The brlA Gene Deletion Reveals That Patulin Biosynthesis Is Not Related to Conidiation in Penicillium expansum

 , , and
, , and
Abstract
:1. Introduction
2. Results
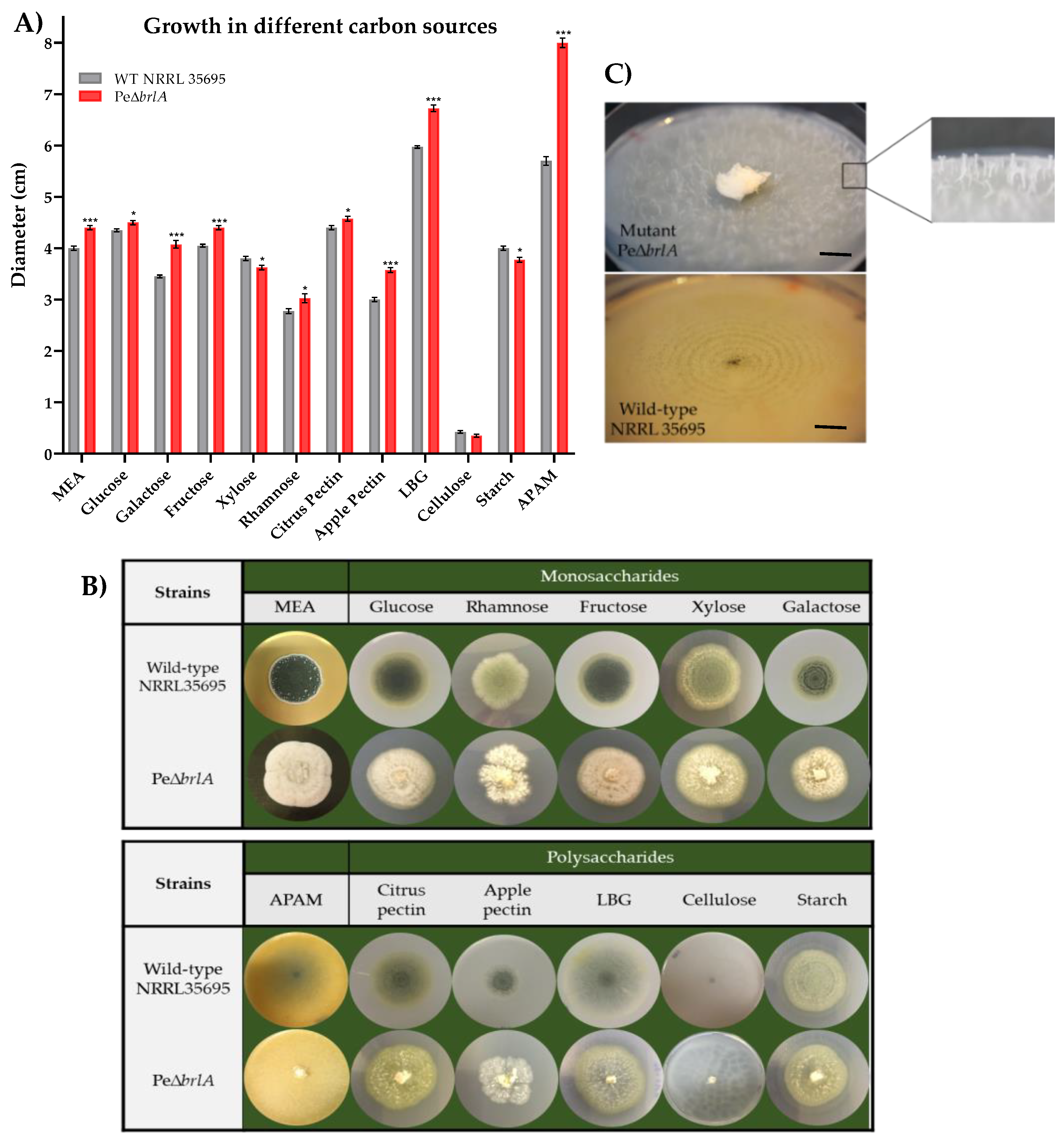
2.1. Effect of brlA Deletion on In Vitro Macroscopic and Microscopic Morphology
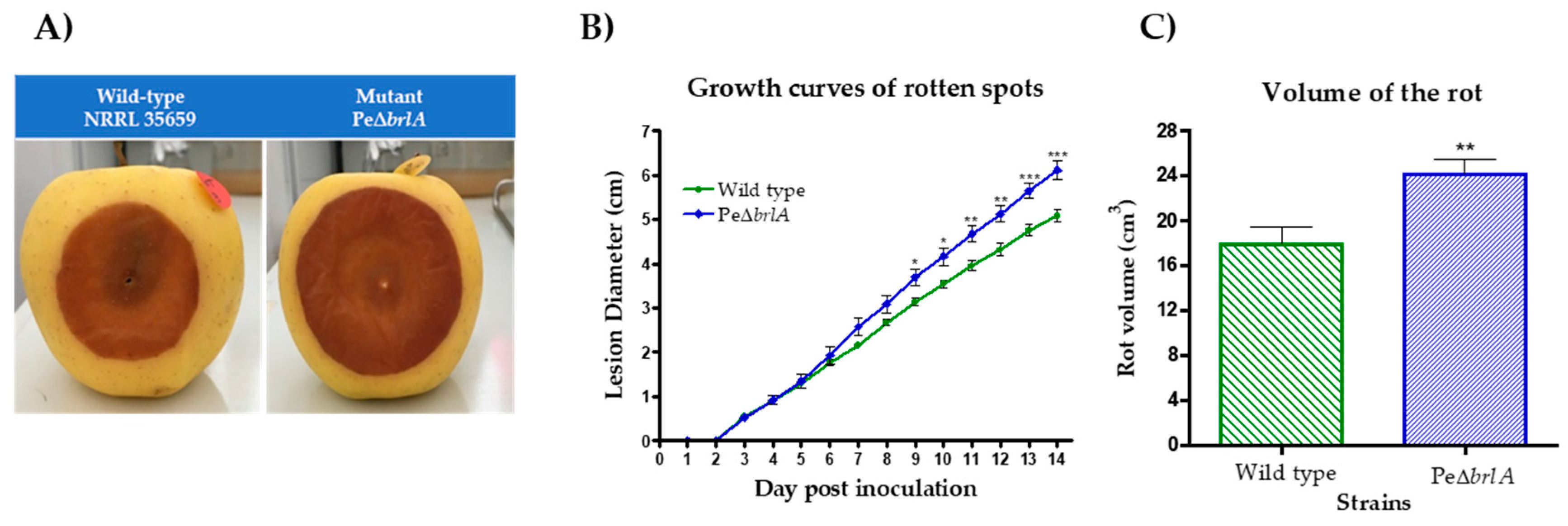
2.2. Effect of brlA Deletion on Apple Colonization
2.2.1. Effect of brlA Deletion on Pathogenicity
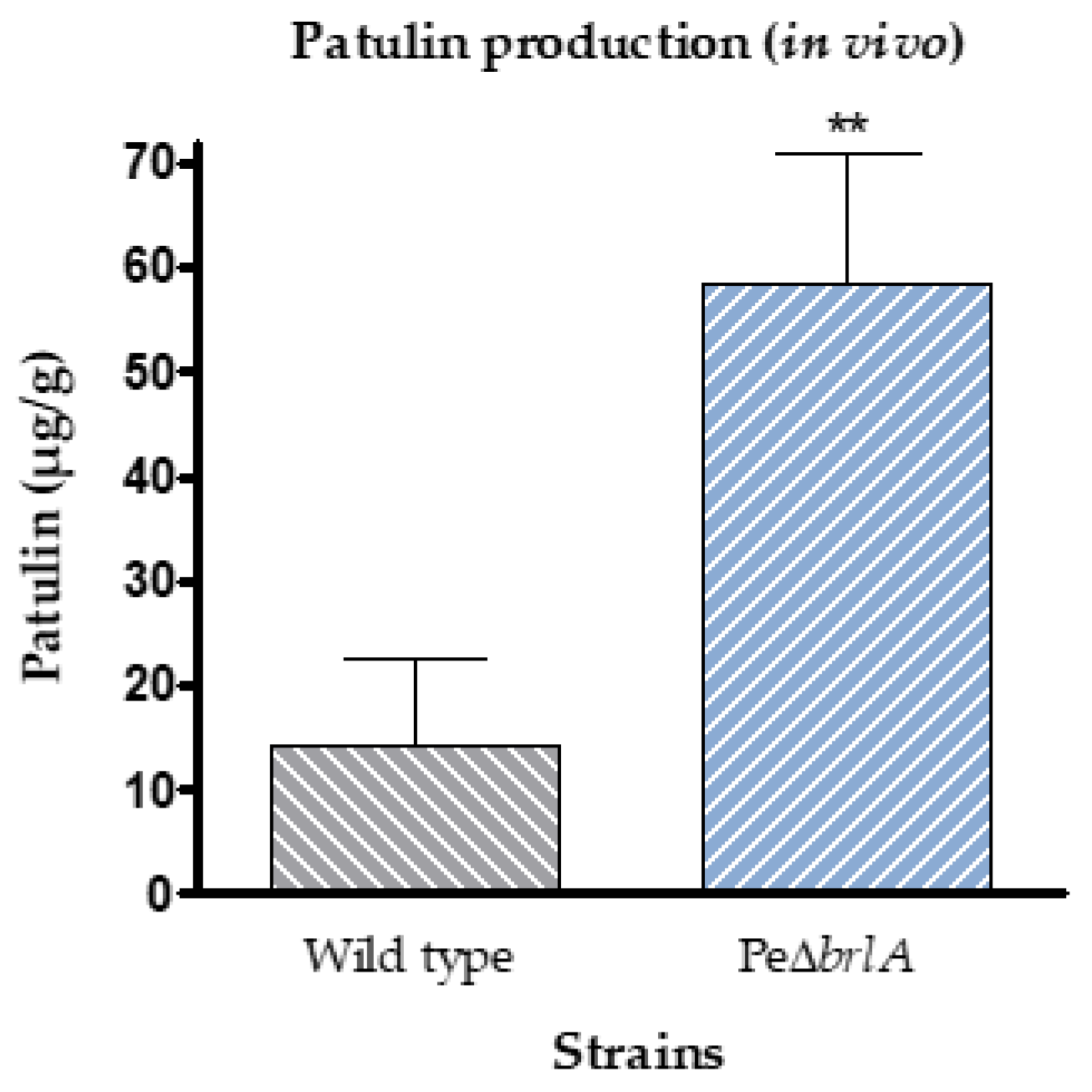
2.2.2. Effect of brlA Deletion on In Vivo Patulin Production
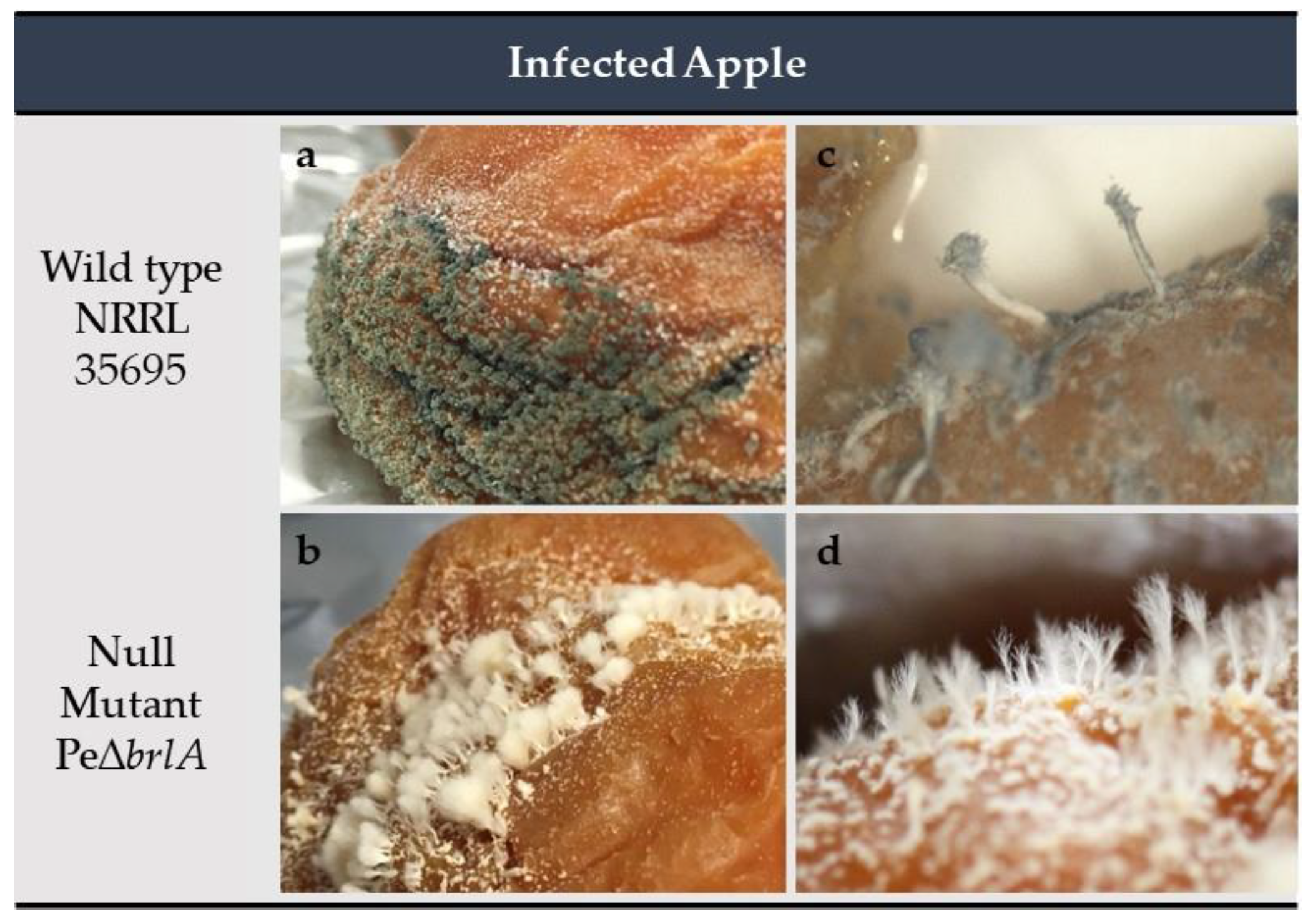
2.2.3. Effect of brlA Deletion on In Vivo Macroscopic Morphology
2.3. Growth Profile in Different Carbon Sources
2.4. BrlA Is a Key Factor in the Regulation of Penicillium expansum Secondary Metabolites
2.4.1. Secondary Metabolites Produced In Vitro
2.4.2. Secondary Metabolites Produced In Vivo
2.5. Analysis of the Transcriptome of PeΔbrlA
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Pe∆brlA Mutant Strain Construction
4.2. Validation of Pe∆brlA Mutant Strain
4.3. Macroscopic and Microscopic Morphology
4.4. Pathogenicity Study and Patulin Production
4.5. Analysis of Growth on Different Carbon Sources
4.6. Secondary Metabolism Study
4.6.1. Fungal Growth Conditions on Labeled Wheats
4.6.2. In Vivo Production of Secondary Metabolites
4.6.3. Analytical Parameters for LC-HRMS
4.6.4. Parameters for High Performance Liquid Chromatography-Diode Array Detector (HPLC-DAD)
4.6.5. Identification of Fungal Metabolites
4.7. Identification of Secondary Metabolites Clusters
4.8. Microarray Gene Expression Studies
4.9. Statistical Analysis of Microarray Data
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APAM | Apple Puree Agar Medium |
| bp | base pair |
| CRP | Central Regulatory Pathway |
| Com | Communesin |
| CYA | Czapek Yeast extract Agar |
| DEG | Differential Expressed Genes |
| DHN | 1,8-Dihydroxynaphthalene |
| dpi | day post inoculation |
| GEO | Gene Expression Omnibus |
| HPLC | High Performance Liquid Chromatography |
| LBG | Locus Bean Gum |
| LC-HRMS | Liquid Chromatography-High Resolution Mass Spectrometry |
| Lhgu | hyphal growth unit Length |
| MEA | Malt Extract agar |
| PDA | Potato Dextrose Agar |
| Rt | Retention time, min |
| SM | Secondary Metabolite |
| TF | Transcription Factor |
| WT | Wild Type |
References
- Cho, H.S.; Hong, S.B.; Go, S.J. First report of Penicillium brasilianum and P. daleae isolated from soil in Korea. Mycobiology 2005, 33, 113–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitt, J.I.; Hocking, A.D. Fungi and food spoilage. In Fungi and Food Spoilage; Springer: Berlin/Heidelberg, Germany, 2009; pp. 243–245. [Google Scholar]
