Intravenous Glutamine Administration Improves Glucose Tolerance and Attenuates the Inflammatory Response in Diet-Induced Obese Mice after Sleeve Gastrectomy

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Measurements of Plasma Biochemical Markers and Adipokines
2.4. Inflammatory Cytokine Concentrations in PLF
2.5. Messenger (m)RNA Extraction and a Real-Time Reverse-Transcription (RT) Quantitative Polymerase Chain Reaction (qPCR) Analysis
2.6. Proteomics Sample Preparation
2.7. UPLC-Q-TOF MS Analysis
2.8. Differential Label-Free Quantitative and Qualitative Proteomics
2.9. Ingenuity Pathway Analysis (IPA) of the Proteomic Profiles in the Liver
2.10. Statistical Analyses
3. Results
3.1. BW and Epididymal Weight Changes after Feeding the High-Fat Diet for 10 Weeks
3.2. Plasma Adipokine, Glucose, and Insulin Levels after the High-Fat Diet Intervention
3.3. Effects of GLN on BW Change after a Gastrectomy
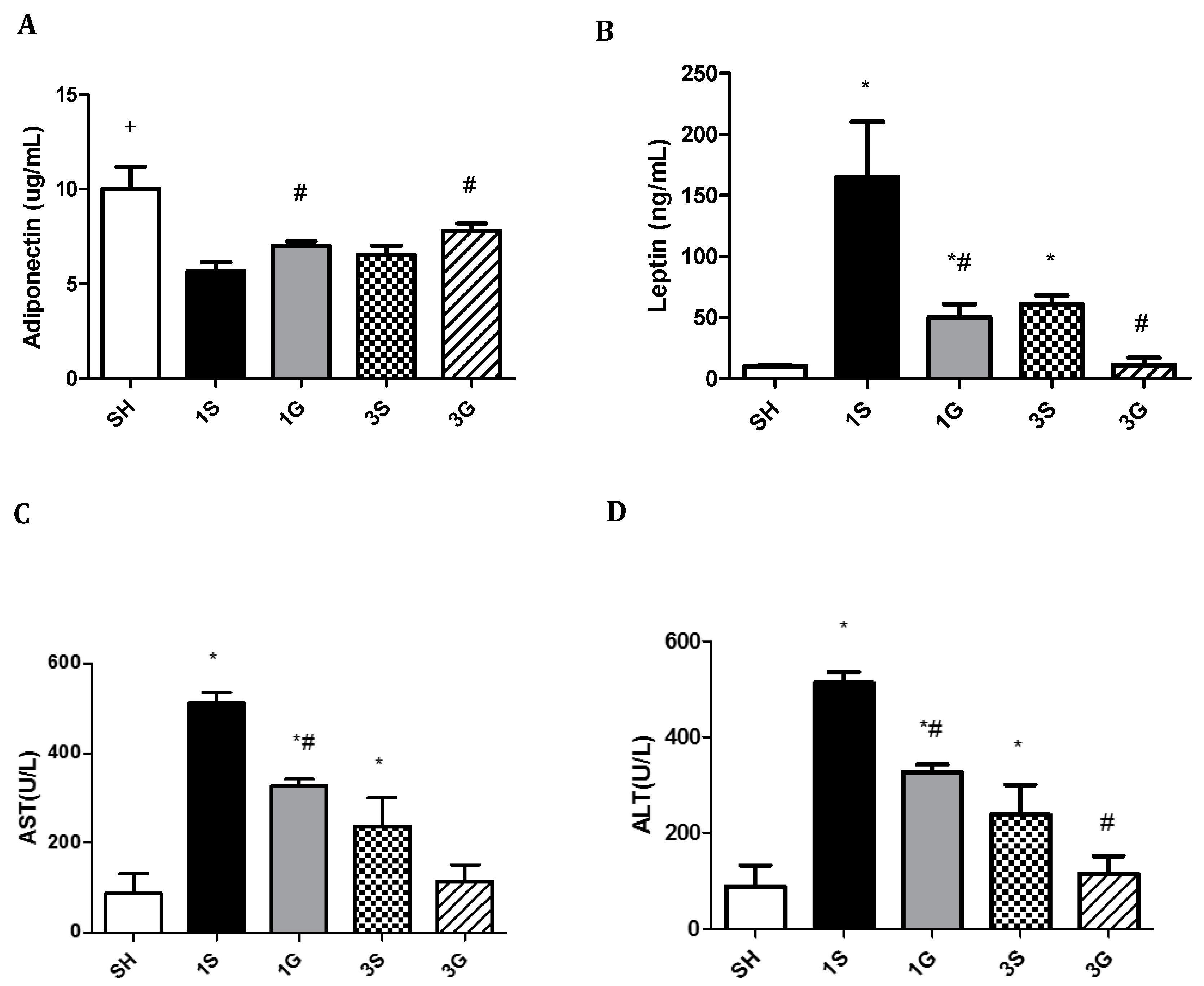
3.4. Effects of GLN on Plasma Biochemical Parameters in Obese Mice after a Gastrectomy
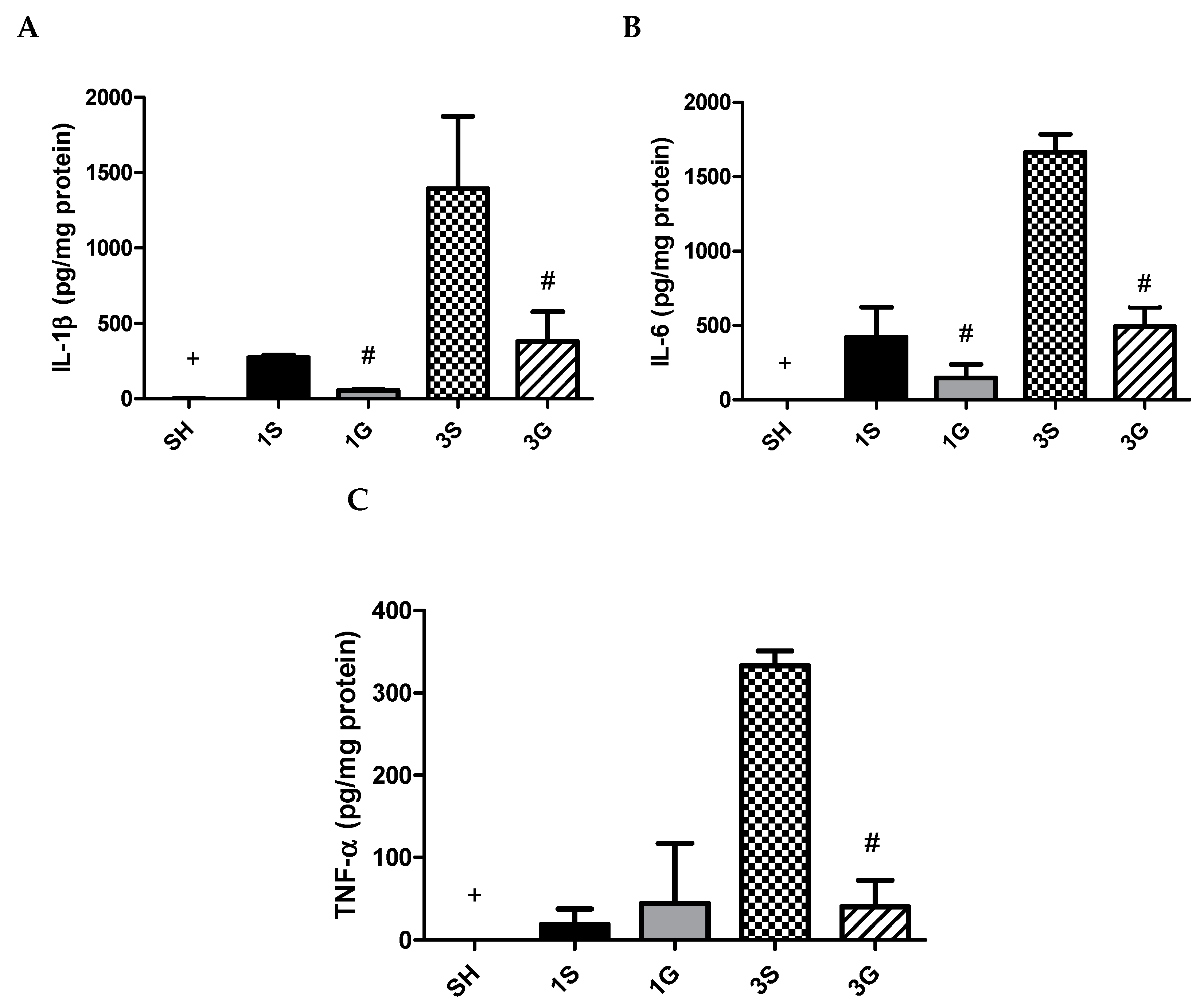
3.5. Effects of GLN on Cytokine Levels in PLF
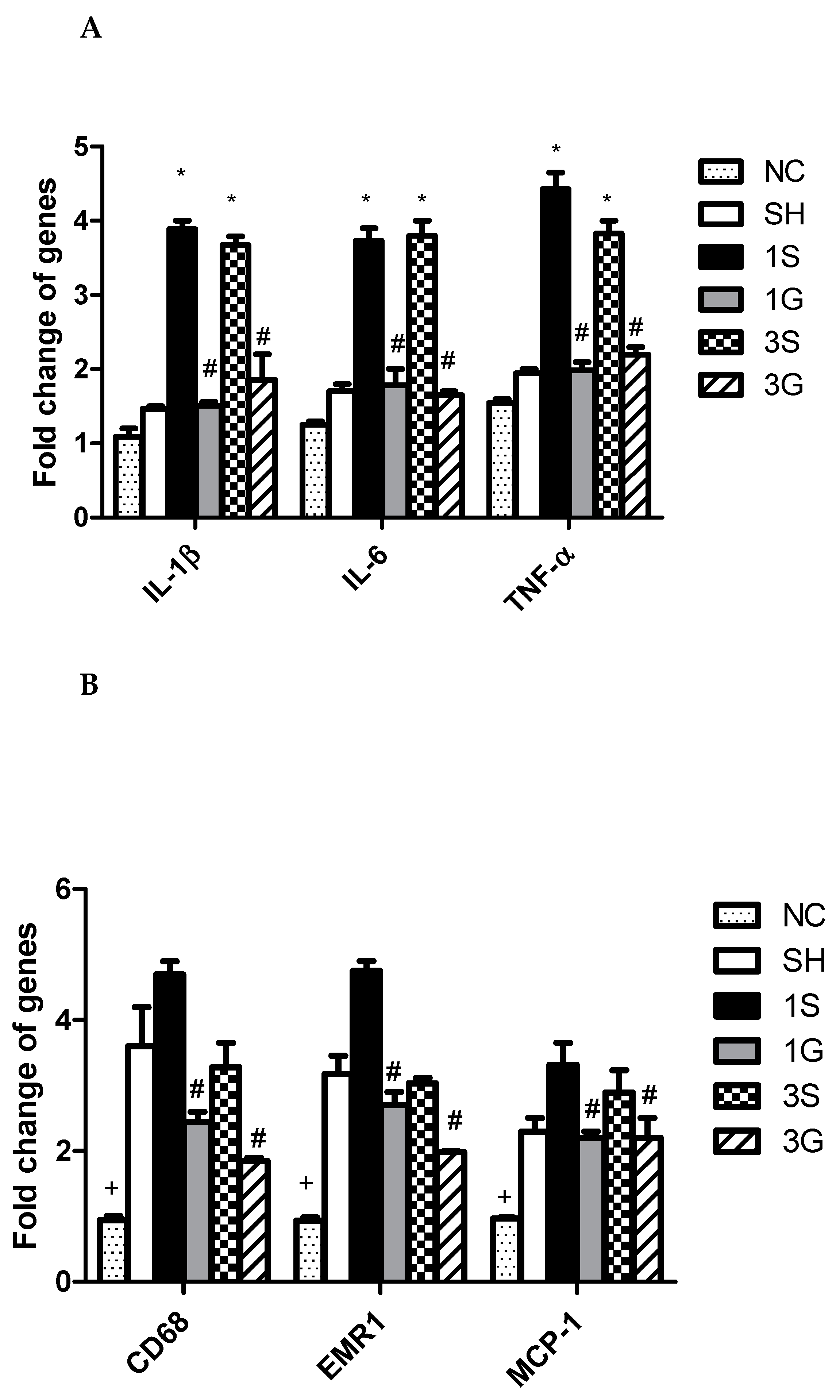
3.6. Expression of Macrophage Infiltration Markers and Inflammatory Mediator mRNAs in Epididymal Tissues after the Gastrectomy
3.7. Hepatic Proteomic Profiles Analyzed by the IPA
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chang, H.C.; Yang, H.C.; Chang, H.Y.; Yeh, C.J.; Chen, H.H.; Huang, K.C.; Pan, W.H. Morbid obesity in Taiwan: Prevalence, trends, associated social demographics, and lifestyle factors. PLoS ONE 2017, 12, e0169577. [Google Scholar] [CrossRef]
- Burhans, M.S.; Hagman, D.K.; Kuzma, J.N.; Schmidt, K.A.; Kratz, M. Contribution of adipose tissue inflammation to the development of type 2 diabetes mellitus. Compr. Physiol. 2018, 9, 1–58. [Google Scholar]
