, S.; Wu, H.; Huang, X.; Kaushik, V.K.; Mozell, R.L.; Byrnes, J.J.; Stricker-Krongrad, A.; Chou, C.J.; et al. Mice deleted for fatty acid transport protein 5 have defective bile acid conjugation and are protected from obesity. Gastroenterology 2006, 130, 1259–1269. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Xiao, Z.; Sun, S.; Wang, K.; Qian, J.; Cui, Z.; Tao, T. Integrated Proteomics and Bioinformatics to Identify Potential Prognostic Biomarkers in Hepatocellular Carcinoma. Cancer Manag. Res. 2021, 13, 2307–2317. [Google Scholar] [CrossRef]
- Kim, M.S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A draft map of the human proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, l1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roessler, S.; Long, E.L.; Budhu, A.; Chen, Y.; Zhao, X.; Ji, J.; Walker, R.; Jia, H.L.; Ye, Q.H.; Qin, L.X.; et al. Integrative genomic identification of genes on 8p associated with hepatocellular carcinoma progression and patient survival. Gastroenterology 2012, 142, 957–966.e12. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Huo, X.; Yang, X.R.; He, J.; Cheng, L.; Wang, N.; Deng, X.; Jin, H.; Wang, N.; Wang, C.; et al. STAT3-mediated upregulation of lncRNA HOXD-AS1 as a ceRNA facilitates liver cancer metastasis by regulating SOX4. Mol. Cancer 2017, 16, 136. [Google Scholar] [CrossRef] [Green Version]
- Győrffy, B. Survival analysis across the entire transcriptome identifies biomarkers with the highest prognostic power in breast cancer. Comput. Struct. Biotechnol. J. 2021, 19, 4101–4109. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Bouyacoub, Y.; Zribi, H.; Azzouz, H.; Nasrallah, F.; Abdelaziz, R.B.; Kacem, M.; Rekaya, B.; Messaoud, O.; Romdhane, L.; Charfeddine, C.; et al. Novel and recurrent mutations in the TAT gene in Tunisian families affected with Richner-Hanhart syndrome. Gene 2013, 529, 45–49. [Google Scholar] [CrossRef]
- Culic, V.; Betz, R.C.; Refke, M.; Fumic, K.; Pavelic, J. Tyrosinemia type II (Richner-Hanhart syndrome): A new mutation in the TAT gene. Eur. J. Med. Genet. 2011, 54, 205–208. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Q.; Yang, H.; Zou, Q.; Lai, C.; Jiang, F.; Zhao, P.; Luo, Z.; Yang, J.; Chen, Q.; et al. Fumarylacetoacetate Hydrolase Knock-out Rabbit Model for Hereditary Tyrosinemia Type 1. J. Biol. Chem. 2017, 292, 4755–4763. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Chen, S.L.; Lin, C.S.; Liu, L.L.; Wang, C.H.; Yun, J.P. Tyrosine metabolic enzyme HPD is decreased and predicts unfavorable outcomes in hepatocellular carcinoma. Pathol. Res. Pract. 2020, 216, 153153. [Google Scholar] [CrossRef]
- Yamashita, D.; Bernstock, J.D.; Elsayed, G.; Sadahiro, H.; Mohyeldin, A.; Chagoya, G.; Ilyas, A.; Mooney, J.; Estevez-Ordonez, D.; Yamaguchi, S.; et al. Targeting glioma-initiating cells via the tyrosine metabolic pathway. J. Neurosurg. 2020, 134, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Wang, Q.; Tang, N.; Wang, K. GSTZ1-1 downregulates Wnt/β-catenin signalling in hepatocellular carcinoma cells. FEBS Open Bio 2020, 10, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Annie-Mathew, A.S.; Prem-Santhosh, S.; Jayasuriya, R.; Ganesh, G.; Ramkumar, K.M.; Sarada, D.V.L. The pivotal role of Nrf2 activators in adipocyte biology. Pharmacol. Res. 2021, 173, 105853. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zhang, G.; Zheng, Y.; Yang, Y.; Chen, C.; Xia, J.; Liang, L.; Lei, C.; Hu, Y.; Cai, X.; et al. SLC27A5 deficiency activates NRF2/TXNRD1 pathway by increased lipid peroxidation in HCC. Cell Death Differ. 2020, 27, 1086–1104. [Google Scholar] [CrossRef]