- Houbraken, J.; Kocsubé, S.; Visagie, C.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium: A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Bazioli, J.M.; Amaral, L.S.; Fill, T.; Rodrigues-Filho, E. Insights into Penicillium brasilianum secondary metabolism and its biotechnological potential. Molecules 2017, 22, 858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chain, E.; Florey, H.; Gardner, A.; Heatley, N.; Jennings, M.; Orr-Ewing, J.; Sanders, A. Penicillin as a chemotherapeutic agent. Lancet 1940, 236, 226–228. [Google Scholar] [CrossRef]
- Roberts, E.C.; Cain, C.K.; Muir, R.D.; Reithel, F.J.; Gaby, W.L.; Van Bruggen, J.T.; Homan, D.M.; Katzman, P.A.; Jones, L.R.; Doisy, E.A. Penicillin B, an antibacterial substance from Penicillium notatum. J. Biol. Chem. 1943, 147, 47–58. [Google Scholar]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Schüffler, A.; Anke, T. Fungal natural products in research and development. Nat. Prod. Rep. 2014, 31, 1425–1448. [Google Scholar] [CrossRef]
- Frisvad, J.C. A critical review of producers of small lactone mycotoxins: Patulin, Penicillic acid and moniliformin. World Mycotoxin J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Li, B.; Zong, Y.; Du, Z.L.; Chen, Y.; Zhang, Z.; Qin, G.; Zhao, W.; Tian, S. Genomic characterization reveals insights into patulin biosynthesis and pathogenicity in Penicillium species. Mol. Plant Microbe Interact. 2015, 28, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Filtenborg, O.; Frisvad, J.; Thrane, U. Moulds in food spoilage. Int. J. Food Microbiol. 1996, 33, 85–102. [Google Scholar] [CrossRef]
- Andersen, B.; Smedsgaard, J.; Frisvad, J.C. Penicillium expansum: Consistent production of patulin, chaetoglobosins, and other secondary metabolites in culture and their natural occurrence in fruit products. J. Agric. Food Chem. 2004, 52, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Moss, M. Fungi, quality and safety issues in fresh fruits and vegetables. J. Appl. Microbiol. 2008, 104, 1239–1243. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; Keller, N.P.; Atoui, A.; El Khoury, A.; Lteif, R.; Oswald, I.P.; Puel, O. Secondary metabolism in Penicillium expansum: Emphasis on recent advances in patulin research. Crit. Rev. Food Sci. Nutr. 2017, 58, 2082–2098. [Google Scholar] [CrossRef]
- Baert, K.; Devlieghere, F.; Flyps, H.; Oosterlinck, M.; Ahmed, M.M.; Rajkovic, A.; Verlinden, B.; Nicolai, B.; Debevere, J.; De Meulenaer, B. Influence of storage conditions of apples on growth and patulin production by Penicillium expansum. Int. J. Food Microbiol. 2007, 119, 170–181. [Google Scholar] [CrossRef]
- Moake, M.M.; Padilla-Zakour, O.I.; Worobo, R.W. Comprehensive review of patulin control methods in foods. Compr. Rev. Food Sci. Food Saf. 2005, 4, 8–21. [Google Scholar] [CrossRef]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EC) No 1881/2006 of 19 December Setting Maximum Levels for Certain Contaminants in Foodsteffs. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex:32006R1881 (accessed on 15 January 2020).