- Eckel, R.H.; York, D.A.; Rossner, S.; Hubbard, V.; Caterson, I.; St Jeor, S.T.; Laura, L.; Hayman, L.L.; Mullis, R.M.; Blair, S.N. Prevention Conference VII: Obesity, a worldwide epidemic related to heart disease and stroke: Executive summary. Circulation 2004, 110, 2968–2975. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.F.; Schatzkin, A.; Harris, T.B.; Kipnis, V.; Mouw, T.; Ballard-Barbash, R.; Hollenbeck, A.; Leitzmann, M.F. Overweight, obesity, and mortality in a large prospective cohort of persons 50 to 71 years old. N. Engl. J. Med. 2006, 355, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Daniel, S.; Soleymani, T.; Garvey, W.T. A complications-based clinical staging of obesity to guide treatment modality and intensity. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 377–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, A.G.; Wadden, T.A. The evolution of very-low-calorie diets: An update and meta-analysis. Obesity 2006, 14, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Hung, C.K.; Chang, Y.Y.; Tai, C.M.; Lin, J.T.; Wang, J.D. Health-related quality of life in adult patients with morbid obesity coming for bariatric surgery. Obes. Surg. 2010, 20, 1121–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenberg, D.; Bellatorre, A.; Bellatorre, N. Sleeve gastrectomy as a stand-alone bariatric operation for severe, morbid, and super obesity. JSLS 2013, 17, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, H.J.; Kim, G.; Chew, C.A.Z.; San, M.T.; So, J.B.; Shabbir, A. Is laparoscopic sleeve gastrectomy for Asian super obese a safe and effective procedure? Ann. Acad. Med. Singapore 2018, 47, 177–184. [Google Scholar]
- Tsai, P.H.; Liu, J.J.; Yeh, C.L.; Chiu, W.C.; Yeh, S.L. Effects of glutamine supplementation on oxidative stress-related gene expression and antioxidant properties in rats with streptozotocin-induced type 2 diabetes. Br. J. Nutr. 2012, 107, 1112–1118. [Google Scholar] [CrossRef] [Green Version]