- Miranda, C.A.; Guimarães, A.; Bizerra, P.F.V.; Mingatto, F.E. Diazinon impairs bioenergetics and induces membrane permeability transition on mitochondria isolated from rat liver. J. Toxicol. Environ. Health A 2020, 83, 616–629. [Google Scholar] [CrossRef]
- Al-Otaibi, A.M.; Al-Balawi, H.F.A.; Ahmad, Z.; Suliman, E.M. Toxicity bioassay and sub-lethal effects of diazinon on blood profile and histology of liver, gills and kidney of catfish, Clarias gariepinus. Braz. J. Biol. Rev. Brasleira Biol. 2019, 79, 326–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Wang, W.; Jiang, Y.; Chu, W. Diazinon exposure produces histological damage, oxidative stress, immune disorders and gut microbiota dysbiosis in crucian carp (Carassius auratus gibelio). Environ. Pollut. 2021, 269, 116129. [Google Scholar] [CrossRef] [PubMed]
- Boussabbeh, M.; Ben Salem, I.; Hamdi, M.; Ben Fradj, S.; Abid-Essefi, S.; Bacha, H. Diazinon, an organophosphate pesticide, induces oxidative stress and genotoxicity in cells deriving from large intestine. Environ. Sci. Pollut. Res. Int. 2016, 23, 2882–2889. [Google Scholar] [CrossRef]
- Yousefizadeh, S.; Farkhondeh, T.; Samarghandian, S. Age-Related Diazinon Toxicity Impact on Blood Glucose, Lipid Profile and Selected Biochemical Indices in Male Rats. Curr. Aging Sci. 2019, 12, 49–54. [Google Scholar] [CrossRef]
- Fu, L.; Dong, S.S.; Xie, Y.W.; Tai, L.S.; Chen, L.; Kong, K.L.; Man, K.; Xie, D.; Li, Y.; Cheng, Y.; et al. Down-regulation of tyrosine aminotransferase at a frequently deleted region 16q22 contributes to the pathogenesis of hepatocellular carcinoma. Hepatology 2010, 51, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′→3′) |
|---|---|
| SLC27A5 | Forward primer: CATGGCGTGACAGTGATCCT Reverse primer: CAGCCCGTAGTCCATTGCC |
| TAT | Forward primer: CTGGACTCGGGCAAATATAATGG Reverse primer: GTCCTTAGCTTCTAGGGGTGC |
| HPD | Forward primer: GGAGCCCTGGGTAGAGCAA Reverse primer: CAAGAATTGGCCGATGTAGTTCA |
| HGD | Forward primer: ATTTACACCGAGTTTGGCAAGA Reverse primer: GGTCTCCTCAAAGACATCTATGC |
| GSTZ1 | Forward primer: CGGGCATATACTGTGTAGGAGA Reverse primer: GGGGTGAGATCCACCTTGAA |
| FAH | Forward primer: TCTGCCACGATGCACCTTC Reverse primer: CCTTGTCCCTGAACATGATTCC |
| CDK1 | Forward primer: GGATGTGCTTATGCAGGATTCC Reverse primer: CATGTACTGACCAGGAGGGATAG |
| CDK4 | Forward primer: ATGGCTACCTCTCGATATGAGC Reverse primer: CATTGGGGACTCTCACACTCT |
| CDC6 | Forward primer: CCAGGCACAGGCTACAATCAG Reverse primer: AACAGGTTACGGTTTGGACATT |
| CDC7 | Forward primer: AGTGCCTAACAGTGGCTGG Reverse primer: CACGGTGAACAATACCAAACTGA |
| ACTIN | Forward primer: GGAAATCGTGCGTGACAT Reverse primer: TGCCAATGGTGATGACCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Qiao, Y.; Sun, H.; Chang, H.; Zhao, H.; Zhang, S.; Shan, C. Decreased SLC27A5 Suppresses Lipid Synthesis and Tyrosine Metabolism to Activate the Cell Cycle in Hepatocellular Carcinoma. Biomedicines 2022, 10, 234. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020234
Wang J, Qiao Y, Sun H, Chang H, Zhao H, Zhang S, Shan C. Decreased SLC27A5 Suppresses Lipid Synthesis and Tyrosine Metabolism to Activate the Cell Cycle in Hepatocellular Carcinoma. Biomedicines. 2022; 10(2):234. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020234
Chicago/Turabian StyleWang, Jiyan, Yaya Qiao, Huanran Sun, Hongkai Chang, Huifang Zhao, Shuai Zhang, and Changliang Shan. 2022. "Decreased SLC27A5 Suppresses Lipid Synthesis and Tyrosine Metabolism to Activate the Cell Cycle in Hepatocellular Carcinoma" Biomedicines 10, no. 2: 234. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020234