- Tannous, J.; El Khoury, R.; Snini, S.P.; Lippi, Y.; El Khoury, A.; Atoui, A.; Lteif, R.; Oswald, I.P.; Puel, O. Sequencing, physical organization and kinetic expression of the patulin biosynthetic gene cluster from Penicillium expansum. Int. J. Food Microbiol. 2014, 189, 51–60. [Google Scholar] [CrossRef]
- Ballester, A.-R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef] [Green Version]
- Snini, S.P.; Tannous, J.; Heuillard, P.; Bailly, S.; Lippi, Y.; Zehraoui, E.; Barreau, C.; Oswald, I.P.; Puel, O. The patulin is a cultivar-dependent aggressiveness factor favoring the colonization of apples by Penicillium expansum. Mol. Plant Pathol. 2016, 17, 920–930. [Google Scholar] [CrossRef]
- Barad, S.; Espeso, E.A.; Sherman, A.; Prusky, D. Ammonia activates pacC and patulin accumulation in an acidic environment during apple colonization by Penicillium expansum. Mol. Plant Pathol. 2015, 17, 727–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannous, J.; Kumar, D.; Sela, N.; Sionov, E.; Prusky, D.; Keller, N.P. Fungal attack and host defence pathways unveiled in near-avirulent interactions of Penicillium expansum creA mutants on apples. Mol. Plant Pathol. 2018, 19, 2635–2650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Barad, S.; Chen, Y.; Luo, X.; Tannous, J.; Dubey, A.; Matana, N.G.; Tian, S.; Li, B.; Keller, N.; et al. LaeA regulation of secondary metabolism modulates virulence in Penicillium expansum and is mediated by sucrose. Mol. Plant Pathol. 2016, 18, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Assaf, C.E.H.; Snini, S.P.; Tadrist, S.; Bailly, S.; Naylies, C.; Oswald, I.P.; Pascal-Lorber, S.; Puel, O. Impact of veA on the development, aggressiveness, dissemination and secondary metabolism of Penicillium expansum. Mol. Plant Pathol. 2018, 19, 1971–1983. [Google Scholar] [CrossRef] [Green Version]
- Tannous, J.; Barda, O.; Luciano-Rosario, D.; Prusky, D.B.; Sionov, E.; Keller, N.P. New insight into pathogenicity and secondary metabolism of the plant pathogen Penicillium expansum through deletion of the epigenetic reader SntB. Front. Microbiol. 2020, 11, 610. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.M.; Theisen, J.M.; Duran, R.M.; Grayburn, W.S.; Calvo, A.M.; Keller, N.P. Secondary metabolism and development is mediated by LlmF control of VeA subcellular localization in Aspergillus nidulans. PLoS Genet. 2013, 9, e1003193. [Google Scholar] [CrossRef] [Green Version]
- Lind, A.L.; Lim, F.Y.; Soukup, A.A.; Keller, N.P.; Rokas, A. An LaeA and BrlA-dependent cellular network governs tissue-specific secondary metabolism in the human pathogen Aspergillus fumigatus. mSphere 2018, 3, e00050-18. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Genet. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Roncal, T.; Ugalde, U. Conidiation induction in Penicillium. Res. Microbiol. 2003, 154, 539–546. [Google Scholar] [CrossRef]
- Boylan, M.T.; Mirabito, P.M.; Willett, C.E.; Zimmerman, C.R.; Timberlake, W.E. Isolation and physical characterization of three essential conidiation genes from Aspergillus nidulans. Mol. Cell. Biol. 1987, 7, 3113–3118. [Google Scholar] [CrossRef]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef]
- Mirabito, P.M.; Adams, T.H.; Timberlake, W.E. Interactions of three sequentially expressed genes control temporal and spatial specificity in Aspergillus development. Cell 1989, 57, 859–868. [Google Scholar] [CrossRef]
- Ni, M.; Yu, J.-H.; Kwon, N.-J.; Shin, K.-S.; Gao, N. Regulation of Aspergillus conidiation. In Cellular and Molecular Biology of Filamentous Fungi; American Society for Microbiology: Washington, DC, USA, 2010; pp. 559–576. [Google Scholar]
- Wang, M.; Sun, X.; Zhu, C.; Xu, Q.; Ruan, R.; Yu, D.; Li, H. PdbrlA, PdabaA and PdwetA control distinct stages of conidiogenesis in Penicillium digitatum. Res. Microbiol. 2015, 166, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-Y.; Mead, M.E.; Lee, M.-K.; Loss, E.M.O.; Kim, S.-C.; Rokas, A.; Yu, J.-H. Systematic dissection of the evolutionarily conserved WetA developmental regulator across a genus of filamentous fungi. mBio 2018, 9, e01130-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, L.; Yu, J.-H. AbaA and WetA govern distinct stages of Aspergillus fumigatus development. Microbiology 2011, 157, 313–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Adams, T. Complex control of the developmental regulatory locus brlA in Aspergillus nidulans. Mol. Genet. Genom. 2001, 266, 260–270. [Google Scholar] [CrossRef]
- Prade, R.; Timberlake, W. The Aspergillus nidulans brlA regulatory locus consists of overlapping transcription units that are individually required for conidiophore development. EMBO J. 1993, 12, 2439–2447. [Google Scholar] [CrossRef]
- Adams, T.H.; Wieser, J.K.; Yu, J.-H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [CrossRef] [Green Version]
- Alkhayyat, F.; Kim, S.C.; Yu, J.-H. Genetic control of asexual development in Aspergillus fumigatus. Adv. Appl. Microbiol. 2015, 90, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Xu, C.; Zhang, Q.; Jiang, B.; Zheng, J.; Jiang, D. C2H2 transcription factor brlA regulating conidiation and affecting growth and biosynthesis of secondary metabolites in Aspergillus clavatus. Int. J. Agric. Biol. 2018, 20, 2549–2555. [Google Scholar]
- Lim, F.Y.; Ames, B.; Walsh, C.T.; Keller, N.P. Co-ordination between BrlA regulation and secretion of the oxidoreductase FmqD directs selective accumulation of fumiquinazoline C to conidial tissues in Aspergillus fumigatus. Cell. Microbiol. 2014, 16, 1267–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.-S.; Kim, Y.-H.; Yu, J.-H. Proteomic analyses reveal the key roles of BrlA and AbaA in biogenesis of gliotoxin in Aspergillus fumigatus. Biochem. Biophys. Res. Commun. 2015, 463, 428–433. [Google Scholar] [CrossRef]
- Qin, Y.; Bao, L.; Gao, M.; Chen, M.; Lei, Y.; Liu, G.; Qu, Y. Penicillium decumbens BrlA extensively regulates secondary metabolism and functionally associates with the expression of cellulase genes. Appl. Microbiol. Biotechnol. 2013, 97, 10453–10467. [Google Scholar] [CrossRef]
- Sigl, C.; Haas, H.; Specht, T.; Pfaller, K.; Kürnsteiner, H.; Zadra, I. Among developmental regulators, StuA but not BrlA is essential for Penicillin V production in Penicillium chrysogenum. Appl. Environ. Microbiol. 2010, 77, 972–982. [Google Scholar] [CrossRef] [Green Version]