- Wischmeyer, P.E.; Dhaliwal, R.; McCall, M.; Ziegler, T.R.; Heyland, D.K. Parenteral glutamine supplementation in critical illness: A systematic review. Crit. Care 2014, 18, R76. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.S.; Wu, J.M.; Lee, P.C.; Kuo, T.C.; Chen, P.D.; Hou, Y.C.; Yeh, S.L.; Lin, M.T. Antecedent administration of glutamine benefits the homeostasis of CD4(+) T cells and attenuates lung injury in mice with gut-derived polymicrobial sepsis. J. Parenter. Enteral Nutr. 2019, 43, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Klek, S.; Kulig, J.; Szczepanik, A.M.; Jedrys, J.; Kolodziejczyk, P. The clinical value of parenteral immunonutrition in surgical patients. Acta Chir. Belg. 2005, 105, 175–179. [Google Scholar] [PubMed]
- Wu, J.M.; Ho, T.W.; Lai, I.R.; Chen, C.N.; Lin, M.T. Parenteral glutamine supplementation improves serum albumin values 1 in surgical cancer patients. Clin. Nutr. 2020, 0261. [Google Scholar] [CrossRef] [PubMed]
- Abboud, K.Y.; Reis, S.K.; Martelli, M.E.; Zordão, O.P.; Tannihão, F.; de Souza, A.Z.Z.; Assalin, H.B.; Guadagnini, D.; Rocha, G.Z.; Saad, M.J.A.; et al. Oral glutamine supplementation reduces obesity, pro-inflammatory markers, and improves insulin sensitivity in DIO wistar rats and reduces waist circumference in overweight and obese humans. Nutrients 2019, 11, 536. [Google Scholar] [CrossRef] [Green Version]
- Yin, D.P.; Boyd, K.L.; Williams, P.E.; Abumrad, N.N.; Wasserman, D.H. Mouse models of bariatric surgery. Curr. Protoc. Mouse Biol. 2012, 2, 295–306. [Google Scholar]
- Hu, Y.M.; Hsiung, Y.C.; Pai, M.H.; Yeh, S.L. Glutamine administration in early or late septic phase downregulates lymphocyte PD1/PD-L1 expression and the inflammatory response in mice with polymicrobial sepsis. J. Parenter. Enter Nutr. 2018, 42, 538–549. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Arterburn, D.E.; Telem, D.A.; Kushner, R.F.; Courcoulas, A.P. Benefits and risks of bariatric surgery in adults. JAMA 2020, 324, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Zakkar, M.; Ascione, R.; James, A.F.; Angelini, G.D.; Suleiman, M.S. Inflammation, oxidative stress and postoperative atrial fibrillation in cardiac surgery. Pharmacol. Ther. 2015, 154, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Burkholder, T.; Foltz, C.; Karlsson, E.; Linton, C.G.; Smith, J.M. Health evaluation of experimental laboratory mice. Curr. Protoc. Mouse Biol. 2012, 2, 145–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneck, A.S.; Iannelli, A.; Patouraux, S.; Rousseau, D.; Bonnafous, S.; Bailly-Maitre, B.; Thuc, O.L.; Rovere, C.; Panaia-Ferrari, P.; Anty, R.; et al. Effects of sleeve gastrectomy in high fat diet-induced obese mice: Respective role of reduced caloric intake, white adipose tissue inflammation and changes in adipose tissue and ectopic fat depots. Surg. Endosc. 2014, 28, 592–602. [Google Scholar] [CrossRef]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [Green Version]
- La Cava, A. Leptin in inflammation and autoimmunity. Cytokine 2017, 98, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Killingsworth, M.C.; Myasoedova, V.A.; Orekhov, A.N.; Bobryshev, Y.V. CD68/macrosialin: Not just a histochemical marker. Lab. Investig. 2017, 97, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austyn, J.M.; Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur. J. Immunol. 1981, 11, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, A.; Hogaboam, C.M.; Lukacs, N.W.; Lincoln, P.M.; Strieter, R.M.; Kunkel, S.L. Endogenous monocyte chemoattractant protein-1 (MCP-1) protects mice in a model of acute septic peritonitis: Cross-talk between MCP-1 and leukotriene B4. J. Immunol. 1999, 163, 6148–6154. [Google Scholar] [PubMed]
- Nguyen, N.T.; Braley, S.; Fleming, N.W.; Lambourne, L.; Rivers, R.; Wolfe, B.M. Comparison of postoperative hepatic function after laparoscopic versus open gastric bypass. Am. J. Surg. 2003, 186, 40–44. [Google Scholar] [CrossRef]
- Murray, R.K.; Granner, D.K.; Mayes, P.A.; Rodwell, V.W. Harper’s Biochemistry, 25th ed.; Appleton & Lange: Stanford, CT, USA, 2000. [Google Scholar]
- Pitaloka, D.M.I.; Ko, C.H.; Lin, M.T.; Yeh, S.L.; Yeh, C.L. Glutamine administration promotes hepatic glucose homeostasis through regulating the PI3K/Akt pathway in high-fat diet-induced obese mice with limb ischemia. Nutr. Res. 2019, 68, 45–53. [Google Scholar] [CrossRef]