- Ojeda-López, M.; Chen, W.; Eagle, C.; Gutiérrez, G.; Jia, W.; Swilaiman, S.; Huang, Z.; Park, H.-S.; Yu, J.-H.; Cánovas, D.; et al. Evolution of asexual and sexual reproduction in the Aspergilli. Stud. Mycol. 2018, 91, 37–59. [Google Scholar] [CrossRef]
- De Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.W.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef] [Green Version]
- Etxebeste, O.; Otamendi, A.; Garzia, A.; Espeso, E.A.; Cortese, M.S. Rewiring of transcriptional networks as a major event leading to the diversity of asexual multicellularity in fungi. Crit. Rev. Microbiol. 2019, 45, 548–563. [Google Scholar] [CrossRef]
- Lin, H.C.; Chiou, G.; Chooi, Y.H.; McMahon, T.C.; Xu, W.; Garg, N.K.; Tang, Y. Elucidation of the concise biosynthetic pathway of the communesin indole alkaloids. Angew. Chem. Int. Ed. Engl. 2015, 54, 3004–3007. [Google Scholar] [CrossRef]
- Fan, Y.; Li, P.-H.; Chao, Y.-X.; Chen, H.; Du, N.; He, Q.-X.; Liu, K.-C. Alkaloids with cardiovascular effects from the marine-derived fungus Penicillium expansum Y32. Mar. Drugs 2015, 13, 6489–6504. [Google Scholar] [CrossRef]
- Hayashi, H.; Matsumoto, H.; Akiyama, K. New insecticidal compounds, communesins C, D and E, from Penicillium expansum link MK-57. Biosci. Biotechnol. Biochem. 2004, 68, 753–756. [Google Scholar] [CrossRef] [Green Version]
- Kerzaon, I.; Pouchus, Y.F.; Monteau, F.; Le Bizec, B.; Grovel, O. Structural investigation and elucidation of new communesins from a marine-derived Penicillium expansum link by liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 3928–3938. [Google Scholar] [CrossRef]
- Numata, A.; Takahashi, C.; Ito, Y.; Takada, T.; Kawai, K.; Usami, Y.; Matsumura, E.; Imachi, M.; Ito, T.; Hasegawa, T. Communesins, cytotoxic metabolites of a fungus isolated from a marine alga. Tetrahedron Lett. 1993, 34, 2355–2358. [Google Scholar] [CrossRef]
- Jadulco, R.; Edrada, R.A.; Ebel, R.; Berg, A.; Schaumann, K.; Wray, V.; Steube, K.; Proksch, P. New communesin derivatives from the fungus Penicillium sp. derived from the mediterranean sponge Axinella verrucosa. J. Nat. Prod. 2004, 67, 78–81. [Google Scholar] [CrossRef]
- Xu, G.-B.; Li, L.-M.; Fang, D.-M.; Li, G.-Y.; Zhang, G.-L.; Wu, Z.-J. Electrospray ionization tandem mass spectrometry of chaetoglobosins. Rapid Commun. Mass Spectrom. 2012, 26, 2115–2122. [Google Scholar] [CrossRef]
- Park, H.-S.; Lee, M.-K.; Kim, S.C.; Yu, J.-H. The role of VosA/VelB-activated developmental gene vadA in Aspergillus nidulans. PLoS ONE 2017, 12, e0177099. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Han, K.-Y.; Kim, K.-J.; Han, D.-M.; Jahng, K.-Y.; Chae, K.-S. The veA gene activates sexual development in Aspergillus nidulans. Fungal Genet. Biol. 2002, 37, 72–80. [Google Scholar] [CrossRef]
- Ni, M.; Yu, J.-H. A novel regulator couples sporogenesis and trehalose biogenesis in Aspergillus nidulans. PLoS ONE 2007, 2, e970. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.-E.; Cho, H.-J.; Chen, W.; Son, S.-H.; Lee, M.-K.; Yu, J.-H.; Park, H.-S. The role of the VosA-repressed dnjA gene in development and metabolism in Aspergillus species. Curr. Genet. 2020, 66, 621–633. [Google Scholar] [CrossRef]
- Gauthier, T.; Wang, X.; Dos Santos, J.S.; Fysikopoulos, A.; Tadrist, S.; Canlet, C.; Artigot, M.P.; Loiseau, N.; Oswald, I.P.; Puel, O. Trypacidin, a spore-borne toxin from Aspergillus fumigatus, is cytotoxic to lung cells. PLoS ONE 2012, 7, e29906. [Google Scholar] [CrossRef]
- Blachowicz, A.; Raffa, N.; Bok, J.W.; Choera, T.; Knox, B.; Lim, F.Y.; Huttenlocher, A.; Wang, C.C.C.; Venkateswaran, K.; Keller, N.P. Contributions of spore secondary metabolites to UV-C protection and virulence vary in different Aspergillus fumigatus strains. mBio 2020, 11, e03415-19. [Google Scholar] [CrossRef] [Green Version]
- Ishiuchi, K.; Nakazawa, T.; Yagishita, F.; Mino, T.; Noguchi, H.; Hotta, K.; Watanabe, K. Combinatorial generation of complexity by redox enzymes in the chaetoglobosin A biosynthesis. J. Am. Chem. Soc. 2013, 135, 7371–7377. [Google Scholar] [CrossRef]
- Qiao, K.; Chooi, Y.-H.; Tang, Y. Identification and engineering of the cytochalasin gene cluster from Aspergillus clavatus NRRL 1. Metab. Eng. 2011, 13, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Baltussen, T.J.H.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular mechanisms of conidial germination in Aspergillus spp. Microbiol. Mol. Biol. Rev. 2019, 84, e00049-19. [Google Scholar] [CrossRef]
- Van Munster, J.M.; Nitsche, B.M.; Akeroyd, M.; Dijkhuizen, L.; Van Der Maarel, M.J.E.C.; Ram, A.F.J. Systems approaches to predict the functions of glycoside hydrolases during the life cycle of Aspergillus niger using developmental mutants ∆brlA and ∆flbA. PLoS ONE 2015, 10, e0116269. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, S.; Torres, G.; Lin, X. Laccases involved in 1,8-dihydroxynaphthalene melanin biosynthesis in Aspergillus fumigatus are regulated by developmental factors and copper homeostasis. Eukaryot. Cell 2013, 12, 1641–1652. [Google Scholar] [CrossRef] [Green Version]
- Perez-Cuesta, U.; Aparicio-Fernandez, L.; Guruceaga, X.; Martin-Souto, L.; Abad-Diaz-De-Cerio, A.; Antoran, A.; Buldain, I.; Hernando, F.L.; Ramirez-Garcia, A.; Rementeria, A. Melanin and pyomelanin in Aspergillus fumigatus: From its genetics to host interaction. Int. Microbiol. 2019, 23, 55–63. [Google Scholar] [CrossRef]
- Si, H.; Rittenour, W.R.; Xu, K.; Nicksarlian, M.; Calvo, A.M.; Harris, S.D. Morphogenetic and developmental functions of the Aspergillus nidulans homologues of the yeast bud site selection proteins Bud4 and Axl2. Mol. Microbiol. 2012, 85, 252–270. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-S.; Yu, J.-H. Developmental regulators in Aspergillus fumigatus. J. Microbiol. 2016, 54, 223–231. [Google Scholar] [CrossRef]
- Kim, M.-J.; Jung, W.-H.; Son, Y.-E.; Yu, J.-H.; Lee, M.-K.; Park, H.-S. The velvet repressed vidA gene plays a key role in governing development in Aspergillus nidulans. J. Microbiol. 2019, 57, 893–899. [Google Scholar] [CrossRef]