- Maeda, N.; Shimomura, I.; Kishida, K.; Nishizawa, H.; Matsuda, M.; Nagaretani, H.; Furuyama, N.; Kondo, H.; Takahashi, M.; Arita, Y.; et al. Diet-induced insulin resistance in mice lacking adiponectin/ACRP30. Nat. Med. 2002, 8, 731–737. [Google Scholar] [CrossRef]
- Yatagai, T.; Nagasaka, S.; Taniguchi, A.; Fukushima, M.; Nakamura, T.; Kuroe, A.; Nakai, Y.; Ishibashi, S. Hypoadiponectinemia is associated with visceral fat accumulation and insulin resistance in Japanese men with type 2 diabetes mellitus. Metabolism 2003, 52, 1274–1278. [Google Scholar] [CrossRef]
- Petrus, P.; Lecoutre, S.; Dollet, L.; Wiel, C.; Sulen, A.; Gao, H.; Tavira, B.; Laurencikiene, J.; Rooyackers, O.; Checa, A.; et al. Glutamine links obesity to inflammation in human white adipose tissue. Cell Metab. 2020, 31, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammatory pathways and insulin action. Int. J. Obes. Relat. Metab Disord. 2003, 27 (Suppl. 3), S53–S55. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | g/kg |
|---|---|
| Casein | 259.13 |
| L-Cysteine | 3.89 |
| Maltodextrin | 161.96 |
| Sucrose | 89.14 |
| Cellulose | 64.78 |
| Soybean oil | 32.39 |
| Lard | 317.44 |
| Mineral mix 1 | 12.96 |
| Dicalcium phosphate | 16.84 |
| Calcium carbonate, 1H2O | 7.13 |
| Potassium citrate | 21.38 |
| Vitamin mix 2 | 12.96 |
| Total | 1000 |
| Primer Sequences (5′→3′) | ||
|---|---|---|
| GAPDH | Forward | TGCACCACCAACTGCTTAG |
| Reverse | GGATGCAGGGATGATGTTC | |
| TNF-α | Forward | AAATGGGCTCCCTCTCATCAGTTC |
| Reverse | TCTGCTTGGTGGTTTGCTACGAC | |
| IL-1β | Forward | TGCCACCTTTTGACAGTGATG |
| Reverse | ATGTGCTGCTGCGAGATTT | |
| IL-6 | Forward | TCCTACCCCAACTTCCAATGCTC |
| Reverse | TTGGATGGTCTTGGTCCTTAGCC | |
| CD68 | Forward | TGTTCAGCTCCAAGCCCAAA |
| Reverse | ACTCGGGCTCTGATGTAGGT | |
| EMR-1 | Forward | ACCTTGTGGTCCTAACTCAGTC |
| Reverse | ACAAAGCCTGGTTGACAGGTA | |
| MCP-1 | Forward | GATTCACATTTGCGCTGCCT |
| Reverse | TGAGCCTGGGAGATCACCAT |
| Parameter | NC | HF |
|---|---|---|