- Son, Y.-E.; Cho, H.-J.; Lee, M.-K.; Park, H.-S. Characterizing the role of Zn cluster family transcription factor ZcfA in governing development in two Aspergillus species. PLoS ONE 2020, 15, e0228643. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Wang, L.; Lee, H.; Frisvad, J.C. New sections in Penicillium containing novel species producing patulin, pyripyropens or other bioactive compounds. Persoonia 2016, 36, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Steenwyk, J.L.; Shen, X.-X.; Lind, A.L.; Goldman, G.H.; Rokas, A. A robust phylogenomic time tree for biotechnologically and medically important fungi in the genera Aspergillus and Penicillium. mBio 2019, 10, e00925-19. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, D.; Suzuki, S.; Kamei, K.; Gonoi, T.; Kawamoto, S.; Hagiwara, D. The role of AtfA and HOG MAPK pathway in stress tolerance in conidia of Aspergillus fumigatus. Fungal Genet. Biol. 2014, 73, 138–149. [Google Scholar] [CrossRef]
- Mortensen, E.M.; McDonald, H.; Yates, J.; Kellogg, D.R. Cell cycle-dependent assembly of a Gin4-septin complex. Mol. Biol. Cell 2002, 13, 2091–2105. [Google Scholar] [CrossRef]
- Vargas-Muñiz, J.M.; Renshaw, H.; Richards, A.D.; Waitt, G.; Soderblom, E.J.; Moseley, M.A.; Asfaw, Y.; Juvvadi, P.R.; Steinbach, W.J. Dephosphorylation of the core septin, AspB, in a protein phosphatase 2A-dependent manner impacts its localization and function in the fungal pathogen Aspergillus fumigatus. Front. Microbiol. 2016, 7, 997. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef]
- Sanzani, S.; Reverberi, M.; Punelli, M.; Ippolito, A.; Fanelli, C. Study on the role of patulin on pathogenicity and virulence of Penicillium expansum. Int. J. Food Microbiol. 2012, 153, 323–331. [Google Scholar] [CrossRef]
- Sekiguchi, J.; Gaucher, G.M. Conidiogenesis and secondary metabolism in Penicillium urticae. Appl. Environ. Microbiol. 1977, 33, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Valente, S.; Cometto, A.; Piombo, E.; Meloni, G.R.; Ballester, A.-R.; González-Candelas, L.; Spadaro, D. Elaborated regulation of griseofulvin biosynthesis in Penicillium griseofulvum and its role on conidiation and virulence. Int. J. Food Microbiol. 2020, 328, 108687. [Google Scholar] [CrossRef] [PubMed]
- Reiß, J. Development of Aspergillus parasiticus and formation of aflatoxin B1 under the influence of conidiogenesis affecting compounds. Arch. Microbiol. 1982, 133, 236–238. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, H.; Sumarah, M.; Gao, Q.; Wang, D.; Zhang, Y. veA gene acts as a positive regulator of conidia production, ochratoxin a biosynthesis, and oxidative stress tolerance in Aspergillus niger. J. Agric. Food Chem. 2018, 66, 13199–13208. [Google Scholar] [CrossRef]
- Satterlee, T.; Nepal, B.; Lorber, S.; Puel, O.; Calvo, A.M. The transcriptional regulator HbxA governs development, secondary metabolism, and virulence in Aspergillus fumigatus. Appl. Environ. Microbiol. 2019, 86, e01779-19. [Google Scholar] [CrossRef]
- Lee, B.N.; Adams, T.H. The Aspergillus nidulans fluG gene is required for production of an extracellular developmental signal and is related to prokaryotic glutamine synthetase I. Genes Dev. 1994, 8, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Wieser, J.; Na Lee, B.; Fondon, J.W.; Adams, T.H. Genetic requirements for initiating asexual development in Aspergillus nidulans. Curr. Genet. 1994, 27, 62–69. [Google Scholar] [CrossRef]
- Rodríguez-Urra, A.B.; Jiménez, C.; Nieto, M.I.; Rodríguez, J.; Hayashi, H.; Ugalde, U. Signaling the induction of sporulation involves the interaction of two secondary metabolites in Aspergillus nidulans. ACS Chem. Biol. 2012, 7, 599–606. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.; Guo, Q.; Yu, W.; Zhang, Y.; He, B. Bioactivities and Future Perspectives of Chaetoglobosins. Evid. Based Complement. Altern. Med. 2020, 2020, e8574084-10. [Google Scholar] [CrossRef] [Green Version]
- Coyle, C.M.; Kenaley, S.C.; Rittenour, W.R.; Panaccione, D.G. Association of ergot alkaloids with conidiation in Aspergillus fumigatus. Mycologia 2007, 99, 804–811. [Google Scholar] [CrossRef]
- Schumann, J.; Hertweck, C. Molecular basis of cytochalasan biosynthesis in fungi: Gene cluster analysis and evidence for the involvement of a PKS-NRPS hybrid synthase by RNA silencing. J. Am. Chem. Soc. 2007, 129, 9564–9565. [Google Scholar] [CrossRef]
- Qi, J.; Jiang, L.; Zhao, P.; Chen, H.; Jia, X.; Zhao, L.; Dai, H.; Hu, J.; Liu, C.; Shim, S.H.; et al. Chaetoglobosins and azaphilones from Chaetomium globosum associated with Apostichopus japonicus. Appl. Microbiol. Biotechnol. 2020, 104, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Twumasi-Boateng, K.; Yu, Y.; Chen, D.; Gravelat, F.N.; Nierman, W.C.; Sheppard, D.C. Transcriptional profiling identifies a role for BrlA in the response to nitrogen depletion and for StuA in the regulation of secondary metabolite clusters in Aspergillus fumigatus. Eukaryot. Cell 2008, 8, 104–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punt, P.J.; Oliver, R.P.; Dingemanse, M.A.; Pouwels, P.H.; Hondel, C.A.V.D. Transformation of Aspergillus based on the hygromycin B resistance marker from Escherichia coli. Gene 1987, 56, 117–124. [Google Scholar] [CrossRef]
- Shevchuk, N.A.; Bryksin, A.V.; Nusinovich, Y.A.; Cabello, F.C.; Sutherland, M.; Ladisch, S. Construction of long DNA molecules using long PCR-based fusion of several fragments simultaneously. Nucleic Acids Res. 2004, 32, e19. [Google Scholar] [CrossRef]
- Lim, F.Y.; Sanchez, J.F.; Wang, C.C.; Keller, N.P. Toward awakening cryptic secondary metabolite gene clusters in filamentous fungi. Enzym. Eng. Evol. Gen. Methods 2012, 517, 303–324. [Google Scholar] [CrossRef] [Green Version]
- Adjovi, Y.; Bailly, S.; Gnonlonfin, B.; Tadrist, S.; Querin, A.; Sanni, A.; Oswald, I.P.; Puel, O.; Bailly, J. Analysis of the contrast between natural occurrence of toxigenic Aspergilli of the Flavi section and aflatoxin B1 in cassava. Food Microbiol. 2014, 38, 151–159. [Google Scholar] [CrossRef]
- De Vries, R.P.; Burgers, K.; Van De Vondervoort, P.J.I.; Frisvad, J.C.; Samson, R.A.; Visser, J. A new black Aspergillus species, A. vadensis, is a promising host for homologous and heterologous protein production. Appl. Environ. Microbiol. 2004, 70, 3954–3959. [Google Scholar] [CrossRef] [Green Version]
- Meijer, M.; Houbraken, J.; Dalhuijsen, S.; Samson, R.A.; De Vries, R.P. Growth and hydrolase profiles can be used as characteristics to distinguish Aspergillus niger and other black Aspergilli. Stud. Mycol. 2011, 69, 19–30. [Google Scholar] [CrossRef]
- Pean, M.; Boiry, S.; Ferrandi, J.-C.; Gibiat, F.; Puel, O.; Delaforge, M. Production and use of mycotoxins uniformly enriched with stable isotopes for their dosage in biological samples: (1) production of uniformly enriched biomass. J. Label. Compd. Radiopharm. 2007, 50, 569–570. [Google Scholar] [CrossRef]
- Hautbergue, T.; Puel, O.; Tadrist, S.; Meneghetti, L.; Pean, M.; Delaforge, M.; Debrauwer, L.; Oswald, I.P.; Jamin, E.L. Evidencing 98 secondary metabolites of Penicillium verrucosum using substrate isotopic labeling and high-resolution mass spectrometry. J. Chromatogr. B 2017, 1071, 29–43. [Google Scholar] [CrossRef]
- Macdonald, S.; Long, M.; Gilbert, J.; Felgueiras, I. Liquid chromatographic method for determination of patulin in clear and cloudy apple juices and apple puree: Collaborative study. J. AOAC Int. 2000, 83, 1387–1394. [Google Scholar] [PubMed]
- Cano, P.M.; Jamin, E.L.; Tadrist, S.; Bourdaud’Hui, P.; Pean, M.; Debrauwer, L.; Oswald, I.P.; Delaforge, M.; Puel, O. New untargeted metabolic profiling combining mass spectrometry and isotopic labeling: Application on Aspergillus fumigatus grown on wheat. Anal. Chem. 2013, 85, 8412–8420. [Google Scholar] [CrossRef] [PubMed]
- Laatsch, H. AntiBase: The Natural Compound Identifier; Wiley-VCH: Weinheim, Germany, 2012. [Google Scholar]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannous, J.; Canlet, C.; Pinton, P.; Lippi, Y.; Alassane-Kpembi, I.; Gauthier, T.; Atoui, A.; Zhou, T.; Lteif, R.; Snini, S.P.; et al. Patulin transformation products and last intermediates in its biosynthetic pathway, E and Z-ascladiol, are not toxic to human cells. Arch. Toxicol. 2016, 91, 2455–2467. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A. Gene expression omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.D.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef]
- Hicks, S.C.; Okrah, K.; Paulson, J.N.; Quackenbush, J.; Irizarry, R.A.; Bravo, H.C. Smooth quantile normalization. Biostatistics 2017, 19, 185–198. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Formula | 12C m/z (Da) | a RT (min) | Proposed Identification | WT | Pe∆brlA | Molecular Formula | 12C m/z (Da) | a RT (min) | Proposed Identification | WT | Pe∆brlA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C7H6O4 | 153.01919 | 3.59 | Patulin † | + | + | C28H32N4O | 441.26589 | 17.14 | Communesin F d | + | ND |
| C7H8O | 109.06509 | 7.11 | m-Cresol † | + | + | C28H32N4O2 | 457.26116 | 23.65 | Communesin A f,† | + | + |
| C7H8O2 | 125.05998 | 6.79 | m-Hydroxybenzyl alcohol † | + | + | C28H32N4O3 | 473.25566 | 15.50 | + | ND | |
| C7H8O3 | 141.05493 | 3.80 | Gentisyl alcohol † | + | + | C28H36N4O4 | 493.27971 | 31.47 | Fungisporin A or cyclo(VFVF) | + | + |
| C7H8O4 | 157.04990 | 2.67 | Ascladiol † | + | + | C28H38N4O5 | 511.29099 | 18.01 | VAL-PHE-VAL-PHE | + | + |
| C7H10O3 | 143.07061 | 4.52 | + | + | C28H38N4O6 | 527.28755 | 12.46 | VAL-PHE-VAL-TYR | + | + | |
| C10H17NO5 | 232.11872 | 8.83 | + | + | C28H38O7 | 485.25405 | 35.32 | + | + | ||
| C10H17NO5 | 232.11872 | 9.65 | + | ND | C28H38O7 | 487.27063 | 36.21 | Andrastin A † | + | + | |
| C13H14O5 | 251.09108 | 22.13 | Citrinin † | + | + | C28H38O8 | 501.24792 | 27.70 | + | + | |
| C15H18N2 | 227.15514 | 6.44 | Aurantioclavine † | + | ND | C28H38O8 | 501.24792 | 28.44 | + | ND | |
| C15H19NO6 | 310.12939 | 12.78 | + | + | C28H38O8 | 501.24786 | 29.57 | + | + | ||
| C15H19NO6 | 310.12964 | 14.80 | + | + | C28H40O6 | 471.27434 | 39.48 | Andrastin C | + | + | |
| C15H20O4 | 265.14412 | 15.91 | Expansolide C/D | + | + | C28H40O7 | 487.26898 | 30.58 | Andrastin B | + | + |
| C15H20O4 | 265.14410 | 16.91 | + | ND | C29H27N5O5 | 526.20693 | 15.70 | ND | + | ||
| C15H20O4 | 265.14415 | 18.49 | Expansolide C/D | + | + | C29H27N5O5 | 526.20689 | 17.61 | ND | + | |
| C15H20O4 | 265.14414 | 19.35 | + | + | C29H31N5O5 | 530.23837 | 14.83 | ND | + | ||
| C16H18N2O2 | 271.14496 | 7.62 | Clavicipitic acid † | + | + | C29H31N5O5 | 530.23812 | 15.85 | ND | + | |
| C16H18N2O2 | 271.14467 | 8.45 | Clavicipitic acid † | + | + | C29H31N5O5 | 530.23840 | 16.68 | ND | + | |
| C16H26N2O4S2 | 375.14202 | 22.02 | + | + | C29H33N5O6 | 548.24883 | 11.36 | ND | + | ||
| C17H22O5 | 307.15471 | 27.39 | Expansolide A/B | + | + | C29H33N5O6 | 548.24860 | 12.14 | ND | + | |
| C17H22O5 | 307.15504 | 30.19 | Expansolide A/B | + | + | C29H33N5O6 | 548.25180 | 15.42 | + | ND | |
| C17H23N3O3 | 318.17943 | 3.12 | ND | + | C31H36N4O2 | 497.29061 | 31.66 | Putative new undetermined communesin | + | ND | |
| C17H23N3O3 | 318.18043 | 25.05 | ND | + | C32H34N4O3 | 523.27152 | 31.84 | Communesin D g | + | + | |
| C18H16N2O2 | 293.12911 | 13.58 | + | ND | C32H36N2O4 | 513.27635 | 39.10 | Chaetoglobosin J or Prochaetoglobosin III | + | + | |
| C18H16N2O2 | 293.12915 | 17.74 | + | ND | C32H36N2O5 | 529.26776 | 20.92 | Chaetoglobosin B/G | + | + | |
| C18H16N2O3 | 309.12421 | 14.76 | + | ND | C32H36N2O5 | 529.26807 | 23.19 | Chaetoglobosin B/G | ND | + | |
| C18H18N2O2 | 295.14479 | 14.61 | + | + | C32H36N2O5 | 529.27058 | 25.21 | Chaetoglobosin B/G | + | + | |
| C18H31NO7 | 374.21663 | 23.72 | + | + | C32H36N2O5 | 529.27029 | 26.71 | Chaetoglobosin B/G | + | + | |
| C18H35N3O4 | 358.26901 | 3.54 | ND | + | C32H36N2O5 | 529.26757 | 29.56 | Chaetoglobosin B/G | + | + | |
| C19H16N2O2 | 305.12911 | 32.50 | + | ND | C32H36N2O5 | 529.27077 | 30.51 | Chaetoglobosin B/G | + | + | |
| C19H16N2O2 | 305.12933 | 33.17 | + | ND | C32H36N2O5 | 529.26807 | 32.43 | Chaetoglobosin B/G | ND | + | |
| C19H16N2O4 | 337.11909 | 13.73 | + | ND | C32H36N2O5 | 529.27067 | 33.36 | Chaetoglobosin A † | + | + | |
| C19H16N2O4 | 337.11916 | 15.20 | + | ND | C32H36N2O5 | 529.26771 | 34.25 | Chaetoglobosin B/G | ND | + | |
| C19H20O5 | 329.13738 | 33.29 | ND | + | C32H36N2O5 | 529.26769 | 35.49 | Chaetoglobosin B/G | ND | + | |
| C19H21NO7 | 376.13901 | 17.45 | + | + | C32H36N2O5 | 529.27063 | 36.41 | Chaetoglobosin C † | + | + | |
| C19H21NO7 | 376.13904 | 18.65 | + | + | C32H36N2O5 | 529.27056 | 37.41 | Chaetoglobosin B/G | + | + | |
| C19H38O6 | 361.25871 | 37.23 | + | ND | C32H36N2O5 | 529.26814 | 38.29 | Chaetoglobosin B/G | ND | + | |
| C19H38O6 | 361.25819 | 38.19 | + | ND | C32H36N2O6 | 545.26332 | 27.60 | Putative cytochalasan | + | + | |
| C20H18N2O2 | 319.14588 | 35.48 | + | ND | C32H36N2O6 | 545.26241 | 29.23 | Putative cytochalasan | ND | + | |
| C20H18N2O2 | 319.14502 | 36.44 | + | ND | C32H36N2O6 | 545.26288 | 30.57 | Putative cytochalasan | + | + | |
| C20H21NO9 | 420.12857 | 19.29 | + | + | C32H36N2O6 | 545.26281 | 31.50 | Putative cytochalasan | + | + | |