| Leptin (ng/mL) | 4.64 ± 0.85 | 11.01 ± 1.40 * |
| Adiponectin (µg/mL) | 5.69 ± 0.56 | 9.98 ± 0.80 * |
| Glucose (mg/dL) | 100.32 ± 20.38 | 221.47 ± 21.36 * |
| Insulin (µIU/mL) | 18.65 ± 0.12 | 55.23 ± 1.12 * |
| Group | Glucose (mg/dL) | Insulin (μIU/mL) |
|---|---|---|
| SH | 213.9 ± 10.6 | 96.2 ± 27.2 |
| 1S | 225.5 ± 10.3 | 88.9 ± 12.7 |
| 1G | 122.4 ± 12.2 *,# | 40.6 ± 6.1 *,# |
| 3S | 189.6 ± 10.1 | 80.2 ± 8.8 |
| 3G | 108.2 ± 6.2 *,# | 42.8 ± 6.6 *,# |
| Identifier | Enzymes | Description | Log2 (3S/3G) | p Value |
|---|---|---|---|---|
| Q91Y97 | ALDOB | Aldolase, fructose-bisphosphate B | 0.76 | 0.001 |
| P17182 | ENO1 | Enolase 1 | 0.68 | 0.006 |
| Q9QXD6 | FBP1 | Fructose-bisphosphatase 1 | 0.69 | 0.03 |
| P14152 | MDH1 | Malate dehydrogenase 1 | 0.41 | 0.04 |
| Q05920 | PC | Pyruvate carboxylase | 1.18 | 0.03 |
| P16858 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | 1.6 | 0.03 |
| P17751 | TPI1 | Triosephosphate isomerase | 1.59 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, C.-L.; Yang, P.-J.; Lee, P.-C.; Wu, J.-M.; Chen, P.-D.; Huang, C.-C.; Yeh, S.-L.; Lin, M.-T. Intravenous Glutamine Administration Improves Glucose Tolerance and Attenuates the Inflammatory Response in Diet-Induced Obese Mice after Sleeve Gastrectomy. Nutrients 2020, 12, 3192. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103192
Yeh C-L, Yang P-J, Lee P-C, Wu J-M, Chen P-D, Huang C-C, Yeh S-L, Lin M-T. Intravenous Glutamine Administration Improves Glucose Tolerance and Attenuates the Inflammatory Response in Diet-Induced Obese Mice after Sleeve Gastrectomy. Nutrients. 2020; 12(10):3192. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103192
Chicago/Turabian StyleYeh, Chiu-Li, Po-Jen Yang, Po-Chu Lee, Jin-Ming Wu, Po-Da Chen, Chun-Chieh Huang, Sung-Ling Yeh, and Ming-Tsan Lin. 2020. "Intravenous Glutamine Administration Improves Glucose Tolerance and Attenuates the Inflammatory Response in Diet-Induced Obese Mice after Sleeve Gastrectomy" Nutrients 12, no. 10: 3192. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103192