| C20H21NO9 | 420.12858 | 22.19 | ND | + | C32H36N2O6 | 545.26294 | 32.48 | Putative cytochalasan | ND | + | |
| C20H26O8 | 395.17142 | 13.61 | + | + | C32H36N2O6 | 543.24836 | 34.05 | Putative cytochalasan | + | + | |
| C22H23N5O2 | 390.19390 | 15.09 | Roquefortine C † | + | + | C32H36N4O2 | 509.29257 | 34.95 | Communesin B g,† | + | + |
| C22H25N5O2 | 392.20913 | 9.99 | Roquefortine D | + | + | C32H38N2O4 | 515.28895 | 35.59 | Putative cytochalasan | + | + |
| C23H24N2O6 | 425.17179 | 26.92 | + | + | C32H38N2O4 | 515.28890 | 37.93 | Putative cytochalasan | + | + | |
| C24H26N2O6 | 437.17091 | 33.79 | + | ND | C32H38N2O4 | 515.28909 | 38.15 | Putative cytochalasan | + | + | |
| C26H30N4 | 399.25568 | 19.94 | Communesin K b | + | ND | C32H38N2O5 | 531.28384 | 21.75 | Putative cytochalasan | ND | + |
| C26H30N4O | 415.25034 | 14.62 | Communesin I c | + | + | C32H38N2O5 | 531.28990 | 22.76 | Putative cytochalasan | + | + |
| C26H30N4O | 415.25033 | 18.43 | Communesin I c | + | + | C32H38N2O5 | 531.28431 | 23.91 | Putative cytochalasan | ND | + |
| C26H32O8 | 473.21511 | 9.67 | ND | + | C32H38N2O5 | 531.28360 | 27.38 | Putative cytochalasan | + | + | |
| C26H32O8 | 473.21552 | 10.64 | ND | + | C32H38N2O5 | 531.28354 | 28.02 | Chaetoglobosin E h | ND | + | |
| C26H32O8 | 473.21489 | 34.14 | ND | + | C32H38N2O5 | 531.28340 | 28.52 | Putative cytochalasan | + | + | |
| C26H40O6 | 449.28939 | 29.93 | + | + | C32H38N2O5 | 531.28338 | 31.56 | Penochalasin F h | ND | + | |
| C27H29N5O4 | 488.22783 | 10.80 | ND | + | C32H38N2O6 | 547.27864 | 23.19 | Putative cytochalasan | ND | + | |
| C27H29N5O4 | 488.22766 | 11.03 | ND | + | C32H38N2O6 | 547.27841 | 25.47 | Putative cytochalasan | ND | + | |
| C27H30N4O2 | 443.24564 | 15.03 | Communesin E d | + | + | C32H38N2O6 | 547.27842 | 27.09 | Putative cytochalasan | ND | + |
| C28H30N4O3 | 471.23997 | 19.56 | Com470 e | + | + | C32H38N2O6 | 547.27851 | 28.27 | Putative cytochalasan | ND | + |
| C28H31N5O5 | 518.24108 | 16.36 | + | + | C33H38N4O5 | 571.29324 | 18.22 | Com570 e | + | ND | |
| C28H31N5O5 | 518.24092 | 17.21 | + | + | C37H42N4O5 | 623.32511 | 29.52 | Com622 e | + | ND |
| Molecular Formula | 12C m/z (Da) | RT (min) a | Proposed Identification | Molecular Formula | 12C m/z (Da) | RT (min) a | Proposed Identification |
|---|---|---|---|---|---|---|---|
| C7H6O4 | 153.01919 | 3.59 | Patulin | C29H27N5O5 | 526.20689 | 17.61 | |
| C7H8O3 | 141.05493 | 3.80 | Gentisyl alcohol | C32H36N2O4 | 513.27635 | 39.10 | Chaetoglobosin J or Prochaetoglobosin III |
| C7H8O4 | 157.04990 | 2.67 | Ascladiol | C32H36N2O5 | 529.26776 | 20.92 | Chaetoglobosin B/G |
| C10H17NO5 | 232.11872 | 8.83 | C32H36N2O5 | 529.26807 | 23.19 | Chaetoglobosin B/G | |
| C13H14O5 | 251.09108 | 21.70 | Citrinin | C32H36N2O5 | 529.27029 | 26.71 | Chaetoglobosin B/G |
| C16H26N2O4S2 | 375.14202 | 22.02 | C32H36N2O5 | 529.26898 | 29.54 | Chaetoglobosin B/G | |
| C17H22O5 | 307.15471 | 27.39 | Expansolide A/B | C32H36N2O5 | 529.27539 | 30.13 | Chaetoglobosin B/G |
| C17H22O5 | 307.15504 | 30.19 | Expansolide A/B | C32H36N2O5 | 529.26807 | 32.43 | Chaetoglobosin B/G |
| C19H21NO7 | 376.13901 | 17.45 | C32H36N2O5 | 529.26769 | 35.49 | Chaetoglobosin B/G | |
| C19H21NO7 | 376.13904 | 18.65 | C32H36N2O5 | 529.27056 | 37.41 | Chaetoglobosin B/G | |
| C22H23N5O2 | 390.19390 | 15.09 | Roquefortine C | C32H36N4O2 | 509.29257 | 36.01 | Communesin B |
| C23H24N2O6 | 425.17179 | 26.92 | C32H38N2O5 | 531.28384 | 21.75 | Putative cytochalasan | |
| C28H38O7 | 485.25405 | 35.32 | C32H38N2O5 | 531.28431 | 23.91 | Putative cytochalasan | |
| C28H38O7 | 487.27063 | 36.21 | Andrastin A | C32H38N2O6 | 547.27864 | 23.19 | Putative cytochalasan |
| C28H38O8 | 501.24792 | 28.44 | C32H38N2O6 | 547.27841 | 25.47 | Putative cytochalasan | |
| C28H38O8 | 501.24786 | 29.57 | C32H38N2O6 | 547.27842 | 27.09 | Putative cytochalasan | |
| C28H40O7 | 487.26898 | 30.58 | Andrastin B |
| Molecular Formula | 12C m/z (Da) | a RT (min) | Proposed Identification | Molecular Formula | 12C m/z (Da) | a RT (min) | Proposed Identification |
|---|---|---|---|---|---|---|---|
| C15H20O4 | 265.14412 | 15.91 | Expansolide C/D | C28H38O8 | 501.24786 | 29.57 | |
| C15H20O4 | 265.14415 | 18.49 | Expansolide C/D | C28H40O6 | 471.27434 | 39.48 | Andrastin C |
| C16H18N2O2 | 271.14496 | 7.62 | Clavicipitic acid | C28H40O7 | 487.26898 | 30.58 | Andrastin B |
| C16H26N2O4S2 | 375.14202 | 22.02 | C29H33N5O6 | 548.25180 | 14.66 | ||
| C17H22O5 | 307.15471 | 27.39 | Expansolide A/B | C32H34N4O3 | 523.27152 | 31.84 | Communesin D |
| C17H22O5 | 307.15504 | 30.19 | Expansolide A/B | C32H36N2O4 | 513.27635 | 39.10 | Chaetoglobosin J or Prochaetoglobosin III |
| C18H16N2O2 | 293.12915 | 17.74 | C32H36N2O5 | 529.27029 | 26.79 | Chaetoglobosin B/G | |
| C18H18N2O2 | 295.14479 | 14.61 | C32H36N2O5 | 529.26757 | 29.56 | Chaetoglobosin B/G | |
| C19H38O6 | 361.25819 | 38.19 | C32H36N2O5 | 529.27077 | 30.51 | Chaetoglobosin B/G | |
| C22H20N3O | 341.15380 | 38.56 | C32H36N2O5 | 529.27067 | 33.36 | Chaetoglobosin A | |
| C22H20N3O | 341.15368 | 39.29 | C32H36N2O5 | 529.26769 | 35.49 | Chaetoglobosin B/G | |
| C22H23N5O2 | 390.19390 | 15.09 | Roquefortine C | C32H36N2O5 | 529.27063 | 36.74 | Chaetoglobosin C |
| C22H25N5O2 | 392.20913 | 9.99 | Roquefortine D | C32H36N2O5 | 529.27056 | 37.41 | Chaetoglobosin B/G |
| C23H24N2O6 | 425.17179 | 26.92 | C32H36N2O6 | 545.26332 | 27.60 | Putative cytochalasan | |
| C24H26N2O6 | 437.17091 | 33.79 | C32H36N2O6 | 545.26241 | 29.23 | Putative cytochalasan | |
| C26H30N4 | 399.25568 | 19.94 | Communesin K | C32H36N2O6 | 545.26288 | 30.57 | Putative cytochalasan |
| C26H30N4O | 415.25034 | 14.62 | Communesin I | C32H36N2O6 | 545.26294 | 32.48 | Putative cytochalasan |
| C26H30N4O | 415.25033 | 18.43 | Communesin I | C32H36N4O2 | 509.29257 | 34.95 | Communesin B |
| C26H40O6 | 449.28939 | 29.93 | C32H38N2O4 | 515.28895 | 35.59 | Putative cytochalasan | |
| C27H30N4O2 | 443.24564 | 15.65 | Communesin E | C32H38N2O4 | 515.28890 | 37.93 | Putative cytochalasan |
| C28H30N4O3 | 471.23997 | 19.56 | Com470 | C32H38N2O4 | 515.28909 | 38.15 | Putative cytochalasan |
| C28H32N4O | 441.26589 | 17.14 | Communesin F | C32H38N2O5 | 531.28990 | 22.76 | Putative cytochalasan |
| C28H32N4O2 | 457.26116 | 23.65 | Communesin A | C32H38N2O5 | 531.28431 | 23.91 | Putative cytochalasan |
| C28H38O7 | 485.25405 | 35.32 | C32H38N2O5 | 531.28354 | 28.02 | Chaetoglobosin E | |
| C28H38O7 | 487.27063 | 36.21 | Andrastin A | C32H38N2O5 | 531.28338 | 31.56 | Penochalasin |
| C28H38O8 | 501.24792 | 27.70 | C32H38N2O6 | 547.27864 | 23.19 | Putative cytochalasan | |
| C28H38O8 | 501.24792 | 28.44 | C32H38N2O6 | 547.27841 | 25.47 | Putative cytochalasan |
| Penicillium expansum d1 Strain Gene ID | Protein Name | Log2 Fold Change PeΔbrlA vs. WT | Adjusted p-Value | Putative Role | |
|---|---|---|---|---|---|
| Regulation of Development | PEXP_029020 | AbaA | −3.62 | 4.07 × 10−11 | Transcription factor |
| PEXP_077410 | WetA | −3.82 | 1.02 × 10−10 | DNA-binding transcription factor | |
| PEXP_085800 | Axl2 | −2.51 | 5.36 × 10−11 | Phialide morphogenesis regulatory protein | |
| PEXP_040110 | PhiA | 1.83 | 1.73 × 10−5 | Phialide development protein | |
| PEXP_003940 | VadA | −2.83 | 1.37 × 10−9 | Spore-specific regulator | |
| PEXP_102520 | DnjA | 1.33 | 2.75 × 10−7 | DnaJ familly chaperone | |
| PEXP_064110 | MedA | 1.13 | 3.51 × 10−5 | Temporal modifier of developmental | |
| PEXP_050580 | PpoC | 1.29 | 6.05 × 10−6 | psi-Producing oxygenase | |
| Hydrophobins | PEXP_062290 | RodA | −13.00 | 1.07 × 10−18 | Rodlet A, Hydrophobic protein |
| PEXP_020490 | RodB/DewB | −11.4 | 1.43 × 10−16 | Rodlet B, Hydrophobic protein | |
| PEXP_071760 | DewC | −0.546 | 2.14 × 10−1 | ||
| PEXP_043320 | DewD | −5.62 | 5.94 × 10−13 | ||
| PEXP_098360 | DewE | −0.906 | 2.41 × 10−3 | ||
| Pigmentation DHN-Melanin Like Pigment | PEXP_096630 | Alb1 | −12.2 | 6.41 × 10−17 | Putative polyketide synthase |
| PEXP_097170 | Arp1 | −8.64 | 1.31 × 10−15 | Putative protein-Conidial pigmentation | |
| PEXP_097180 | Arp2 | −8.34 | 1.13 × 10−13 | HN reductase | |
| PEXP_097190 | Ayg1 | −6.57 | 9.55 × 10−13 | ||
| PEXP_097110 | Abr1 | −6.57 | 1.58 × 10−12 | Multicopper oxidase | |
| Trehalose Biosynthesis | PEXP_050560 | Ccg-9 | −5.04 | 2.47 × 10−6 | Clock-controlled gene 9 |
| Kinase | PEXP_066390 | Gin4 | −5.00 | 2.02 × 10−12 | Localization and function of septins |
| Velvet Protein Family | PEXP_092360 | VeA | 0.89 | 1.05 × 10−5 | Global transcription factor |
| PEXP_065290 | VelB | −0.53 | 1.01 × 10−3 | Velvet-like protein B | |
| PEXP_009420 | VelC | 0.43 | 3.66 × 10−4 | Regulator of sexual development | |
| PEXP_042660 | LaeA | −0.44 | 5.39 × 10−3 | Putative methyltransferase | |
| PEXP_076870 | VosA | −0.98 | 2.35 × 10−4 | Multifunctional regulator of development |
| Penicillium expansum Strain d1 Gene ID | Biosynthetic Gene Cluster | Log2 Fold Change PeΔbrlA vs. WT | Adjusted p-Value | |
|---|---|---|---|---|
| DMATS (Dimethylallyl tryptophane synthase) | PEXP_030140 | Roquefortine C | 1.68 | 1.40 × 10−5 |
| PEXP_030510 | Communesins | −4.64 | 3.08 × 10−10 | |
| PEXP_058590 | - | −1.31 | 2.72 × 10−8 | |
| PKS (Polyketide synthase) | PEXP_006700 | - | −4.51 | 5.61 × 10−15 |
| PEXP_028920 | - | 4.51 | 1.77 × 10−4 | |
| PEXP_030540 | Communesins | −1.76 | 5.50 × 10−7 | |
| PEXP_037250 | - | −2.14 | 4.85 × 10−8 | |
| PEXP_063170 | - | 2.73 | 1.29 × 10−2 | |
| PEXP_076200 | - | −1.44 | 4.74 × 10−5 | |
| PEXP_094460 | Patulin | −1.03 | 1.21 × 10−5 | |
| PEXP_094770 | - | 1.12 | 3.62 × 10−3 | |
| PEXP_095510 | - | −2.89 | 7.63 × 10−8 | |
| PEXP_096630 | Pigment | −12.2 | 4.72 × 10−20 | |
| PEXP_097790 | - | 1.52 | 4.63 × 10−3 | |
| PEXP_099180 | - | −1.81 | 4.23 × 10−10 | |
| PEXP_102410 | - | −1.92 | 3.65 × 10−10 | |
| NRPS (Non ribosomal peptide synthetase) | PEXP_012360 | - | −1.35 | 7.72 × 10−8 |
| PEXP_015170 | Fungisporins | 1.71 | 1.34 × 10−7 | |
| PEXP_018960 | - | −8.55 | 2.90 × 10−11 | |
| PEXP_029660 | - | −2.97 | 7.36 × 10−10 | |
| PEXP_030090 | Roquefortine C | 0.99 | 2.57 × 10−3 | |
| PEXP_055140 | - | 1.55 | 1.17 × 10−8 | |
| PEXP_095540 | - | −3.46 | 7.65 × 10−9 | |
| PEXP_096300 | - | 2.95 | 1.96 × 10−11 | |
| PEXP_104890 | - | −1.1 | 2.04 × 10−4 | |
| Hybrid PKS/NRPS | PEXP_008740 | - | 1.02 | 1.41 × 10−5 |
| PEXP_074060 | Chaetoglobosins | 2.17 | 5.15 × 10−10 | |
| NRPS-like | PEXP_045260 | - | 3.89 | 8.31 × 10−7 |
| PEXP_050450 | - | 1.03 | 2.53 × 10−7 | |
| PEXP_060620 | - | 2.11 | 5.21 × 10−11 | |
| PEXP_072870 | - | −2.05 | 6.25 × 10−8 | |
| PEXP_080590 | - | −1.4 | 1.68 × 10−8 | |
| PEXP_082750 | - | 1.18 | 7.81 × 10−9 | |
| PEXP_095480 | - | 1.43 | 2.51 × 10−10 | |
| Terpene cyclase | PEXP_043150 | - | −2.68 | 1.19 × 10−4 |
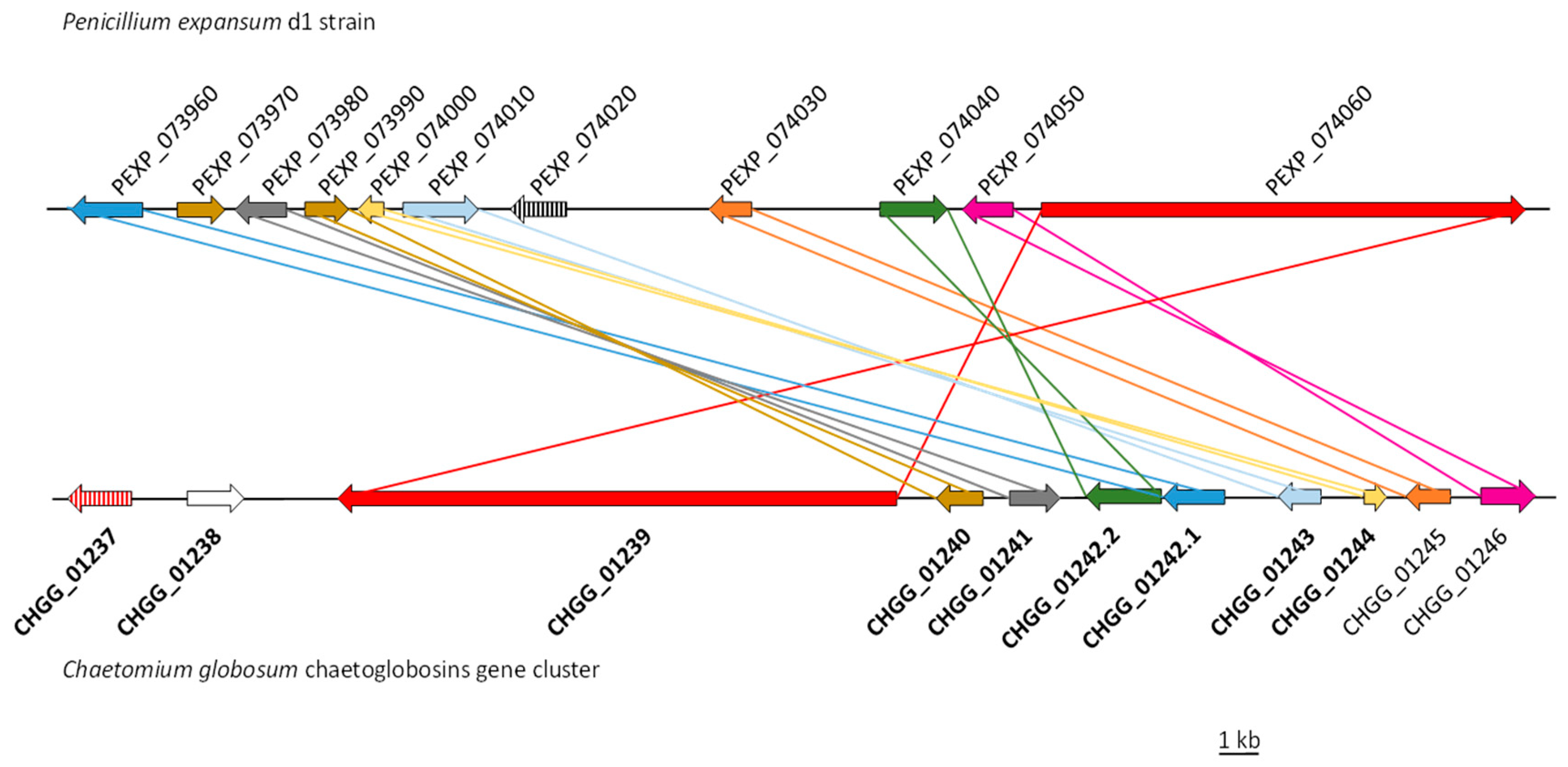
| Penicillium expansum Strain d1 | Chaetomium globosum Strain CBS 148.51 | % Identity/Similarity | Log2 Fold Change PeΔbrlA vs. WT | Putative Function |
|---|---|---|---|---|
| PEXP_073960 | CHGG_01242.1/CHGG_05285 | 48/64; 53/70 | 1.84 | CYP450 |
| PEXP_073970 | CHGG_01240/CHGG_05283 | 41/63; 64/79 | 2.07 | Enoyl reductase |
| PEXP_073980 | CHGG_01241/CHGG_05282 | 48/64; 65/80 | 1.68 | Hypothetical protein |
| PEXP_073990 | CHGG_01240/CHGG_05283 | 47/65; 39/54 | 1.89 | Enoyl reductase |
| PEXP_074000 | CHGG_01244 | 41/55; | 2.08 | Hypothetical protein |
| PEXP_074010 | CHGG_01243/CHGG_05281 | 47/66; 53/71 | 1.82 | CYP P450 |
| PEXP_074020 | CHGG_05287 | 31/49; | 1.82 | Transcription factor * |
| PEXP_074030 | CHGG_01245/CHGG_05284 | 47/62; 43/59 | 1.95 | Short-chain dehydrogenase |
| PEXP_074040 | CHGG_01242.2/CHGG_05280 | 38/53; 48/65 | 1.84 | FAD-dependent oxidoreductase |
| PEXP_074050 | CHGG_01246/CHGG_05287 | 54/70; 54/72 | 1.97 | Alpha/beta hydrolase |
| PEXP_074060 | CHGG_01239/CHGG_05286 | 43/61; 52/68 | 2.17 | PKS-NRPS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zetina-Serrano, C.; Rocher, O.; Naylies, C.; Lippi, Y.; Oswald, I.P.; Lorber, S.; Puel, O. The brlA Gene Deletion Reveals That Patulin Biosynthesis Is Not Related to Conidiation in Penicillium expansum. Int. J. Mol. Sci. 2020, 21, 6660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186660
Zetina-Serrano C, Rocher O, Naylies C, Lippi Y, Oswald IP, Lorber S, Puel O. The brlA Gene Deletion Reveals That Patulin Biosynthesis Is Not Related to Conidiation in Penicillium expansum. International Journal of Molecular Sciences. 2020; 21(18):6660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186660
Chicago/Turabian StyleZetina-Serrano, Chrystian, Ophélie Rocher, Claire Naylies, Yannick Lippi, Isabelle P. Oswald, Sophie Lorber, and Olivier Puel. 2020. "The brlA Gene Deletion Reveals That Patulin Biosynthesis Is Not Related to Conidiation in Penicillium expansum" International Journal of Molecular Sciences 21, no. 18: 6660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186660